动物学论文

摘 要: 触形大疣蛛是中国中东部唯数不多的广布性的地穴型原蛛。对触形大疣蛛地理区系演化进行研究, 旨在探明其时空演化规律, 核实或细化学界的某些观点。作者在前期研究基础上, 通过采用形态特征比值的定量聚类分析方法获得了该蛛26个样点间演化关系的支序图, 分析表明:触形大疣蛛种系源自于泥盆纪前东南盆地西北侧西南至东北向短暂相融形成的弧形海湾的多个侵入点, 同时, 伴随地质构造运动出现的种间分化是发生于这段时期的主要事件;结合近期在缅甸琥珀中发现1亿年前的有尾蜘蛛, 推测该蛛来源于西南方向的可能性更大些;随着浙闽地区的抬升使保留于“湘赣岛海”的种系逐渐由西向东渗入, 因第三纪地质气候变迁归于平缓, 便出现了江西境内的适应性辐射演化;第四纪冰期后, 泛江西区域的触形大疣蛛种系进入到深度分化和种群细分阶段。由于线粒体标记被广泛用于种内遗传结构和种群历史等各类研宄中。为进一步核实和细化该地理演化路径或明确其与地质地层发育的关系, 作者拟采用线粒体标记等对更多样点的触形大疣蛛样本的遗传分化状态进行综合分析, 期待更真实的种群细分, 以破解其演化历程。在综述其相关的研究背景后, 作者认为该蛛作为该区域地球地质等中长期环境变化的指示生物或许是最佳选择。
关键词: 地穴型原蛛; 触形大疣蛛; 区域性演化; 生物地理;
Abstract: Macrothele palpator is a crypt original spider of the only widely distributed in the central and eastern of China. The geographical evolution of M. palpator is studied to ascertain a law of this spider speciation within temporal and spatial evolution of the region, to verify or to refine a view put forward by some scholars. On the basis of previous studies, the authors obtain the cladistic chart of 26 points between these taxa by using the quantitative cluster analysis with the ratio of morphological characteristic value of M. palpator, and the results show that the species of M. palpator originated from several intrusive points of the arc-shaped bay formed from the southwest to the northeast of the south-west side of the pre-Devonian basin. And at the same time, the interspecific differentiation accompanied by geological tectonic movement is the main event in this period. Combined with the recent discovery of a 100 million years ago tail spider in Myanmar Amber, it is more likely that the spider originated in the southwest. With the uplift of Zhejiang and Fujian regions, the species retained in the "Xianggan Island Sea" gradually infiltrated from west to east, and the adaptive radiation evolution occurred in Jiangxi because the Tertiary geological and climatic changes were gentle. After the Quaternary glacial period, the species of M. palpator in Pan-Jiangxi region entered the stage of deep differentiation and population subdivision. The mitochondrial marker is widely used in the research of intraspecific genetic structure and population history and so on. So, to further verify and refine the geographic evolution way or clarify its relationship with the geological formation and development of its distribution areas, the applicant intends to analyze the state of genetic differentiation of spider samples from more diverse points in molecular level by using the mitochondrial marker and so on, looking for a more reliable population subdivision to unveil its evolution. In a word, it is concluded that M. palpator as an organism indicator of medium and long-term environmental changes in the earth's geology, may be the best choice after the review of its related research background.
Keyword: crypt original spider; Macrothele palpator; regional evolution; biogeography;
从物种多样性、生态功能和进化史来讲, 蜘蛛是陆地生态系统的重要组成部分, 更是农业生态系统中防治害虫的基础, 蛛丝和蛛毒也是新材料合成的重要资源[1,2]。蜘蛛是一个生物多样性丰富的类群, 也是陆地最古老谱系的代表之一, 其起源可以追溯到泥盆纪[3]。其实, 作为人类活动的结果, 有相当多的蜘蛛已濒临灭绝, 更具体地说, 它们的自然栖息地受到破坏[4]。
1、 蜘蛛地理分布格局与特点
蜘蛛是一类种类繁多、数量巨大、分布广泛的捕食性生物。至2019 年, 全世界蜘蛛目共有47 963个种, 隶属于117 科 4 118 属[2]。其科、属、种 3 个分类阶元的分布域非常悬殊, 90%的种分布在一个区界内, 90%的科是跨区界分布或全球分布, 属介于之间。按动物地理区域, 古北界和新热带界较多, 新北界较少。根据地理条件、生态条件和蜘蛛的分布状况, 申效诚等用新近提出的相似性通用公式和据此创立的多元相似性聚类分析方法, 分别对蜘蛛目的属、种两级的分布进行分析, 将全球陆地分为 53 个基础地理单元。和华莱士主要以哺乳动物建立的世界动物地理区划相比, 主要差异是:1) 古北界东、西两部分差异显着, 可分设两界;2) 新西兰和澳洲大陆相似性较低, 可单独设界;3) 新几内亚和太平洋岛屿与澳洲大陆的关系远于和东洋界的关系, 华莱士线在两界间的作用似乎不存在;4) 新热带界的中美地区似乎属于新北界更为合适, 并由此产生了南北美洲间的紧密联系;5) 新北界与古北界的相似性关系弱于与新热带界的关系, 全北界的概念几近消失[5]。这是生物进化与地球板块结构共同作用的结果, 也是低等动物与高等动物分布格局不同的具体体现。

蜘蛛是节肢动物门的一个类群, 进化历史已有3亿多年, 比只有6 500万年的哺乳动物经历过多得多的历史事件;蜘蛛是变温动物, 受环境影响比哺乳动物大得多;蜘蛛多以昆虫为食, 昆虫又大多以植物为食, 食物链的关系决定蜘蛛在哺乳动物、植物和昆虫之间, 会与昆虫最密切, 再与植物为接近, 和哺乳动物最远。况且, 目前仍然沿用的19世纪对哺乳动物和有花植物的阐释[6]都是定性研究的结果, 各大地理区域间的关系还必须接受定量分析的检验[5]。
2、 蜘蛛类群 (蜘蛛目:原蛛下目) 的生物地理学研究
生物地理学研究包括历史生物地理学、保护生物地理学和系统发生生物地理学等方面的研究。对于蜘蛛目、原蛛下目的蜘蛛类群来说, 近来有关其研究的报道也涉及到上述诸多方面。
在历史生物地理学方面, Paul 等人做了很多富有意义的开拓性工作, 他系统整理和研究了现有的化石蜘蛛[3]。化石蜘蛛可追溯到古生代, 它可能是中-古生代首波登陆动物的一部分[7]。化石蜘蛛记录已经显示出, 它们的主要捕食猎物即昆虫可能存在历经地质学年代的捕食者-猎物间协同辐射演化的历程。现存蜘蛛的某些种类、科、或群落 (集群) 可以被视为其它物种存在、气候条件、特定栖息地类型等的指标[3]。化石掺入生物地理学的研究可以对某个结果导致形成的结论产生深远的影响, 特别是当化石与现存范围不重叠时。为了更好地了解古蜘蛛和它们的谱系亲戚的生物地理格局, Hannah 等人结合分子和形态学数据采用宽松时钟来估计二纺蛛的系统发育。分析表明, 北半球和南半球二纺蛛的古老谱系与泛大陆解体成劳亚大陆和冈瓦纳大陆重合。该实例也证实了一个基于古大陆板块分裂而导致的地理分隔的生物地理假说[8]。
在保护生物地理学方面, Nelson 等人采用洞悉原蛛的定量生物地理学对阿根廷优先保护区域的识别进行了研究。保护生物地理学包括将生物地理学原理和方法应用于保护问题中。特有区域的识别对历史和保护生物地理学两者都很重要。原蛛下目蜘蛛扩散能力较弱, 具有有限的扩散机制和不爱活动的习惯。在这项研究中他们使用泛生物地理学方法 (通过追踪分析) 和最优性准则来分析其分布规律, 以确定阿根廷特有分布地区和优先保护地区。最终, 他们确定了7个普遍性足迹和4个生物地理节点[9]。
在系统发生生物地理学方面, 物种是生物研究的基本单位, 但以客观的方式确定物种的界限仍可能是高度主观和有偏见的。虽然形态常用来确定物种的界限, 但可用和流行的遗传数据使研究人员能够使用它为有关物种的界限作出推论。在其它方面的证据 (特别是形态) 可能低估物种的多样性的时候, 遗传数据也能用于隐匿物种 (Cryptic species) 的检测。Jason 等人采用加州暗门蜘蛛物种复合体微小暗门蛛 Aptostichus atomarius 作为模型系统, 提出一种客观的可重复的划定物种的方法, 即在该物种种群有分化且地理上高度结构化时, 通过寻找具有极端遗传分化和地理结构的该蛛类群中的种-群接口来划分内聚物种 (Cohesion species) 的综合方法。该新系统地理学的方法是一种普遍适用的方法, 也特别适用于高度结构化的系统, 它采用系谱排他性建立起一个拓扑架构, 来探讨在界定内聚物种的努力中的遗传和生态的可交换性。生境与基于生态位的分布模型两者的定性评价采用评估选择制度和遗传谱系间生态的互换性来进行;种群间的自适应分化比简单的地理一致性加权更重。根据这些分析, 他们可以得出的结论是, 该物种复合体被确认有5个内聚物种, 其中有3个在科学上是新的[10]。Jordan 等人探讨了一个复杂的形态保守的别的加洲暗门蜘蛛 (穴蛛科 Antrodiaetidae、异地蛛属 Aliatypus) 的物种界限。他们采用多种方法来确定该高度遗传碎片化类群的物种界限, 包括多位点的发现和验证的方法 (增加嵌合方法) 。此外, 还介绍了一种新的基于物种树的发现方法。结果表明, 该复合体包括多个隐匿物种。在采取保守的方法后, 确认它是1个三物种的复合体, 并正式描述了2 个新种 (罗塞异地蛛 Aliatypus roxxiae, sp.nov.和斯塔雷特异地蛛 Aliatypus starretti, sp.nov.) 。这项研究有助于阐明遗传碎片化类群中物种间的界限, 为识别和定义盛行于许多生物体类群中的隐匿谱系的多样性提供了一个框架[11]。
3、 大疣蛛的研究现状及意义
大疣蛛科Macrothelidae (原名:异纺蛛科 Hexathelidae) 隶属于蛛形纲 Arachinida 蜘蛛目 Araneae 原蛛下目Mygalomorphae。全世界现已记述大疣蛛科1属31种, 中国大陆已记录11种[2]。
在亚洲, 触形大疣蛛 Macrothele palpator Pocock, 190l 是唯一一种分布广泛的地穴型原蛛, 除对其形态特征进行了系统研究外, 其它工作开展有限。触形大疣蛛 M. palpator 原名触形粒突蛛。雌雄蛛体长10~20 mm。八眼密集, 前眼列端直或稍后曲, 后眼列后曲。下唇宽大于长, 密被黑色疣突。颚叶具大量疣突。足式:Ⅳ-Ⅰ-Ⅱ-Ⅲ。后中纺器不分节, 两纺器间的距离小于后中纺器长, 后侧纺器, 分三节, 末节指状[12]。触形大疣蛛 M. palpator 分布仅限于中国和中国香港地区, 在华东、华南和西南各省广泛存在[13] (见图1) 。
图1 触形大疣蛛Macrothele palpator (♀)

克里大疣蛛 M. cretica Kulczynski, 1903 (发现于克里特岛) 和环锤大疣蛛 M. calpeiana (Walckenaer, 1805) 的分布遍及东南伊比利亚半岛和北非的两个地方, 在欧洲它们受立法保护[14]。环锤大疣蛛 M. calpeiana 体大而笨重, 具简单生殖器, 常穴居, 与地栖生境有关。大疣蛛在石头下和树皮里结漏斗网且朝上或向外延伸, 形成片状的捕捉网[15]。虽然已经有人对它的形态变异[16]和生殖生物学[17]进行了研究。但该蛛分布仅限于小的地理区间, 它的分布被限制在直布罗陀和其主要的群山附近的范围内。环锤大疣蛛 M. calpeiana 适宜于炎热的夏天、温暖的冬天, 降水量相对较高, 它的庇护处需局部湿润的东风。它的圈养维护对温度和湿度要求的理解是至关重要的。这无疑与该蜘蛛某些生活史中的关键事件密切相关, 如卵囊的生产。环锤大疣蛛 M. calpeiana 被认为与栓皮栎森林的破坏和碎片化密切相关, 这是将该物种列入伯尔尼公约的原因。然而, 有些作者质疑这一观点。Miquel 等人在对其主要种群样品线粒体标记的分子系统发育进行分析的基础上, 从多方面检测了它的保护现状。数据分析表明环锤大疣蛛 M. calpeiana 分布零散, 且种群遗传分化水平高。作为一个谱系整体, 环锤大疣蛛 M. calpeiana 显然没有经历过种群的增长。对谱系年龄的估计表明, 环锤大疣蛛 M. calpeiana 是在墨西拿盐度危机时期迁入伊比利亚半岛的, 目前的种群碎片化起源于上新世及更新世。由于其进化具有独特性与奇异性, 它在欧洲已逐渐成为生物多样性的一个主要热点, Miquel等人推测, 其碎片化和种群深度的遗传分化, 支持其进入依法保护状态[14]。
4 、江西境内触形大疣蛛区系演化的形态定量聚类分析
日本学者在对东亚地区的七纺蛛亚科 Heptathelinae 进行地理区系研究时, 按外雌器特征将东亚各种七纺蛛分成5个类群, 类群间地理梯度隔离各异, 而类群内从西南朝向东北方向递进的痕迹却十分明显[18] (见图2) 。各种迹象表明, 原蛛下目的时空演化与东亚地质板块运动、各山系山脉走向与梯度变化以及生物地理区系的经纬度分布等地质环境因素密切相关。据此, 陈连水等采用形态定量的聚类分析方法对江西境内触形大疣蛛M. palpator区域性演化模式进行了研究, 以细化或核实上述观点。
图2 东亚七纺蛛亚科Heptathelinae各物种的分布
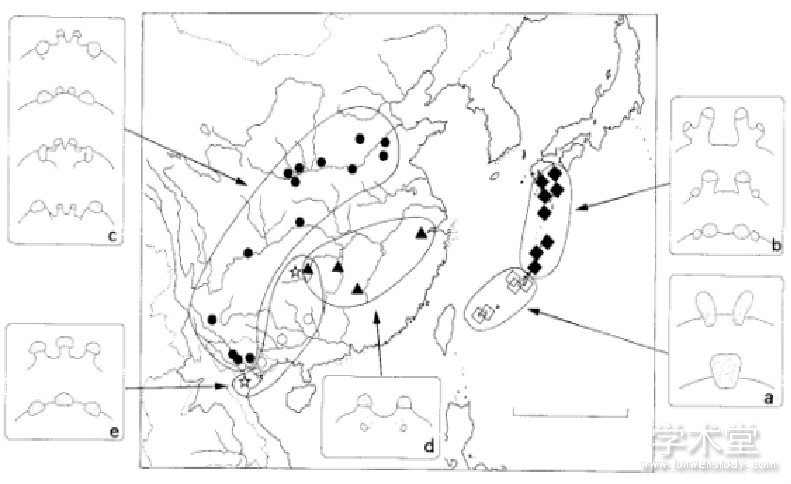
根据外雌器将各物种分成a~e的5个类群, 其中:“□”代表a类群;“■”代表b类群;“●”代表c类群;“▲”代表d类群;“☆”代表e类群 (引自C.Hyakunia等, 1999年)
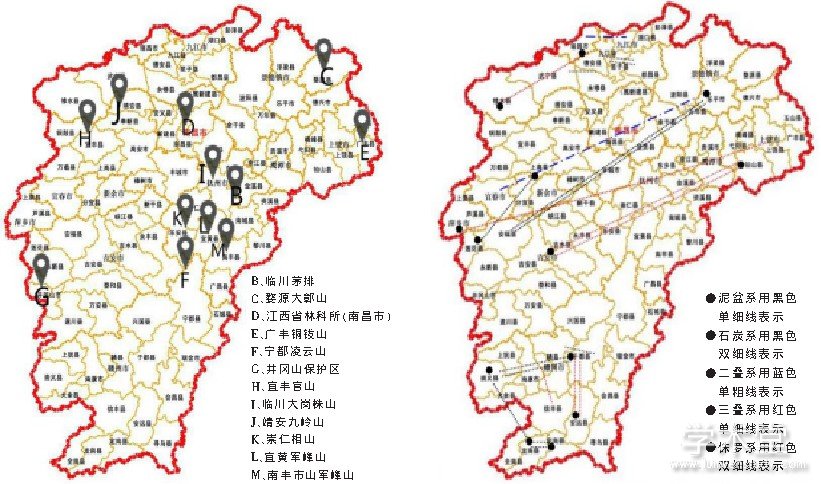
陈连水等从形态学角度出发, 采用各种显微测量工具测得了触形大疣蛛 M.palpator雌性成蛛Ⅰ步足长/总足长、Ⅱ步足长/总足长、Ⅲ步足长/总足长、Ⅳ步足长/总足长、Ⅰ腿足比、Ⅱ腿足比、Ⅲ腿足比、Ⅳ腿足比、体重/体积、前纺器/总长、后纺器/总长、唇颚疣突数比、唇长与宽比、前中眼距/前眼列宽、前中侧眼距/前眼列宽、后中眼距/后眼列宽、后中侧眼距/后眼列宽、前中眼宽/前眼列宽、前侧眼宽/前眼列宽、后中眼宽/后眼列宽、后侧眼宽/后眼列宽、头胸长/体长、腹长/体长、头胸宽/腹宽和后纺器长/体长 25 项主要形态分类特征的比值作为加权样本值进行类群定量聚类分析, 结果得到江西境内触形大疣蛛 M. palpator 地理演化支序图 (见图 3) 。将该支序图与江西地层图 (见图 4) 进行细致比对, 可发现上述触形大疣蛛 M. palpator 形态演化支序与江西泥盆系后地质构造运动所形成的地层地理区基本吻合。由此推测, 江西境内触形大疣蛛形态演化支序的形成受泥盆纪后地质构造运动影响, 其侵入线路可能是由赣西南到赣西再到赣中南, 真正演化中心在“湘赣岛海”的海区地貌区域, 即崇仁相山 K 支序;它经古鄱阳湖盆地断陷及新构造运动的漫长地层断陷、隆起、抬升和变质等地质作用, 再向赣中或赣中南四周扩散, 婺源独立支序 C 的形成可能与信江的地理隔断有关。显然它其中隐约存在从赣西南到赣中再到赣东北的形态地理演化方向的痕迹, 或者说, 对触形大疣蛛形态演化过程的追溯能一定程度上指示江西境内地质地层的发育和古生物的演化进程[19]。
图3 江西境内13个样点触形大疣蛛M. palpator形态定量聚类分析支序图
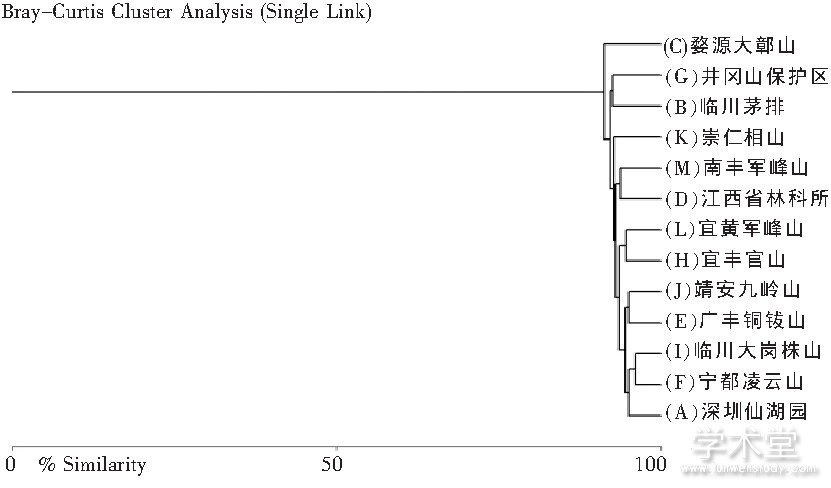
图4 江西境内触形大疣蛛M. palpator采集点与江西各时代地层地理区划示意图
5、 泛江西区域触形大疣蛛区系演化的形态定量聚类分析
为了更细致地研判, 使形态推定结果更有说服力, 陈连水等将触形大疣蛛 M. palpator 区系演化的形态定量聚类分析扩展到江西多地和周边、乃至该种整个分布区。近年来, 陈连水等在泛江西区域进行了补充采样, 增补了江西铅山、江西武夷山、江西贵溪阳际峰、江西分宜大岗山、江西九江庐山、江西崇义阳岭、福建武夷山、福州植物园、湖南长沙岳麓山、湘西白云山、湖北恩施星斗山等地雌蛛样本, 并增加了台湾大疣蛛、贵州大疣蛛和雷氏大疣蛛雌蛛样本及江西庐山白鹿洞雄蛛作为外群或对照样本。同前, 对这些样本的 25 项主要形态分类特征的比值进行测量、数据统计处理, 将前后共29个样本定量数据进行聚类分析, 得出泛江西区域触形大疣蛛 M. palpator 区系演化的形态定量支序图 (见图 5) 。
图 5 泛江西区域25 个样点触形大疣蛛M. palpator及外群形态定量聚类分析支序图
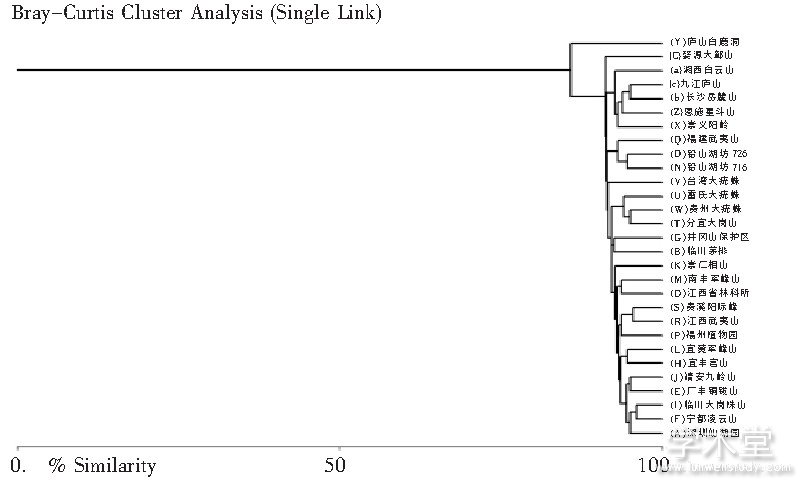
与中国中东部古地理图 (见图6) 进行比对, 由图 5 支序图可看出, 除对照的 (Y) 庐山白鹿洞雄蛛处于基部外, 从靠近基部的 (C) 婺源大鄣山独立支序到 (V) 台湾大疣蛛独立支序之间的 (a) 湘西白云山、 (c) 九江庐山、 (b) 长沙岳麓山、 (Z) 恩施星斗山、 (X) 崇义阳岭雌蛛样本形态相似度较高, 形成一个小集群;这一集群的形成或起源可能与泥盆纪前赣湘粤海湾与苏皖地区海侵融合及后来海退和古长江露头中保留的类蛛形动物有关[20]。而江西东北部的某些露头中的类蛛形动物在随后侏罗纪、白垩纪和第三纪泛江西区域东南盆地的形成中得以演化和向东南方向扩散, 形成了 (Q) 福建武夷山、 (O) 铅山湖坊 726 和 (N) 铅山湖坊 716 集群, 后由于浙闽高地的隆起而停止。随着第三纪中国南方地质构造运动的逐渐放缓和气候湿暖, 占据不同地域环境的大疣蛛种间分化加剧, 形成了 (V) 台湾大疣蛛、 (U) 雷氏大疣蛛、 (W) 贵州大疣蛛、 (T) 分宜大岗山种间支序集群[21]。进入晚第三纪和第四纪, 中国东南部触形大疣蛛的种系主要在湘赣区域的华南盆地中进行适应性辐射演化, 其主要原因是华厦系鄱阳湖构造运动形成的赣抚平原、崇仁盆地和玉山盆地及北北东断裂形成的抚河和信江等造成的[22]。由于金溪山字形运动, (K) 崇仁相山支序向东扩散, 这样就有图 5 中的 (S) 贵溪阳际峰和 (R) 江西武夷山支序, 后来新华厦构造运动促成武夷山的隆起, 加上东南山地的断裂形成了闽江, 闽江将武夷山中部资溪处的种系扩散到东南沿海的福州, 这样便形成了 (P) 福州植物园支序, 上述 (S) 贵溪阳际峰、 (R) 江西武夷山和 (P) 福州植物园就聚集成形态种内支序集群[23]。最后, 推测顶部的 (A) 深圳仙湖园支序是在第四纪由宁都凌云山等东江源区域的种系随东江断裂形成而扩散到深圳的[24]。
图6 中国中东部古地理图 (引自王鸿祯等, 1985)

(a) 晚三叠世; (b) 早侏罗世; (c) 早第三纪; (d) 中晚更新世
由此看来, 中国东部地区触形大疣蛛M. palpator种系源自于泥盆纪前东南盆地西北侧西南至东北向短暂相融形成的弧形海湾的多个侵入点, 同时, 伴随地质构造运动出现种间分化是发生于这段时期的主要事件;结合近期在缅甸琥珀中发现1亿年前有尾蜘蛛[25], 推测该蛛来源于西南方向的可能性更大;随着浙闽地区的抬升使保留于“湘赣岛海”的种系逐渐由西向东渗入, 因第三纪地质气候变迁归于平缓, 便出现了江西境内的适应性辐射演化;第四纪冰期后, 泛江西区域的触形大疣蛛 M. palpator 种系进入到深度分化和种群细分阶段。
6、 结束语
触形大疣蛛M. palpator是一种东亚热带、亚热带或温带地穴型原蛛, 也是原蛛下目唯数不多的广布性种类;因原蛛较为古老, 在地球上经历了长期的演化, 有证据显示它的遗传多样性背景的形成以及对地质环境的表观响应与地质地貌等变化相关。近年来, 经对江西境内各级自然保护区或多种矿区蜘蛛资源及区系分布的调查发现[26,27], 触形大疣蛛 M. palpator 是江西各保护区和矿区中常见地穴性原蛛, 用其作为地球地质等中长期环境变化的指示生物或许是一种最佳选择。
陈连水等对泛江西区域触形大疣蛛M. palpator地理区系的形态演化进行了研究, 结果表明泛江西区域各样点间触形大疣蛛M. palpator的地理演化呈分散的点线状, 存在从赣西南到赣中再到赣东北的形态地理演化方向的痕迹, 或者说, 对触形大疣蛛形态演化过程的追溯能一定程度上指示泛江西区域内地质地层的发育和古生物的演化进程。由于线粒体标记被广泛用于物种鉴定、种间进化关系、种内遗传结构和种群历史等各类研究中[28]。为进一步核实这个地理演化方向或明确其与泛江西区域地质地层发育的关系, 陈连水等拟综合采用形态特征比值、生态环境指数差与行为谱等定量聚类分析及与线粒体分子标记等系统发育相结合的方法对中国中东部该物种分布区内更多样点样本的遗传分化状况进行研究, 期待更真实的种群细分, 以破解其演化历程。
总之, 开展中国地穴型原蛛触形大疣蛛M. palpator (蜘蛛目:大疣蛛科) 种系发生与区系演化的研究定能为我国区域物种尤其是多姿多彩的无脊椎动物的微进化和生物地理学的研究添砖加瓦, 也能为有别于脊椎动物的更真实的无脊椎动物地理区系分布的划分提供更多实际案例和事实依据, 同时, 结合古地质、古气候、化石蜘蛛和分子遗传与进化生态学等新成果还能推动蜘蛛自身的系统发育学的研究以及蛛形与生态学更多深层次理论问题的破解, 如蜘蛛内聚物种和隐匿物种的界定、蜘蛛种系演化过程中遗传与生态因素的交换性、优先保护区的确定, 等等;或许作为新发现的一种环境指示生物, 它还能为该区域地质地层环境的中长期变化提供有效便捷的预警手段。
参考文献
[1] Wolfgang N.Spider Ecophysiology[M].Berlin:Springer-Verlag Berlin and Heidelberg GmbH & Co.K, 2013:3-4.
[2] Natural History Museum Bern.World Spider Catalog.Version 20.0[EB/OL]. (2019-1-11) .[2019-1-17].http://wsc.nmbe.ch.
[3] Paula S, David P.Fossil spiders[J].Biolgical Reviews, 2010, 85:171-206.
[4] Hafemik J E J.Threats to invertebrate biodiversity:implications for conservation strategies.In:Fiedler P L, Jain SK (eds) Conservation biology:the theory and practice of nature conserv-ation, preservation, and management[M].New York and London:Chapman and Hall, 1992:171-195.
[5] 申效诚, 张保石, 张锋, 等.世界蜘蛛的分布格局及其多元相似性聚类分析[J].生态学报, 2013, 33 (21) :6795-6802.
[6] Cox C B, Moore P D.Biogeography:An Ecological and Evolutionary Approach, 7th ed[M].Oxford:Blackwell Publishing Ltd, 2005:108-110.
[7] Penney D.Does the fossil record of spiders track that of their principal prey, the insects?[J].Earth Sciences, 2004, 94:275-281.
[8] Hannah M W, Nichols J M, Rosemart G G, et al.Treating fossils as terminal taxa in divergence time estimation reveals ancient vicariance patterns in the palpimanoid spiders[J].Systems Biology, 2013, 62 (2) :264-284.
[9] Nelson F, Alda G, Fermando P-M.Identification of priority areas for conservation in Argentina:quantitative biogeography insights from mygalomorph spiders (Araneae:Mygalomorphae) [J].Journal of Insect Conservation.2014, 18 (6) :1087-1096.
[10] Jason E B, Amy K S T.An integrative method for delimiting cohesion species:finding the population-species interface in a group of californian trapdoor spiders with extreme genetic divergence and geographic structuring[J].Systems Biology, 2008, 57 (4) :628-646.
[11] Jordon D S, Bryan C C, Marshal H.Multilocus species delimitation in a complex of morphologically conserved trapdoor spiders (Mygalomorphae, Antrodiaetidae, Aliatypus) [J].Systems Biology, 2013, 62 (6) :805-823.
[12] 朱明生, 李廷辉, 宋大祥.中国大疣蛛属 (蜘蛛目:异纺蛛科) 一新种[J].河北大学学报 (自然科学版) , 2000, 20 (4) :952-955.
[13] 徐湘.中国六疣蛛科和线蛛科的分类及应用RAPD技术于蜘蛛亲缘关系的初步探讨[D].长沙:湖南师范大学, 2001.
[14] Arnedo A M, Ferrandez M-A.Mitochondrial markers reveal deep population subdivision in the European protected spider Macrothele calpeiana (Walckenaer, 1805) (Araneae, Hexathelidae) [J].Conservation Genetics, 2007 (8) :1147-1162.
[15] Gallon R C.Observations on Macrothele calpeiana (Walckenaer, 1805) in southern Iberia[J].J Br Tarantula Society, 1994 (1) :1-12.
[16] Calvo-Hernández D, Santos-Lobatón M C.Variabilidad morfológica de las poblaciones de Macrothele calpeiana (Walckenaer, 1805) (Araneae, Hexathelidae) en la provincia de Cádiz (Espan?n?a) [J].Reviber Aracnol, 2001 (3) :43-45.
[17] Perry L.Captive breeding of funnelweb spider Macrothele calpeiana (Walckenaer, 1805) [J].J Br Tarantula Society, 2002, 17:113-121.
[18] Hvakunia C, Shinjuku K.Spider of the genus heptathela (Araneae, Liphistiidae) from vietnam, with notes on their natural history[J].The Journal of Arachnology, 1999, 27:37-43.
[19] 黄珍, 陈连水, 吴心怡, 等.江西境内触形大疣蛛 (蜘蛛目:异纺蛛科) 种系形态的区系演化[J].中国科技论文在线精品论文, 2015, 8 (19) :2067-2074.
[20] 崔克信, 刘椿, 潘云唐, 等.中国西南区域古地理及其演化图集[M].北京:地质出版社, 2004:78-155.
[21] 戎嘉余, 方宗杰, 周忠和, 等.生物的起源、辐射与多样性演变:华夏化石记录的启示[M].北京:科学出版社, 2006:383-398.
[22] 中国地质科学研究院.中华人民共和国地质图集[M].北京:地图出版社, 1974:66-73.
[23] 王鸿祯, 楚旭春, 刘本培, 等.中国古地理图集[M].北京:地图出版社, 1985:93-138.
[24] 马丽芳, 乔秀夫, 闵隆瑞, 等.中国地质图集[M].北京:地质出版社, 2002:202-208.
[25] Wang B, Dunlop J A, Selden P A, et al.Cretaceous arachnid Chimerarachne yingi gen.et sp.nov.illuminates spider origins.[2018-8-27].[J/OL].Nature Ecology & Evolution:614-622. (2018-2) https://doi.org/10.1038/s41559-017- 0449-3.
[26] 陈连水, 袁凤辉, 周谷春, 等.江西省蜘蛛资源的研究报告[J].江西科学, 2013, 36 (3) :175-178.
[27] 陈连水, 袁凤辉, 周谷春, 等.江西特色矿区蜘蛛资源分布及多样性比较研究[J].中国科技论文在线精品论文, 2015, 8 (7) :707-722.
[28] 刘名信, 彭作刚, 张志升.蜘蛛分子系统学研究进展[J].蛛形学报, 2012, 21 (2) :103-114.