动物学论文


空间学习与记忆在动物的生命历程中扮演着非常重要的角色,它使动物能够记住自身所处的环境特征和路线,或者更进一步,在大脑中形成认知地图,从而帮助动物更高速准确地向目的地移动或获得目标物,动物的空间学习与记忆能力,可帮助其在返巢、觅食及避敌等关键行为中获得最大利益,并通过对其生存环境的空间认知,调整自己的行为模式,从而最大限度地保证自身在自然环境中的生存。
动物的空间认知能力越来越受到行为学家的重视,大量的研究结果显示,动物的空间学习和记忆能力受到遗传、内分泌、环境及个体经历等多个因素的影响。为了在变化的环境中获得准确的空间信息,从而建立起正确的认知地图,动物在空间学习过程中会采用2种不同的认知策略:以体轴为中心的左右转向策略和感觉线索提示。采取前一种策略的动物是通过记住一系列的左转或右转来定位目标,采取后一种策略的动物是通过视觉、嗅觉或触觉等的提示来找到目的地。研究结果显示,哺乳动物能够同时使用2种策略,并会根据环境不同优先选择某一种认知策略。
对于动物空间认知能力的研究结果大多来自于哺乳类,而以鱼类为对象的研究较少。已有的研究结果显示,鱼类同样具有复杂的空间认知能力,我们有理由相信,鱼类也会根据环境的不同采取不同的认知策略。鳑鮍鱼类具有特殊的生殖行为,在繁殖季节,雄性个体守护河蚌并吸引雌鱼来河蚌中产卵,雌鱼对雄鱼及河蚌的质量进行考察并最终选择某一个河蚌作为其产卵位点。本研究使用这种小型淡水鲤科鱼类———鳑鮍为对象,利用T形迷宫装置,研究在不同的环境中(有一个标记物vs无标记物),通过训练的鳑鮍鱼类能否完成空间学习和记忆的任务,并进一步比较其对2种不同认知策略的使用,以期为我国鱼类认知行为学研究提供资料。
1 材料与方法
1.1 试验鱼的采集与饲养选择16尾性成熟、体形完整、身体健康的雌性高体鳑鲏为实验对象(为避免性别差异的影响,选择单一性别个体进行本试验)。本试验使用的高体鳑鲏均采自湖北省大冶市保安湖。所采集的试验鱼饲养在具有充氧设备的大型鱼缸中。在整个实验过程中使用加热棒保证鱼缸的水温维持在约20℃左右。
同时人工控制光周期,保证每日有光照与无光照的时间比为12∶12。在饲养期间,每天给试验鱼提供充足的冻孑孓。
1.2 试验装置本试验采用“T”形迷宫设计。由灰色不透明塑料制成十字形实验缸(图1)。每个臂长30cm,宽15cm,高30cm。如果利用隔板封闭任意一个臂,其他3个臂就会形成“T”形迷宫。试验过程中,距左右两臂末端15cm处放置可移动的活动门(15cm×20cm),活动门下方中央开有长5cm,宽3cm的洞口。将挡板升起,则试验鱼可以通过活动门中央洞口进入两臂的末端。将挡板放下,则试验鱼将被限制在臂末端的空间中。【1】
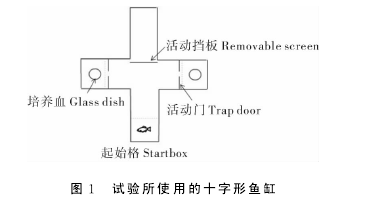
1.3 试验过程试验过程由预备实验、记忆获得实验和记忆验证实验三部分组成。
1)预备实验。为了使试验鱼熟悉实验缸的环境,进行了3次预备实验。预备实验阶段,在迷宫两臂末端的活动门后各放置一个装满冻孑孓的培养皿。将16尾试验鱼随机分为2组,以组为单位进行预备实验。每次实验开始,将同组的8尾试验鱼同时放入迷宫起始格(起始格随机选择),同时撤除左右两臂的活动挡板。在试验鱼适应5min后,撤去起始格的活动挡板,则试验鱼可通过洞口游出起始格,并在整个T形迷宫中自由游动及摄食冻孑孓。
试验鱼在实验缸中停留4h后,转移回饲养缸,该次预备实验结束。试验鱼只有在实验缸中才能获得食物,被移回饲养缸后将不能再取食。其中1组试验鱼(无路标组)在进行预备实验时,实验缸中除培养皿外不放置任何可见标记物,另1组(有路标组)则在实验缸中放置一可见路标(路标为绿色人造水草,无气味)。全部预备实验结束后,16尾试验鱼被分别编号并以小型鱼缸单独隔离饲养,断食3d后,开始进行记忆获得实验。
2)记忆获得实验。记忆获得实验共进行8d,每天对试验鱼进行3次训练,每尾试验鱼共计训练24次。训练过程中,与预备实验阶段相同,T型迷宫的左右两臂末端仍各放置一个装有冻孑孓的培养皿,但是只有一个培养皿中的冻孑孓可供试验鱼取食(可获得的食物报偿),另一个培养皿中的冻孑孓以消毒纱布包裹,以排除嗅觉对试验鱼方向选择的影响。另外,为了测试试验鱼是否存在方向偏好性,在对无路标组试验鱼进行训练时,我们将一半试验鱼可获得的食物报偿固定在T型迷宫左臂末端,另一半则固定在右臂末端,比较试验鱼对左、右两边的选择是否存在显着性差异。在对有路标组的试验鱼进行训练时,我们将可见标记物(绿色人工水草)放置在与可获得的食物报偿端一致的转角处。该组试验鱼的训练过程中,食物报偿端的方向会随机选择,则可见标记物的放置方向也会相应变动,从而避免试验鱼对固定的左右方向形成记忆。每次训练时,随机选择十字型实验缸的一端作为起始格,封闭起始格对向臂,形成T型迷宫。将试验鱼放入起始格适应5min后,升起起始格挡板,允许试验鱼从中央洞口自由游出,并记录试验鱼从离开起始格至进入食物臂末端所消耗的时间、试验鱼第1次所选择的方向以及运动轨迹(如折返、两臂间往返、停顿等)。在试验鱼成功取食冻孑孓或离开起始格10min后,本次训练结束。每天3次训练完成,将试验鱼转移回原小型鱼缸。记忆获得实验过程中,试验鱼只有在T形迷宫中可以得到食物。
3)记忆验证实验。在记忆获得阶段最后6次训练过程中,对每尾试验鱼穿插进行了3次验证实验,以检验本试验中的16尾试验鱼经过训练后对方向的记忆情况。本阶段的基本实验操作和数据记录与记忆获得阶段相同,但实验缸设置有所改变:对每尾试验鱼进行验证实验时,实验缸中均不放置培养皿及可见路标。
1.4 试验数据的采集
1)消耗时间。即记录试验鱼离开起始格到进入食物臂末端所花费的时间,s。
2)正确率。即实验对象对方向选择的正确情况。以数值1或0标示试验鱼首次选择方向的正确与否,若试验鱼第1次就选择并且进入了正确端,则记为最大值1;若试验鱼第1次就选择进入了错误端,则记为最小值0。同时记录试验鱼在实验缸中的运动轨迹,以数据加权的形式对每次选择的正确率进行处理。
3)习得次数。为比较不同路标条件对高体鳑鲏形成稳定记忆的影响差异,制定了习得标准:在记忆获得实验阶段,如果实验对象在连续10次选择中有6次选择了正确的方向,则认为其获得了稳定记忆,记录该对象的习得次数。已经获得稳定记忆的高体鳑鲏继续参与训练直到完成24轮实验。
1.5 试验数据的统计分析采取成对t检验对无路标组受控选择不同方向的个体的选择情况进行比较,两者间没有显着差异,表明试验鱼没有方向偏好性,所以我们将2组数据合并进行分析。
使用Excel软件统计消耗时间和正确率的平均值。使用Spss19.0软件对试验数据进行相应的统计分析。消耗时间和正确率分析采用t检验。
2 结果与分析
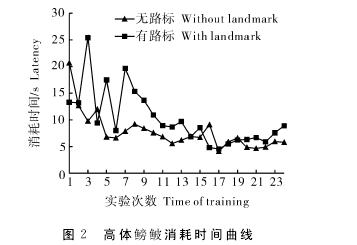
2.1 试验过程中消耗时间变化在2种不同的实验条件下,高体鳑鲏到达其目标位置所消耗的时间变化曲线见图2,从图2中可以看出,无路标条件下和有路标条件下,时间消耗随着实验进程均表现出明显下降的趋势。对无路标组前3次记忆获得实验消耗时间均值与最后3次记忆获得实验消耗时间均值进行配对t检验,t=1.651,P=0.143,未表现出显着性差异;对有路标组前3次记忆获得实验消耗时间均值与最后3次记忆获得实验消耗时间均值进行配对t检验,t=3.972,P=0.005,表现出显着性差异。从图2可以看出,2组试验鱼在前期训练中表现出的消耗时间存在较大差异,甚至有路标情况下消耗时间更长,该差异是由每组试验对象的个体活动性不同所引起的,但从获得稳定学习记忆后消耗时间变化趋势可以看出,随着试验鱼获得稳定的记忆及对实验环境熟悉度的加大,这种个体差异的影响基本已经消失了,两组的消耗时间变化趋势平稳。对无路标组和有路标组的最后3次实验消耗时间均值进行比较,t=0.864,P=0.402,两组间未表现出显着性差异。【2】
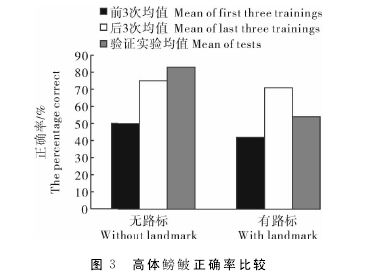
2.2 试验过程中正确率变化对试验鱼在记忆获得阶段前3次正确率均值、记忆获得阶段最后3次正确率均值及验证实验正确率均值进行了统计,不同路标条件下的正确率比较见图3。图中显示,经过24轮记忆获得实验阶段的训练,在无路标组和有路标组,实验对象的正确率相对训练前都有所增高。对无路标组前3次和后3次正确率均值进行比较,t=-1.549,P=0.196,未表现出显着性差异;对有路标组前3次和后3次正确率均值进行比较,t=-4.950,P=0.008,表现出显着性差异。【3】
2.3 不同路标条件下形成稳定记忆所需实验次数比较对每尾参与试验的高体鳑鲏达到学习标准所需的习得次数进行了统计,16尾个体均在24轮训练中达到了制定的学习标准,其中无路标组的习得次数最少的为15次,最多的为23次,有路标组的习得次数最少为10次,最多为16次。对2组实验对象的习得次数进行比较,t=5.528,P=0.000<0.05,两者表现出显着性差异。
3 讨论
3.1 高体鳑鲏学习与记忆的表现早在1978年,O'Keefe等就指出,动物能够通过日常的活动,收集自身所处环境的空间信息,从而在大脑中形成“认知地图”。“认知地图”是动物对其身处环境的空间格局产生的神经反映,它有助于动物在较复杂的环境条件中行使一定的行为,以保证其生存及繁殖。如要快捷、高效地完成觅食、避敌以及返回栖息地等基本行为,都要求动物对当前环境的空间布局相当熟悉。不同环境提供给动物的空间信息是不同的。环境直接影响动物对空间问题的解决。近年来,越来越多的行为学者对鱼类的空间学习与记忆的能力表现出极大的关注。
Williams等以斑马鱼为实验对象所作的有关空间转换任务的研究、Odling-Smee等以三刺鱼为实验对象所作的不同地理种群的认知策略研究、Croy等以斑马鱼为实验对象所作的对不同颜色标记物的联想学习,都显示出鱼类具有很好的空间认知能力。
我们对高体鳑鲏的研究也显示出相同的结果,无路标组前3次记忆获得实验消耗时间均值与最后3次记忆获得实验消耗时间均值比较,t=1.651,P=0.143,正确率比较,t=0.-1.549,P=0.196,虽在时间和正确率变化上未表现出显着性差异,但消耗时间曲线却呈现出明显下降趋势,同时正确率相对试验前明显提高。有路标组时间和正确率的比较结果分别为:t=3.972,P=0.005和t=-4.950,P=0.008,两者在训练前后均表现出显着性差异。结果表明经过训练的高体鳑鲏,能够依赖自身方向感或可见标记物定位,从而更快更准确地找到食物报偿的位置。反映出高体鳑鲏具有良好的空间认知能力,在不同的环境条件下,它们都能够根据感觉的或者环境的线索,建立对食物报偿的联想性记忆。
3.2 标记物对高体鳑鲏学习与记忆的影响在有可见标记物存在的环境中,动物将可见标记物作为定向标志,在无可见标记物存在的环境中,动物也可依赖自身的左右方向感定位,这是2种不同的认知策略。随机分组的高体鳑鲏,在无标记物存在的条件下,达到学习标准的习得次数为15~23次,在有路标存在条件下,习得次数为10~16次,2种不同环境下的习得次数比较,t=5.528,P=0.000<0.05,两者表现出显着性差异。结果显示,不同条件下表现出的习得次数的显着差异,可能揭示了不同的认知策略会影响高体鳑鲏稳定记忆的形成速度,在有可见标记物的条件下,高体鳑鲏能更快形成稳定的空间认知,路标的视觉提示作用能够起到比自身方向感更强烈的刺激记忆形成的作用。对2组实验对象记忆获得后的消耗时间进行比较,t=0.864,P=0.402,未表现出显着性差异,表明通过不同联想途径获得的记忆,一旦稳定形成,表现在行动力上的效果是一致的,并没有显着差异。
Crook等对一种深水食腐动物鹦鹉螺Nautilus pompi-lius的空间认知策略进行了研究,结果表明,这种只有简单脑结构的水生动物虽然被认为缺少与学习和记忆相关的神经结构,但是它们依然能够利用或远或近的视觉线索对目标物进行定位,却不会使用自我中心策略,这可能表示相对利用自身方向感,利用视觉线索涉及到的神经反应过程更简单,动物更易于利用视觉线索获得空间信息,因此其对空间认知策略的选择可能取决于环境中是否有可见线索的存在,如果环境中存在可见标记物,个体会优先采用视觉提示策略。本文对鳑鲏鱼类空间认知策略的研究结果表明,在有一个标记物(视觉提示)存在时高体鳑鲏学习能力较强,和Crook的研究结果是一致的。