病毒学论文

摘要:猪细小病毒(Porcine parvovirus, PPV)是猪繁殖障碍最常见和最重要的病原之一。一直以来,人们认为PPV的遗传变异性很低,通过接种疫苗可以很好地控制该病毒的传播。然而,该病毒的多样性比预期的要大得多,并且一些新出现的强毒株不能被经典PPV疫苗株的抗血清有效中和,导致免疫失败。通过总结PPV研究的最新成果,全面更新对PPV生物学的理解,揭示在选择压力下,PPV新毒株出现的潜在风险及其对养猪业的危害,旨在为制定PPV的有效防控策略提供参考。
关键词:猪细小病毒; 生物学特性; 发病机制; 遗传变异;
作者简介: 陈玉梅( 1982-) ,女,河南南阳人,讲师,博士,主要从事动物疫病免疫机制与疫苗研究。E-mail: yumeichen2012@163.com; 周景明( 1972-) ,男,河南新野人,教授,博士,主要从事畜禽疫病快速诊断技术、动物疫病免疫机制与疫苗研究。E- mail: zhjingming@126.com;
The Novel Progress of the Biology of Porcine Parvovirus
Abstract:Porcine parvovirus (PPV) is the most common and important infectious agents causing porcine reproductive failure. All the time, it was thought that PPV had low genetic variance, and its harmful effect on pig industry could be well-controlled by vaccination. However, subsequent studies found that the persity of PPV was much greater than previously anticipated.Some of the new highly virulent, which cannot be neutralized effectively by antisera raised agains told classical PPV vaccine strains, were isolated. The purpose of this review is to summarize the recent results of PPV research, update the present understanding of PPV biology,and reveal the potential risks of new PPV strains and their harm to pig industry under the pressure of selection, which will provide reference for formulating effective prevention and control strategies of PPV.
Keyword:Porcine parvovirus; Biological characteristics; Pathogenesis; Genetic variation;
猪细小病毒(Porcine parvovirus, PPV)属细小病毒科(Parvoviridae)、原细小病毒属(Protoparvovirus)中的有蹄类原细小病毒1(Ungulate parvovirus 1,UPV1)[1].PPV最早于20世纪60年代末被确认为细小病毒科成员,是SMEDI综合征(Stillbirths, mummification, embryonic death, and infertility,SMEDI)即猪死胎、木乃伊化、胚胎死亡和不孕的病原体[2],对养猪业危害极大。一直以来,人们认为PPV的遗传变异性很低,通过接种疫苗可以很好地控制该病毒的传播。然而,随着不能被经典PPV疫苗的抗血清有效中和的强毒株的出现,人们逐渐意识到该病毒的多样性比先前预期的要大得多。因此,追踪PPV研究的最新成果,全面更新对PPV生物学的理解,对于制定PPV的有效防控策略具有重要参考意义。
1、PPV的分子生物学特性
PPV是一种小的、无包膜的单链DNA病毒,基因组长约4~6.3 kb[3].病毒的2个末端序列形成大约120~200个碱基的复杂回文发夹结构(呈类似的“Y”或“T”形)[4].PPV的紧凑型基因组只包含2个启动子,共编码7种蛋白质,并利用选择性剪接技术来扩展基因组的编码能力。2种非结构蛋白NS1和NS2是从p4启动子转录而来,这些蛋白质对病毒复制特别是DNA复制很重要[4];另外,还可能含有一种假定的非结构蛋白NS3.PPV的2种结构蛋白(VP1和VP2)从p40启动子转录。二者是从一组嵌套的编码序列中翻译出来的,它们的氨基末端不同,较小的VP2是从与较大的VP1相同的RNA模板中剪接产生的。VP1有729个氨基酸残基(aa),其中氨基末端前120个氨基酸是VP1特有的(VP1 unique region,VP1u)。而PPV的第三种结构蛋白VP3是VP2的翻译后修饰产物。此外,晚期非结构蛋白(SAT)是由与VP2相同的mRNA启动VP2起始密码子下游的7个核苷酸表达而来的。PPV的外壳是由60个VP1或VP2结构蛋白分子组成的球形结构,呈二十面体对称排列。每个衣壳亚单位由8条反平行β链及1个α螺旋和4个环(Loop)组成(图1) [4,5].VP1蛋白大约占病毒粒子的10%,主要在病毒复制和感染细胞时发挥作用,VP2蛋白是PPV的主要结构蛋白,占病毒粒子的60%以上,也是病毒的主要免疫原性蛋白。VP1/VP2基因对于PPV的系统发育分析至关重要[6].
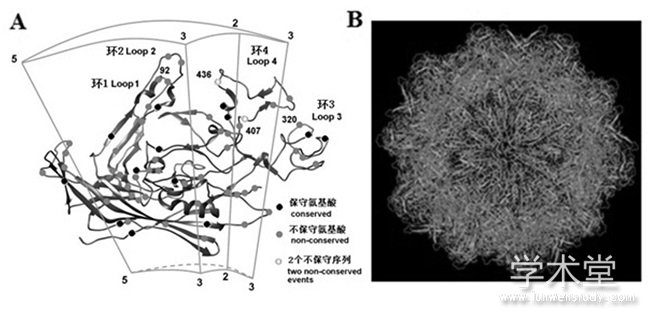
图1 PPV VP2蛋白3D模型(A)及病毒衣壳结构(B)
2、PPV发病机制
PPV毒株毒力是由其可能引起的繁殖失败的严重程度来定义的[7].大多数情况下,未怀孕成年猪或仔猪单纯感染PPV不会出现临床症状,而胎儿感染的结果随母猪妊娠的进展而变化,流行病学研究表明,母猪妊娠前半期PPV感染可导致生殖衰竭,PPV经口服途径初次感染母猪后23~32 d穿过胎盘,经肌肉注射途径感染时间稍短,约15 d [8].研究发现,仅在胎儿产生抗体前发生的感染才会导致死亡和木乃伊化,妊娠70 d后,胎儿产生抗体反应,通常在这一时期以后的感染胎儿可以存活(图2)[2].除感染时间外,病毒的遗传组成对胎儿感染的结果有决定性影响,低致病性毒株和疫苗株(如NADL-2和MSV)不能像高致病性毒株(如Kresse毒株和27A毒株)一样有效地穿过胎盘屏障[7],因此,它们对妊娠的影响不易检测到。然而,直接向羊水中注射NADL-2毒株可导致胎儿死亡[9].而关于PPV如何通过胎盘屏障诱导生殖失败还不清楚,没有证据表明PPV可在子宫上皮或原肌中复制,因此,通过屏障复制理论不成立[7].单核细胞和腹腔巨噬细胞可吞噬NADL-2,据此有研究者猜测PPV可通过母体巨噬细胞侵入胎儿[10].事实上,目前并没有任何直接证据表明巨噬细胞可以将PPV从母亲直接传递到胎儿。
用不同毒株PPV感染妊娠母猪的试验表明,NADL-8毒株要达到可经胎盘妊娠感染所需的毒量是NADL-2毒株的10 000倍以上[11].而PPV不同毒株感染的胚胎中病毒DNA的组织分布和数量存在显着差异,在肝脏中通过原位杂交试验检测到Kresse毒株和NADL-8毒株,而在大脑和脾脏仅检测到Kresse毒株[12].实时荧光定量PCR检测结果显示,PPV 27a分离株在包括结肠、十二指肠、空肠、心脏、肝脏、肺脏、肾脏、脾脏、胸腺、性腺等10个不同胎儿器官中的病毒滴度在1011~1015拷贝/106个细胞,而毒性较小的PPV-143a毒株和疫苗株(NADL-2毒株和MSV毒株)主要在肾脏中检出,且病毒滴度较低(<103拷贝/106个细胞)[9].相关研究结果表明,PPV毒株的组织特异性及其在胚胎中感染起始能力的差异可能在胎儿感染及致病性方面发挥重要作用[9,10,11,12].
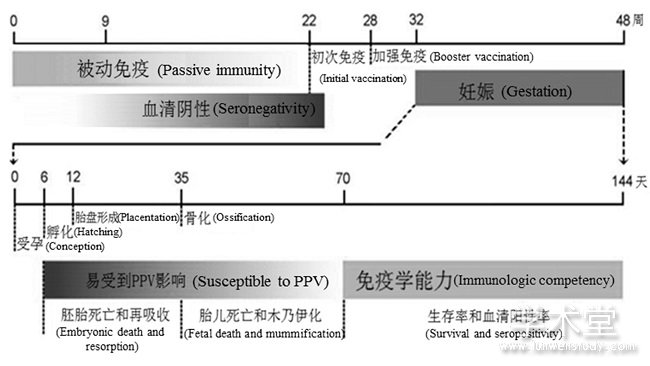
图2 PPV感染和免疫应答的主要时间点
3、PPV与细胞的相互作用
在乳猪和老年猪体内靶细胞中检测PPV比较困难。研究显示,PPV可以在活化的淋巴细胞中复制,但不能在血单核细胞中复制,而在巨噬细胞中的复制能力还存在争议[2].PCR检测发现,PPV可在心脏、肺脏、肾脏、脾脏、子宫内膜和小肠的细胞中繁殖,然而,PCR检测却不能区分是器官自身细胞中产生的病毒还是通过血液循环系统运输的病毒[9,13].PPV进入细胞的第一步是病毒离子与细胞表面糖蛋白的末端唾液酸位点结合,通过胞吞作用抑制剂阻断试验结果表明,除了网格蛋白介导的内吞作用和大胞饮作用外,还有第3种未知的进入机制可能参与PPV入胞,而小窝蛋白(Caveolae)介导的内吞途径不起任何作用,单个PPV颗粒倾向于通过网格蛋白介导的内吞作用进入胞内,而PPV聚集物倾向于大胞饮作用[14].核内体易位到晚期核内体或溶酶体以及感染后2~10 h的酸化作用对PPV的有效感染至关重要[14].研究表明,泛素化以及细胞质中蛋白酶体的相互作用是PPV有效感染必不可少的步骤,病毒颗粒向细胞核运动时涉及微管和肌动蛋白网络。微管在感染前8~10 h至关重要,这表明微管在PPV向核周区的内质转运中起重要作用。在感染后期(12~16 h)肌动蛋白活性对增殖性感染是必需的,它可能对传入病毒的转运以及对新合成蛋白质的核转运都是必需的[14].
核定位序列(Nuclear localization signals, NLS)对蛋白质能被顺利转运进细胞核至关重要。腺相关病毒的衣壳蛋白具有4个潜在的NLS,分别为以下4个基本区域(Basic region, BR):BR1、BR2、BR3和BR4.类似于小鼠微小病毒(Minute virus of mice,MVM),PPV的VP1u含有5个潜在的NLS,即BR1 (3PPAKRAR9)、BR2 (86RAKR89)、BR3 (110RRSPRK115)、BR4 (124KR125)和BR5 (133KKKAK137) [15].其中BR1是一个经典的含有7个氨基酸的NLS,而BR4和BR5是典型的二元核定位信号(Bipartite NLSs, B-NLSs),这2种NLSs都是病毒复制的必要条件[16],它们的突变不影响病毒的装配,但可影响病毒增殖,表明它们在感染早期负责病毒的核转运。另外,在PPV衣壳中还可能存在3个非线性核定位基序(Nuclear localization motif, NLM),包装好的PPV衣壳表面的外部区域(包含R374、R393和R565)、PPV VP2三聚体内部区域的中心区域(包含K475、R477)及病毒衣壳内表面的内部区域(包含K272、K275、K487、R533、K535、R576) [15,16].在所有原细小病毒中,VP1 NLS和VP2 NLM在病毒装配过程中被内化到细胞核中,且不可接近转运蛋白。研究证明,MVM是主动从核中运输出来的[17].据推测在原细小病毒粒子上,NLM或NLS的存在会干扰这种传输。然而,对PPV而言,目前并没有任何证据表明组装好的PPV病毒离子可以向细胞膜的任何囊泡转运[2].
目前,已知的大多数影响PPV生物学特性的突变都是在衣壳蛋白上发现的,唯一例外的是NADL-2毒株NS1蛋白的I481L突变,该突变位点与VP2的N348H突变一起作用有助于NADL-2毒株在犬细胞系A72中产生细胞病变效应并以高滴度复制。NADL-2毒株和Kresse毒株的基因组在右端尾部发夹结构附近有13个核苷酸(Nucleotides, nt)和127 nt的重复序列。这2种病毒的结构蛋白之间只有6个氨基酸差异,其中5个氨基酸差异(I215T、D378G、H383Q、S436P和R565K)也存在于其他毒力毒株中。定位于衣壳表面的3个氨基酸差异(D378G、H383Q和S436P)使NADL-2毒株不能在原代牛睾丸细胞(TV)中的复制,并将滴度和细胞病变效应降低到与猪源细胞系(PT和PFT)中Kresse毒株相当的水平[18],后来发现,S436P在体外不参与组织向性[3].由于在毒力毒株(27a毒株)和无毒力毒株(143a毒株)的436位氨基酸均为苏氨酸(Thr),进一步从侧面证实VP2的D378G和H383Q的氨基酸突变可能改变PPV的致病性[7].
PPV感染后3 h可促进PK-15细胞中p53的积累,p53通过线粒体介导的凋亡途径激活Caspase-9和Caspase-3,从而使线粒体释放细胞色素C[19].在PPV YL株感染60 h后的猪睾丸细胞(ST)和PK-15细胞中,凋亡细胞的比例可达50%,而在PPV Kresse毒株感染的PT细胞中,凋亡细胞的数量仍低于14%[2,20].Kresse毒株感染期间PT细胞死亡的主要形式包括感染核肿胀、早期细胞膜衰竭(如碘化丙啶摄取)、乳酸脱氢酶快速释放和病毒DNA自由释放,这些形式最终均指向坏死[20].即使PPV衣壳的微小突变也能改变其与宿主间的相互作用,并改变病毒的细胞病变效应[2,21].PPV感染所引发的病变效应在很大程度上取决于毒株类型和细胞类型。然而,细胞早期解体加速了PPV的体外释放和扩散,这些过程对体内病毒的生命周期有非常显着的影响。在PPV中已经进化出一种可以促进细胞快速溶解和病毒释放的小的选择性翻译蛋白(Small alternatively translated protein, SATp),该蛋白与VP2共用1套mRNA,其起始密码子是VP2起始密码子下游的7个核苷酸。SATp是一种内质网(ER)驻留的短膜蛋白(68AA),包含1个跨膜螺旋,可加速细胞死亡和病毒传播[2].无论是否存在SATp,在受感染的PT细胞中,PPV感染都会诱导未折叠蛋白反应(unfolded protein response, UPR)。在感染后12~14 h,它还导致抗凋亡的内质网应激蛋白X盒结合蛋白1(X-box binding protein 1, XBP1)可逆地激活。在感染后期,SATp的存在通过使ER应激不可逆加速细胞凋亡。
4、PPV的遗传变异与进化
一直以来,研究者认为PPV的基因组比其他细小病毒更保守[5].然而,最近研究者发现PPV的基因组并不是原以为的那么保守,通过系统研究野外分离株中VP蛋白的遗传多样性,PPV至少分为7个簇(Cluster),其中C和D簇中欧洲毒株占优势,而F簇中中国毒株占优势,未观察到野猪分离株的分簇与地理分布的相似关系[4].在同一地区,野猪种群中检测到的PPV比家猪中的PPV的具有更多的多样性。相关研究表明,传统的PPV疫苗株的抗血清不能有效中和D簇中一些新的强毒株[22].据推测,这些突变体可能是逃避疫苗接种所带来的免疫压力而产生的。然而,根据现有的序列分析发现,在组织培养或接种牛群中存在抗体时,观察到的PPV遗传多样性降低,而在疫苗株衣壳上发现的一些突变位点(NADL-2毒株 I320S,H383Q;Str.Challenge毒株S45T,P436S)也没有出现在新的PPV突变体上[23].因此,可以认为疫苗接种失败和未接种动物(如野猪)种群对新出现的表型产生的影响可能比接种种群更重要。
PPV分离株的系统发育与毒力之间没有显着的相关性。病毒毒力强弱可能直接或间接影响如组织特异性和长期/高滴度脱落等生物学特征,这也决定了PPV毒株在高度可变的田间条件下的适宜性。对田间毒株的NS和VP基因突变模式的研究表明,这2个编码区的进化有明显差异,VP基因的突变率是其中1个NS基因(10-5)的30~50倍(约3~5×10-4突变/核苷酸·年)[2].这与在NS区域发现的非同义和同义取代率之间的负差异一起表明,允许NS蛋白质维持功能的核苷酸变化数量是有限的,同时纯化选择(Purifying selection)即负选择(Negative selection)决定了该区域的进化。相反,早期的研究表明,VP1/VP2基因是在近中性模式(漂移)下进化的[2],但在决定细胞相互作用和免疫原性的衣壳外环上的几个位置被正选择(Positive selection)[24].此外,大多数突变发生在表面环,其中215、228、383、414、419、436位氨基酸似乎是衣壳中的主要可变位点[2,24].普遍认为,小DNA病毒中CpG缺失的主要原因是来自复制优势和/或免疫逃逸的自然选择。PPV基因组是CpG缺失的,CpG位点比PPV基因组中的GC或C和G位点更容易发生突变。因此,突变压力而不是选择力,是导致PPV基因组中CpG表达不足的原因[2].
5、PPV的检测与预防
PPV常用的检测方法有分子生物学检测方法、血清学检测方法等,其中分子生物学检测方法主要包括常规PCR、多重PCR、实时荧光定量PCR[25]、环介导等温扩增技术(LAMP)、聚合酶重组酶扩增技术(RPA)[26]以及基因芯片检测技术等[2].由于PPV能凝集鸡、豚鼠、小鼠、人、猴、鼠和猫的红细胞,因此,血凝(Hemagglutinationassays,HA)和血凝抑制试验(Hemagglutination inhibition assays,HAI)仍然是一种有效的PPV血清学检测方法,在研究和实践中广泛使用。而酶联免疫吸附试验(ELISA)是最常用的检测PPV特异性抗体的方法;血清学方法还包括病毒中和试验、改良的直接补体结合(Modified direct complement-fixation,MCDF)试验等。另外,还有基于NS1蛋白的区分感染动物和接种动物的重组pA(Differentiating infected from vaccinated animals)试验等[2].PPV病毒的感染一旦开始则很难清除,所以控制PPV流行的主要措施是接种疫苗。目前,使用较广泛的疫苗是PPV灭活苗和弱毒苗,另外PPV病毒样颗粒(VLPs)疫苗的研究也很多,PPV疫苗的应用在很大程度上控制了PPV的流行和传播[27].但由于PPV新的强毒株的出现及经典疫苗防治失败的案例的出现对PPV疫苗提出来新的挑战。
6、小结
随着生物学技术的飞速发展,PPV的分子生物学相关研究已取得了很大进展,在PPV的检测与预防方面也取得了进展,PPV疫苗的使用在很大程度上控制了PPV的流行和传播。但是,依然会有因免疫失败引发的PPV疫情报道。一直以来,人们认为该病毒的遗传变异性很低,然而,随后的研究揭示了该病毒的多样性比先前预期的要大得多,PPV的核苷酸替换率大约为每年每个位点对NS1基因进行10-5次替换,每年每个位点对VP1和VP2基因进行10-4次替换,这些比率与RNA病毒中的核苷酸替代率相似[2,3,28].因此,随着时间的推移,常有新的PPV毒株出现,对养猪业造成潜在的威胁。在过去的几年里,也有许多新的PPV毒株被报道,但是关于这些病毒在发生和流行的信息及其更深入的研究却很缺乏。适时追踪PPV研究前沿,密切监测新毒株的出现,并对这些新毒株与经典流行毒株的致病性等特征进行深入研究,对于新型广谱疫苗的研制及有效防控PPV新毒株的感染流行具有重要意义。
参考文献

人受凉后出现的喷嚏、咽喉疼、流涕、鼻塞、咳嗽、头痛、发烧等感染性症状称为感冒[1].现代医学将感冒归结为病毒感染引起,是人体抵抗力下降,病原微生物趁机侵入、大量繁殖所致[2].针对普通感冒的药物不计其数,多以减轻充血、抗过敏、止咳化痰及解热镇痛为...