动物学论文

摘 要: 为掌握喀斯特地区同域分布的小麂(Muntiacus reevesi)和毛冠鹿(Elaphodus cephalophus)共存的时空分化格局,利用公里网格布设的红外相机对贵州麻阳河国家级自然保护区内的这2种鹿类进行监测。于2019年2—8月,在151个网格位点分别获得1 328张小麂和167张毛冠鹿的独立有效照片,总有效工作时长21 234 d。结果表明:(1)在时间活动节律上,小麂与毛冠鹿均属晨昏活动型,均在7月活动强度最高,日和季节性活动节律上两者均无显着差异(P> 0.05),仅日活动高峰有所不同。(2)在生境利用分布上,小麂与毛冠鹿活动分布的重叠程度并不高,同期监测到两者的位点数仅占总监测位点数的22.11%。(3)在空间格局中(包括植被类型、海拔梯度、坡位等),两者活动强度均存在极显着差异(P <0.01)。研究结论表明,麻阳河同域分布的小麂与毛冠鹿主要以生境利用分化和空间分布差异方式共存,而时间分化差异不明显。研究结果对于进一步掌握喀斯特地区鹿科动物同域分布的共存维持机制及其资源保护与管理有重要意义。
关键词: 同域分布物种; 共存时空格局; 小麂; 毛冠鹿; 麻阳河保护区;
Abstract: We studied the spatiotemporal pattern of co-existence of two sympatric Cervid species Muntiacus reevesi and Elaphodus cephalophus at Mayanghe National Nature Reserve in the Karst area of Guizhou Province,China. We sampled by use of infrared cameras with the camera-layout on a km-grid. Over 21,234 camera working days we accumulated 1,328 independent photographs of Muntiacus reevesi and 167 of Elaphodus cephalophus from 151 camera sites. Based on these camera data, the results showed that:( 1) in terms of time activity rhythm, Muntiacus reevesi and Elaphodus cephalophus both were more active in morning and evening with activity peaks at different times,both were most active in July,and their daily and seasonal activity patterns were similar.( P > 0. 05);( 2) the extent of geographic overlap of the two species was not high,as shown by the number of camera sites detecting both Muntiacus reevesi and Elaphodus cephalophus in the same period only accounting for 22% of all camera sites;( 3) we conclude that the co-existence of these two sympatric Cervidae species was mainly due to their differentiation of habitat use and population distribution,rather than differentiation in timing of activity periods. Our study objectives were to further understanding of the co-existence and maintenance mechanism of sympatric species in karst areas,and to enhance conservation management of these species in this region.
Keyword: Sympatric species; Spatial temporal-patterns of co-existence; Muntiacus reevesi; Elaphodus cephalophus; Mayanghe Nature Reserve;

同域分布物种的共存机制一直是群落生态学的核心研究内容之一,根据竞争排斥原理,完全的竞争者不能共存,同域物种的长期稳定共存建立在种间的生态位分化上[1]。同域物种生态位的分化是多维度的,主要包括栖息地利用的分化、食性的分化、行为节律分化等[2]。对于相似近缘种而言,由于相似的生理需求与形态特征,甚至是行为特征,生态位的分化与重叠则可能更为显着地决定着其是否能够共存,以及以何种方式共存[3]。近年来,采用红外相机监测技术进行野生动物的活动节律和生态位分化的研究屡见不鲜,为探索同域物种共存机制提供了新的研究技术和方法[4,5]。比如,同为麂属(Muntiacus)的黑麂(Muntiacus crinifrons)与小麂(Muntiacus reevesi),主要是通过日活动高峰的错峰、活动海拔区间和植被类型的差异选择来实现二者的生态位分化,以实现在同一地区内长期共存[6]。但在南方山地中较为常见的鹿科(Cervidae)动物——小麂与毛冠鹿(Elaphodus cephalophus)的共存模式、生态位分化则尚无研究报道。
毛冠鹿在中国分布范围较为广泛,只产于中国,最适生存区在亚热带范围内[7],最西至西藏东部,南限在北回归线附近[8],最北可以到宁夏六盘山[9]。小麂主要分布在我国秦岭以南、云南哀牢山以北地区,属于中亚热带至北亚热带范围[10]。随着天然林保护成效显现,大型猫科(Felidae)动物缺乏等因素,这2种鹿科动物在贵州山地森林常常同域分布且均呈种群迅速增长趋势[11,12,13,14],然而关于这两种同域分布的鹿类是如何分化共存的?其生境利用与种群分布的空间格局和活动节律的时空格局是否存在明显差异?为回答这些问题,本研究选择贵州麻阳河国家级自然保护区,利用红外相机监测数据探讨了这2种鹿科动物活动节律的时空分化,以期为喀斯特地区同域分布物种的共存机制研究提供参考。
1、 研究地概况
贵州麻阳河国家级自然保护区(以下简称“麻阳河”或“保护区”,坐标为(28°37′—28°54′N,108°3′—108°20′E)位于黔北大娄山脉东北段,总面积为311.13 km2,以黑叶猴(Trachypithecus francoisi)及其栖息地为主要保护对象,是全球最大的黑叶猴自然种群的分布地,种群数量700余只[15],该保护区不仅以占全球种群数量1/3的黑叶猴着称,其他野生动物也十分丰富[11]。麻阳河保护区地跨沿河、务川两县,并与重庆接壤,是国家生态环境部颁布的“武陵山生物多样性保护优先区”内十分重要的国家级自然保护区。
麻阳河保护区属于典型的喀斯特地貌,区内海拔800—1 000 m以上地段地势开阔,而800 m以下则多为峡谷。保护区属于中亚热带湿润气候类型,雨量充沛、水热同季[16]。区内有2条主要河流,南部河流是麻阳河,北部河流是洪渡河及其支流兰字厂河、缠溪河等[15]。保护区内动植物资源丰富,已记录种子植物1 168种,鸟类198种,兽类44种,两栖爬行动物44种,鱼类55种,保护区总体森林植被覆盖率为65%[17]。保护区已记录的鹿科动物主要有小麂、毛冠鹿、赤麂(Muntiacus muntjak),其中以毛冠鹿和小麂较为常见[11,17],优越的自然条件为鹿科动物繁衍提供了丰富食物和生境资源。
2、 研究方法
2.1 、红外相机布设及数据处理
于2019年2—8月,利用1 km×1 km的网格(2个网格1台隔行交错布设,即每2 km2布设1个相机位点)对保护区全域布设红外相机(猎科LTL6210MC和东方红鹰E1B型号),共连续设置151个相机监测位点。通过ArcGIS 10.0对保护区全域进行公里网格区划及网格中心标注(理论相机位点),相机安装在各网格中心点周围20 m区域的适宜位置(人为干扰少、生境条件好、野生动物痕迹多样的兽径、水源地等),部分不能到达的区域、网格内人为活动干扰强烈或网格中心没有合适布设的区域,根据实际情况做出相应调整[11],但基本保证相邻2个网格间的2台相机之间的距离至少500 m[18]。相机安装过程中尽量减少人为因素对布设地点选择的影响,不投放任何诱饵,相机布设时注意隐蔽和伪装,根据地形调节相机使镜头基本与地面平行。进行红外相机的参数设置:照片+视频模式、间隔1 min、3张连拍、灵敏度中等,对相机进行试拍,核验正常后即详细记录像机编号及坐标、生境(植被类型、海拔、坡位等)、距居民区、水源、道路的距离等信息[4]。相机布设的详细方法、参数设置与监测记录程序等参照肖治术等[4]的研究。
使用Bio-Photo V2.1对红外相机监测到的照片按位点进行编号并生成Excel表[19],统计物种种类、名称、数量、性别、独立探测首张、干扰类型等信息并进行汇总整理,拍摄的每个视频看作单张照片进行统计。
2.2、 数据统计分析
2.2.1、 物种相对丰富度
物种相对丰富度(relative abundance index,RAI)的大小一定程度上代表着该物种在该域中的分布状况以及活动强度[20],以相对丰富度表征小麂和毛冠鹿在各监测位点和时段的活动节律:
RAI=(Ti/Ni)·100 %
式中:Ti表示i物种在某一分析因素(时间段、植被类型、海拔等)下的独立探测首张照片数,Ni表示i物种的独立探测首张照片数总数。
2.2.2 、活动节律重叠分析
为描述两物种的日活动节律及其重叠程度,利用核密度估计方法(kernel density estimation)分析小麂与毛冠鹿的日活动节律重叠模型[21]。该模型利用核密度估计方法,从小麂与毛冠鹿的独立有效探测照片采取随机样本,用以描述小麂与毛冠鹿的日活动节律特征。同时,比较2条不同物种的核密度曲线之间相似程度以估计不同物种日活动节律之间的对称重叠程度(⊿),其⊿取值范围从0(无重叠)到1(完全重叠),并比较小麂与毛冠鹿的日活动节律差异显着性,显着水平为P=0.05[22]。以上所有数据处理结果在R软件v3.2的“overlap”包中进行[23]。
2.2.3、 种间空间差异分析
采用Microsoft Office 2016中的Excel进行数据汇总整理。利用卡方Fisher's确切概率法分析小麂与毛冠鹿活动节律在生境类型(空间)上的分化差异[24],在SPSS 26软件中进行。运用ArcGIS 10.6中ArcMap进行小麂与毛冠鹿空间分布差异图绘制,采用Origin 2019b进行绘制雷达图,对小麂和毛冠鹿的空间分化差异进行分析比较。
3 、结果
于2019年2—8月间的整个监测期内共布设151个监测位点,得有效工作时长21 234 d,有78个位点监测到小麂活动,38个位点监测到毛冠鹿活动;其中小麂的独立探测首张照片为1 328张,毛冠鹿的独立探测首张照片为167张。
3.1、 活动节律的种间时间分化
日活动节律差异比较的结果显示(图1),小麂与毛冠鹿日活动节律无显着差异(⊿=0.85,P>0.05),日活动节律虽未完全重叠但大致相同,小麂与毛冠鹿的日活动节律高峰时段相近,昼间活动均近似“M”形,呈明显的“双峰型”。在日活动节律上,小麂主要在上午和傍晚活动,存在晨昏2个活动高峰(7:00—8:00和19:00—20:00),日出活动开始变得积极,13:00活动强度开始较快下降,直到18:00活动强度迅速增加,20:00活动强度急速下降,在20:00—6:00活动强度偏低。毛冠鹿在日活动节律中,早高峰比小麂推迟1 h左右,呈现出3个活动高峰(8:00—9:00,19:00—20:00、23:00—24:00),夜间除23:00—24:00为第3活动高峰外,活动相对较少,从6:00逐渐开始活跃,毛冠鹿总体在夜间(18:00—6:00)的活动强度高于小麂的夜间活动强度。
从季节性活动节律上看(图2),两者几乎相同,春末开始为较活跃时段,夏末活动强度迅速下降,活动高峰均在7月,小麂相对丰富度总和RAI=27.11,毛冠鹿相对丰富度总和RAI=29.94。卡方检验表明,两者在活动节律无显着差异(基于95%置信区间,χ2=14.024,P=0.052)。
3.2 、活动分布的种间空间分化
在布设的151个相机位点中,有95个位点监测到小麂或毛冠鹿的活动(各位点分布情况见图3,位点监测汇总见表1),其中有21个位点同时监测到小麂与毛冠鹿的活动,占监测到两物种位点总数的22.11%。在位点监测中,小麂与毛冠鹿的位点重叠程度并不显着,存在较大的点位分布差异。
图1 麻阳河国家级自然保护区小麂与毛冠鹿日活动节律差异比较
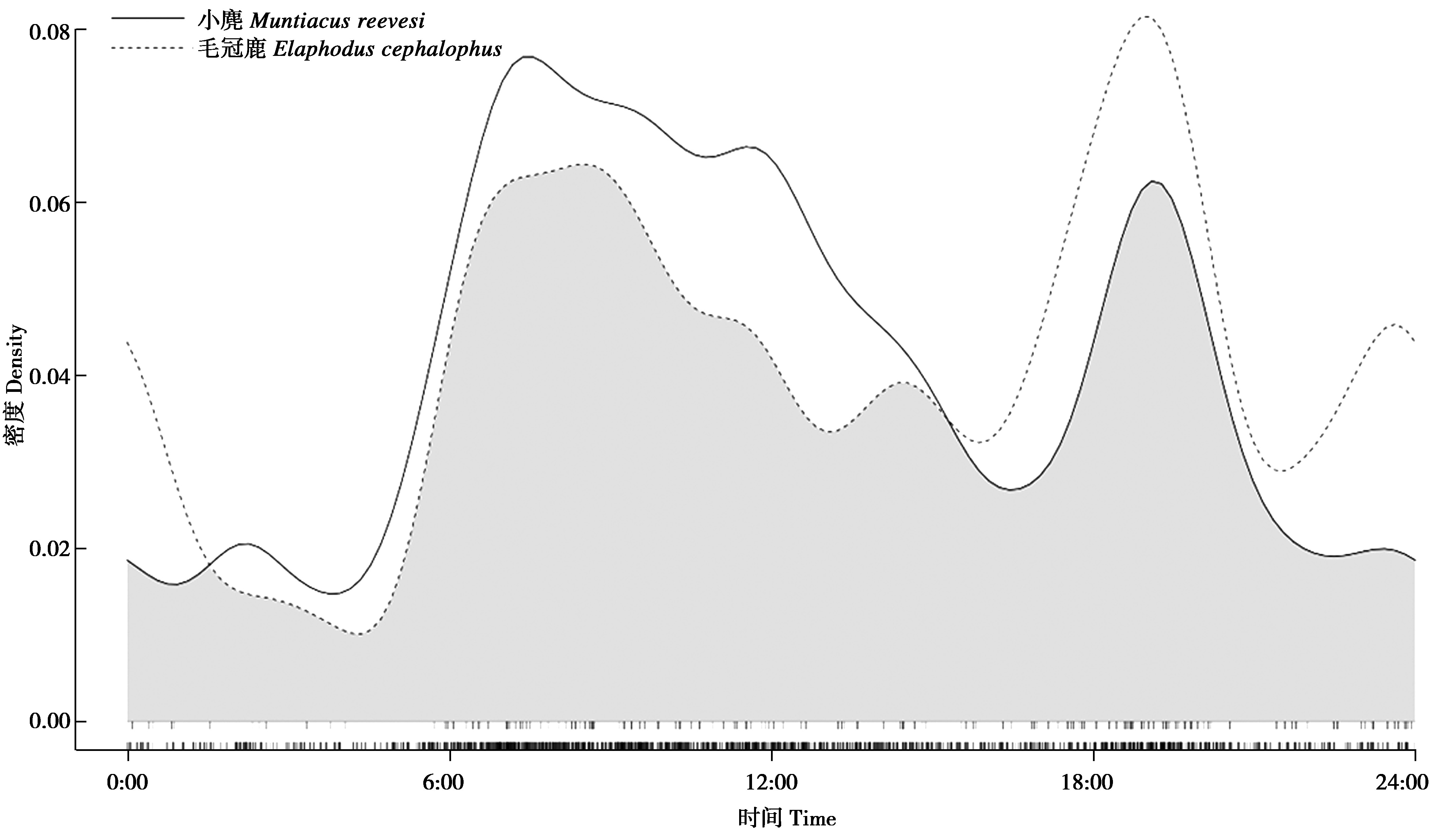
Fig.1 Comparison on daily activity rhythms between Muntiacus reevesi and Elaphodus cephalophus in Mayanghe National Nature Reserve,China
图2 麻阳河国家级自然保护区小麂与毛冠鹿季节性活动节律差异比较
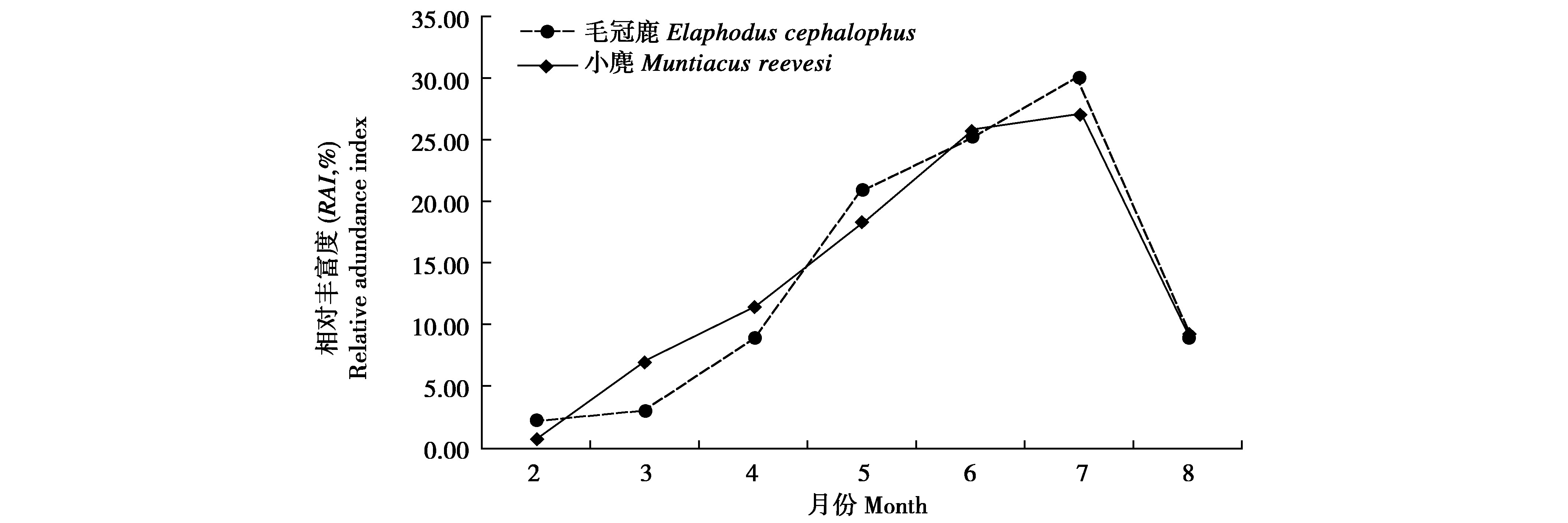
Fig.2 Comparison on seasonal activity rhythms between Muntiacus reevesi and Elaphodus cephalophus in Mayanghe National Nature Reserve
图3 麻阳河国家级自然保护区小麂与毛冠鹿红外相机位点探测分布多度与重叠图
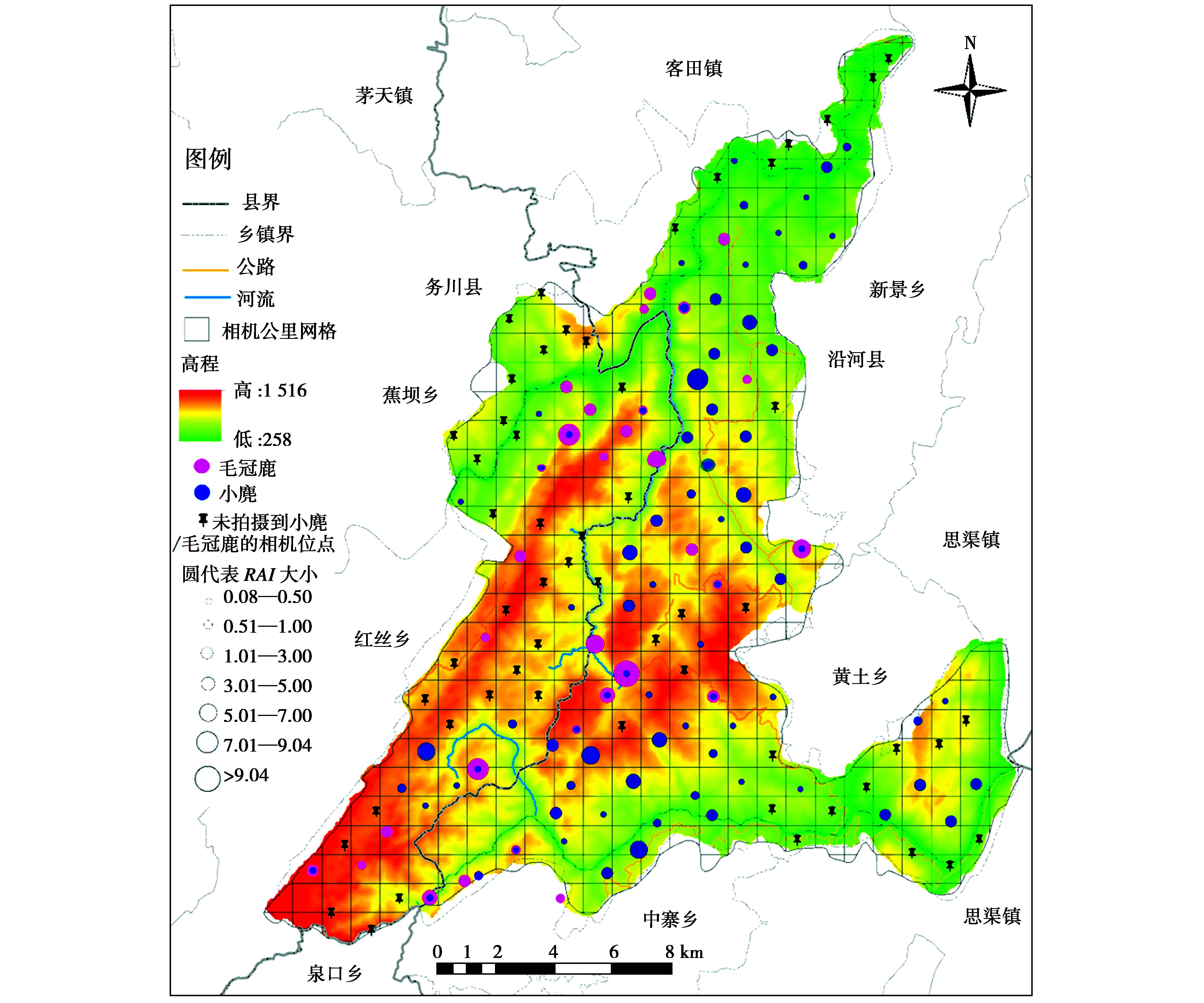
Fig.3 Abundance(RAI)and overlap of distributions of Muntiacus reevesi and Elaphodus cephalophus detected in each camera site in Mayanghe Nature Reserve
表1 麻阳河国家级自然保护区小麂与毛冠鹿监测位点有效照片数及相对丰富度指数(RAI)
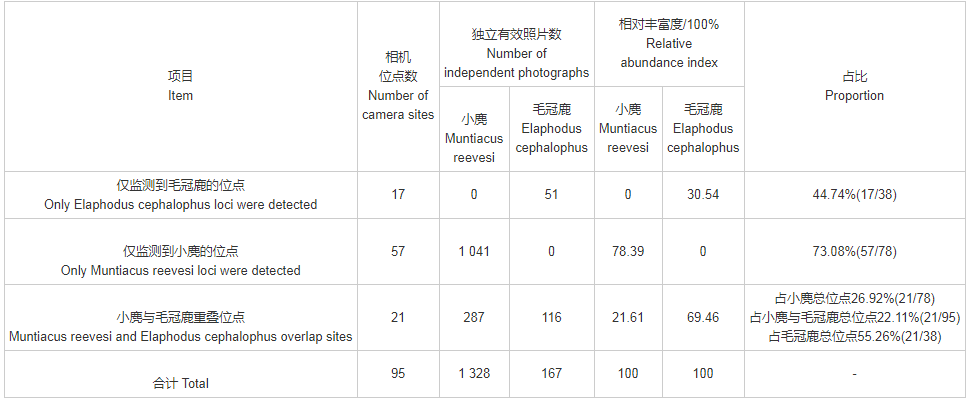
针对小麂与毛冠鹿所重叠的21个监测位点进行统计(表1,表2),分析同一位点拍摄到小麂和毛冠鹿的最近时间距离。在21个位点中,毛冠鹿的RAI平均值高于小麂的平均值,是其3倍之多;最近时间距的平均值较大,大于30 d,仅有一个位点是几乎同一时间监测到两者出现。
经Fisher's确切概率法检验,在所监测的位点中,小麂与毛冠鹿在植被类型上、坡位梯度上和海拔梯度上均存在极显着差异(P<0.01),二者存在明显的空间分布差异(图4)。
整体上看,在坡位差异上,小麂在中上坡位(总和RAI=34.19)、中坡位上活动频繁,而在下坡位相对丰富度总和最低(RAI=2.56),各坡位上相差并不太大。毛冠鹿虽也是在中坡位相对丰富度总和最高(RAI=70.06)、下坡位总和最低(RAI=0.60),但各坡位的差异较大。在植被类型上,小麂在针阔混交林中的相对丰富度总和最高(RAI=50.90),小麂在茂密的竹林之中也有所活动(RAI=0.15)。毛冠鹿在常绿阔叶林中的相对丰富度总和最高(RAI=38.32),在竹林中尚未监测到毛冠鹿的活动。在海拔梯度上,于海拔≤300 m、≥1 350 m间均未监测到有小麂与毛冠鹿的活动,根据当地地形情况,按200 m高差分级进行统计(图4C)。在300—500 m的河谷低海拔区仍有毛冠鹿少量活动但未发现小麂,两者均主要分布在500—1 300 m区段,毛冠鹿在此区域各区段内活动较为均匀,活动范围内海拔最高达到1 347 m(RAI=1.20),而小麂则主要在900—1 100 m的狭窄范围内(RAI=53.29)活动频繁。
表2 麻阳河国家级自然保护区小麂与毛冠鹿重叠位点最近时间距汇总
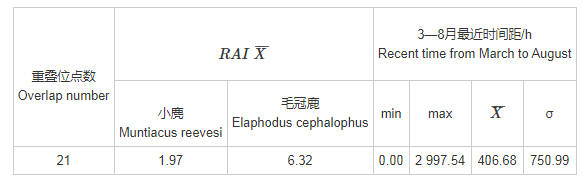
图4 麻阳河国家级自然保护区不同生境类型中小麂与毛冠鹿相对丰富度(RAI)雷达图

Fig.4 Radar chart for relative abundance index(RAI)of Muntiacus reevesi and Elaphodus cephalophus in different habitat types detected by cameras in Mayanghe National Nature Reserve
4 、讨 论
竞争排除法则(即高斯假说,Gause's hypothesis)的生态位分化理论常被认为是物种共存遵守的基本法则[1],但共存种间如何分化却不尽相同。显然,掌握同域分布物种的行为活动在空间和时间上的分化,从群体行为上判断其生态位分化,是揭示其共存机制的重要途径。
行为活动的时间差异常常是共存物种的生态位分化形式之一[25]。活动节律既是动物本身进化适应的一部分,又是对环境因素变化采取生存对策的一种适应[26,27],在长期的自然选择中,每一个物种都形成了利于自己的最适活动节律[28]。毛冠鹿和小麂属同科不同属近缘种,但繁殖节律却并不相同。毛冠鹿多在10—11月发情交配,4—5月产仔,而小麂在贵州则全年可发情繁殖[29],但在季节性活动节律上,两者却不是在繁殖期呈现最活跃状态,而是均在6—7月呈现一年中最活跃的状态,这可能与当地的气温和物候有关。麻阳河保护区7月达到年度气温最高(均温27.2℃)[30],因此可能在6—7月由于中午较炎热,而导致2种动物中午的活动减少,而在早晨和黄昏的活动频繁度增加。6—7月正是麻阳河春季及初夏嫩枝叶期已过,尚未结实的间隔期,而主要木本植物种(麻阳河阔叶植物群落主要为栓皮栎Quercus variabilis、丝栗栲Castanopsis fargesii等壳斗科Fagaceae植物)多开始于8—9月结实[17],也可能是此时期适口性和营养性的食物减少,导致活动频度增加。
在日活动节律上,小麂与毛冠鹿日活动节律虽未完全重叠却大致相同,无显着差异(⊿=0.85,P>0.01),但小麂早高峰比毛冠鹿相对提前了1 h,且毛冠鹿存在第三高峰(23:00—24:00)。这种活动高峰的时间偏移,使得这两种食性相似、取食部位有所不同的同域分布的物种出现了生态位的偏离,从而降低了二者间竞争的激烈程度[31]。两物种的活动节律可能与当地的食物丰度、人为干扰、气候环境等均有一定关系[32],本区域的小麂与毛冠鹿的时间节律与贵州宽阔水的小麂[13](小麂为晨昏型,存在 9:30—11:30 、 17:00—19:00 两个高峰)、贵州习水的毛冠鹿[12](毛冠鹿主要在傍晚活动,18:00—20:00)的时间节律存在较大差异。
在物种生境利用的种群分布分化上,在95个监测位点中仅有21个均出现两者重叠现象,表明两物种的分布的生境重叠度并不高,但并非完全没有重叠,在其中1个位点还出现两者仅间隔22″同时出现的情况。但大多数情况下,两者并不重叠,多数相机位点两者出现的时间平均相差30 d以上,从RAI分布图(图3)上,也可看出,两者的重叠度并不高。在生境利用的差异上,麻阳河的毛冠鹿与江西武夷山的研究结果相似,喜在常绿阔叶林和针阔混交林中活动[33];而小麂主要在针阔混交林中活动,这可能与针阔混交林中植被更新较快、植物物候期多样、能提供更多小麂喜食的嫩芽嫩叶有关[34]。食物资源常常是驱动生态位分化的主要因素[35]。根据以往报道,这两种鹿科动物食性相似性较大,二者均喜食百合科(Liliaceae)、禾本科(Poaceae)、蔷薇科(Rosaceae)、鸢尾科(Iridaceae)等草本植物以及杜鹃花科(Ericaceae)、壳斗科(Fagaceae)等木本植物和部分藤本植物的新鲜组织[7,29,36],而在麻阳河这类植物资源丰富且分布较广泛[17],可为小麂和毛冠鹿提供充分的食物来源,因此是否食物资源差异驱动空间分化仍有待深入研究。在海拔上,小麂和毛冠鹿均在900—1 100 m达到活动强度最高峰,但在900 m以下的海拔地区小麂的活动强度明显高于毛冠鹿,说明小麂在麻阳河的低海拔地区仍有较大种群分布。在中海拔地区,小麂与毛冠鹿活动强度相似,重叠拍摄的机位也主要在这一地段,因此可能两者存在一定的生态位重叠,总体而言,在麻阳河,毛冠鹿在海拔范围上的占域大于小麂,本域海拔1 100 m以上的地区坡度较大,且针阔混交林减少,毛冠鹿在较高海拔的相对丰富度不高,活动相对较少。在坡位的差异上,小麂的活动强度最高峰出现在中上坡(RAI=34.19),毛冠鹿的活动强度最高峰在中坡位(RAI=70.06),这可能与小麂更适于坡度较陡的区域有关。同时受研究方法影响,我们主要从相机位点拍摄情况及所处生境角度分析两者群体分布和生境利用分化,至于两者更细致的生境选择偏好差异,仍有待进一步深入研究。
综上,可见麻阳河的小麂与毛冠鹿在空间分布和生境利用上存在显着差异性,这种差异性分化将减少小麂与毛冠鹿相遇的可能性,从而产生空间上的分离以维持共存,这与吕忠海等[37]对同为鹿科动物的东北马鹿(Cervus elaphus xanthopygus)和西伯利亚狍(Capreolus pygargus)的同域分布研究结果相似,但毛冠鹿与小麂的亲缘关系则要比马鹿和狍近得多[38],而两者在活动节律上并无显着差异,因此,可能表明同域分布的近缘鹿科动物主要以空间利用分化的形式维持共存,而不是在活动时间分化上。根据本研究结果,我们建议在自然保护区的资源保护与巡护管理中,要充分注意两种物种的生境利用区别,注重生境的差异性保护管理及物种高峰活动期的人为活动干扰控制,避免两者因各自主要分布范围内的生境丧失和退化,造成交叠空间竞争加剧,从而更好地促进其共存和繁衍。
参考文献
[1] GAUSE G F.The struggle for existence[M].New York:Hafner,1934.
[2] 储诚进,王酉石,刘宇,等.物种共存理论研究进展[J].生物多样性,2017,25(4):345-354.
[3] 夏珊珊,胡大明,邓玥,等.同域分布红腹锦鸡和红腹角雉在不同空间尺度下的生境分化[J].生态学报,2019,39(5):1627-1638.
[4] 肖治术,李欣海,王学志,等.探讨我国森林野生动物红外相机监测规范[J].生物多样性,2014,22(6):704-711.
[5] 陈立军,束祖飞,肖治术.应用红外相机数据研究动物活动节律——以广东车八岭保护区鸡形目鸟类为例[J].生物多样性,2019,27(3):266-272.
[6] 戚伟尧,陈智强,王芳,等.遂昌牛头山两种麂属动物的生态位分化[J].丽水学院学报,2020,42(2):33-38.
[7] 盛和林,陆厚基.毛冠鹿的分布、资源和习性[J].动物学报,1982,28(3):307-311.
[8] 宋胜利,宋文辉,史文清,等.中国药用鹿科动物简述[C]//中国畜牧业协会,辽宁省外经贸厅,辽宁省服务业委员会,等.第六届(2015)中国鹿业发展大会论文汇编.辽宁铁岭,2015:34-41.
[9] 高惠,马忠其,滕丽微,等.宁夏回族自治区六盘山国家级自然保护区发现毛冠鹿[J].动物学杂志,2017,52(6):1066-1068.
[10] 鲁庆彬,于江傲,高欣,等.冬季清凉峰山区小麂和野猪的生境选择及差异[J].兽类学报,2007,27(1):45-52.
[11] 杨雄威,吴安康,邹启先,等.贵州麻阳河国家级自然保护区红外相机鸟兽监测[J].生物多样性,2020,28(2):219-225.
[12] 穆君,王娇娇,张雷,等.贵州习水国家级自然保护区红外相机鸟兽监测及活动节律分析[J].生物多样性,2019,27(6):683-688.
[13] 粟海军,李光容,陈光平,等.贵州宽阔水自然保护区野生动物红外相机调查初报[J].兽类学报,2018,38(2):221-229.
[14] 张明明,杨朝辉,王丞,等.贵州梵净山国家级自然保护区鸟兽红外相机监测[J].生物多样性,2019,27(7):813-818.
[15] 牛克锋,肖志,王彬,等.中国麻阳河国家级自然保护区黑叶猴种群数量估计与分布[J].动物学杂志,2016,51(6):925-938.
[16] 李仕泽,吕敬才,李灿,等.麻阳河国家级自然保护区两栖爬行动物资源调查[J].动物学杂志,2015,50(1):59-67.
[17] 苟光前,魏鲁明,谢双喜.贵州麻阳河国家级自然保护区生物多样性研究[M].贵阳:贵州科技出版社,2017.
[18] 李佳琦,徐海根,万雅琼,等.全国哺乳动物多样性观测网络(China BON-Mammals)建设进展[J].生态与农村环境学报,2018,34(1):12-19.
[19] 李晟,王大军,肖治术,等.红外相机技术在我国野生动物研究与保护中的应用与前景[J].生物多样性,2014,22(6):685-695.
[20] 武鹏峰,刘雪华,蔡琼,等.红外相机技术在陕西观音山自然保护区兽类监测研究中的应用[J].兽类学报,2012,32(1):67-71.
[21] RIDOUT M S,LINKIE M.Estimating overlap of daily activity patterns from camera trap data[J].Journal of Agricultural,Biological,and Environmental Statistics,2009,14(3):322-337.
[22] ZAR J H.Biostatistical analysis[M].5th ed.New Jersey:Prentice-Hall,2010.
[23] MEREDITH M,RIDOUT M.Overlap:estimates of coefficient of overlapping for animal activity patterns.R package version 0.2.4.[CP/OL].[2020-10-22].http://h-s.cran.r-project.org.forest.naihes.cn/.
[24] 刘小斌,韦伟,郑筱光,等.红腹锦鸡和红腹角雉活动节律——基于红外相机监测数据[J].动物学杂志,2017,52(2):194-202.
[25] 牛克昌,刘怿宁,沈泽昊,等.群落构建的中性理论和生态位理论[J].生物多样性,2009,17(6):579-593.
[26] 刘宇,缪华,蒋桂莲,等.云南古林箐省级自然保护区四种哺乳动物日活动节律及活动模式分析[J].野生动物学报,2019,40(4):832-839.
[27] 李生强,汪国海,施泽攀,等.红外相机技术监测喀斯特生境兽类和鸟类多样性及活动节律[J].兽类学报,2016,36(3):272-281.
[28] 王渊,初红军,韩丽丽,等.基于红外相机陷阱技术的卡拉麦里山有蹄类自然保护区狼(Canis lupus)的活动节律[J].干旱区研究,2014,31(4):771-778.
[29] 罗蓉.贵州兽类志[M].贵阳:贵州科技出版社,1993.
[30] 崔洪涛.贵州省4个黑叶猴(Trachypithecus francoisi)分布的自然保护区栖息地质量评估[D].贵阳:贵州师范大学,2019.
[31] 崔鹏,康明江,邓文洪.繁殖季节同域分布的红腹角雉和血雉的觅食生境选择[J].生物多样性,2008,16(2):143-149.
[32] 刘振生,王小明,李志刚,等.贺兰山岩羊不同年龄和性别昼间时间分配的季节差异[J].动物学研究,2005,26(4):350-357.
[33] 程松林,林剑声.江西武夷山的毛冠鹿[J].动物学杂志,2011,46(5):40.
[34] 贾晓东,刘雪华,杨兴中,等.利用红外相机技术分析秦岭有蹄类动物活动节律的季节性差异[J].生物多样性,2014,22(6):737-745.
[35] 昶野,张明明,刘振生,等.贺兰山同域分布岩羊和马鹿的夏季食性[J].生态学报,2010,30(6):1486-1493
[36] 盛和林,王培潮.小麂的生态和利用[J].动物学杂志,1976,11(1):39-40.
[37] 吕忠海,冯源,于沿泽,等.高食性重叠度对同域物种生境分离的影响——以马鹿和狍为例[J].东北林业大学学报,2020,48(2):72-75.
[38] SMITH A T,解焱.中国兽类野外手册[M].长沙:湖南教育出版社,2009.