
�������������쳣�ۺ�������������������������������������壬�ͣģӣ���һ����Դ����Ѫ��ϸ���쳣��¡��ѪҺϵͳ��������Ҫ����Ϊ����Ѫһϵ���ϵѪϸ�����٣�������ɼ������Ե�����ǰ��ϸ���쳣��λ�����ṹ���ģΣ�����ת��ø���Ƽ����ģΣ��ͣ�������������������ɣ��������ģΣͣԣɣ��ij���Ϊ�ͣģӻ����ṩ���µ����Ʋ��ԡ�
�����ģΣ�����ת��ø ���ᣨ�ģΣ��ͣ������������������壱�ᣬ�ģΣͣԣ��ᣩ��һ��ά�� �ģΣ�����״̬��ø���ģΣ��ڸ���ʱ �ģΣͣԣ�����Լ��������ºϳɵ����� �ģΣ���ʹ �ģΣ�ά���ױ��ļ���״̬�������������ģ���������壬�ģ��ã���Ϊһ�� �ģΣͣԣɿ�ͨ������ �ģΣͣԣ��ᣬ�� �� �� �� �� �ģΣ���′���� ˮ ƽ��
�����ӣ����ȱ����� �ģΣͣԣ������ͨ�� �ζ˵�һ���ź������ж�λ���������У��������� �ģΣ���ϡ������� �ģΣ�����ĵ��ײ��������������������ĵ��Ӵ��ݹ��̣��������������������幦�ܵ��쳣���Ե��������幦�ܵĸı䡣Ŀǰ�������йأģ����Ƿ����ͨ��Ӱ���������������뵰�ı���Ӷ��ı�ϸ���Ĺ�����״̬���о����١����о���Ŀ������̽�� �ģ����Ƿ���ڳ���������������Ӱ�������幦�ܵ����á�
�������������뷽��
����������������
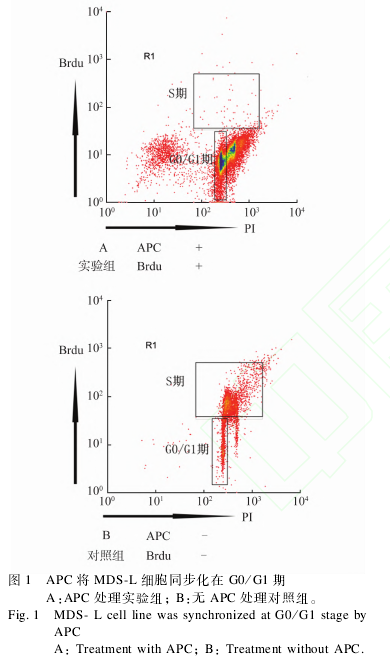
����������������ϸ��ϵ���ͣģӪ���ϸ���������Ϻ���ͨ��ѧ��������ҽԺ�������ڡ�
������������������Ҫ�Լ���������ϸ�������� ����������������̥ţѪ��ֱ������� �ǣ����� �ȣ������幫˾��
�����Уɡ��£�������Դ �����骲�£������塢�ÿ���������ƣɣԣá����ǵ�ù�ع������� �ӣ����᪲��������蹫˾��ϸ���ܣңΣ���ȡ�� �ԣ�����칺������ �̣���������������幫˾����ת¼�Լ��� ��ţ�����ģΣ�������������Ϊ�ձ� �ԣ����ṫ˾������ңԪ��Уã����� �ӣ٣£� �ǣ������ձ��ԣϣ٣ϣ£Ϲ�˾��ʵʱ���� �Уã��� �̣����������壴����Ϊ��ʿ �ң���幫˾�����ңϣӼ���Լ��й��ڱ����������Ƽ�����˾��
����������������
����������������ϸ��������ϸ������ͬ�������ͣģӪ���ϸ��ϵ�� �����桢�����ãϣ������������ں��У������գ�������ù�ء�������μ�磯������ù�ء�������̥ţѪ�� ���������������У���������������ӣ��ΰ��ت������ɣ̪����������ã������ã�����������ÿ���������䴫�����Ρ����У���Ũ�ȣ����磯�����ܽ��� �ģͣӣ��в������� �������泬���±��䡣���ͣģӪ���ϸ���� ��×�������������̽����� �����װ��У�ʵ���� �ͣģӪ���ϸ������ ���У���Ũ��Ϊ����μ�磯���̣������� �ͣģӪ���ϸ������ �ģͣӣϣ������桢�����ãϣ����������� ������ͬ���� �ͣģӪ���ϸ����
�������������������� �£�������������ϸ������ͬ����ʵ���飨���У�ͬ������ϸ�������Զ����飨�ģͣӣϣ�ϸ����������Ũ��Ϊ����μ�磯���̵� �£������廯��������ण���ϸ���������з������裬���������£������Ҵ��̶����裬���̶����ϸ������ ������죯�̣ȣã̴������������ӿ� �£����������·��� ���衣�� �ƣɣԣñ�ǵ��ÿ���������·��� �������������ں��� �Уɵ� �У£��У���ʽϸ���� �ƣ̣����ƣ̣�ͨ����� �£�������������
�����������������ģãƣȪ��ģ���� �ңϣӵIJ������� ���У�ͬ������� �ͣģӪ���ϸ���ֱ��벻ͬŨ�ȵ� �ģ������ã����裬ʵ���� �ģ���Ũ��Ϊ ����������������������μ����죯�̣�������Ϊ�� �ģ��õ� �У£ӣ��ңУͣɣ�������ϴ��ϸ�� ���Σ���ϸ�������� ����������죯�̣ģãƣȪ��ģ���̥ţѪ�壱�������У�ϸ���������з��� ���������� ����Ԥ��ģȣ���’��Һ��ϴ ���Σ������ ������μ�̣ȣ���’��Һ������������ʽϸ���� �ƣ̣���⡣
������������������ңԪ��Уãҷ���������� �ģΣ����������Σģ����Σģ����ңΣ�ת¼�仯����ͬ������ �ͣģӪ���ϸ���벻ͬŨ�� �ģ��ã���������������������������μ����죯�̣����� �����裬ʵ���� �ģ���Ũ��Ϊ����������������������μ����죯�̣�������Ϊ�� �ģ��õ� �У£ӡ����� �ԣ�����취��ȡϸ���� �ңΣ���ʹ�� ��ţ�����ģΣ������������ԣ����ᣩ�Լ��н��� ���ңΣ���ת�ʳ� ��ģΣ����� β���������Ϊ�ڲ�����Զ��� �Уãҷ������ �Σģ����Σģ�������
�������� �ԣ������ģΣ���ȡ�Լ�����ȡ ��×������ϸ��ȫ������ �ģΣ������ �����ģΣ�����Կ������仯���ȣ£�Ϊ�ڲλ�������Ϊ ��ңԪ��Уã�������������У��Σģ������Σ��ԣǣãǣ��ǣã��ǣԣ��ǣããã������ã����Ԫ��ãԣ��� �Σ��ԣԣ��ԣǣǣãã����ǣǣǣԣã��ԣǣ��ԣǣǣã����Σģ������Σ��ԣǣԣǣǣԣãǣǣǣԣǣԣǣԣԣ��ԣԣ��ԣԣã����Σ��ǣ��ã����ãã��ԣã��ԣԣãããããԣ������ԣ�β������������Σ��ǣ��ԣǣǣãã��ãǣǣãԣǣãԣԣ��� �Σ����ǣǣ��ãԣê��ã��ԣǣããã��ǣǣ������ͣ��ģΣ������Σ��ã��ããã����ǣ������ã��ǣǣǣԣԣԣǣԣ��� �Σ��ԣǣǣãã��ԣǣǣǣԣ��ԣǣԣԪ��ǣԣԣ����ȣ£ǣ��� �Σ��ǣãԣԣãԣǣ��ã��ã����ãԣǣԣǣԪ��ԣã��ãԣ��ǣã����Σ��ã��ãã����ãԣԣã��ԣãã��ãǣԣԣê����ãã��ܷ�Ӧ��ϵΪ ����μ�̣��Уãҷ�Ӧ����Ϊ�������棳�������� ������ͬ�����˻��¶��趨���£��Σģ����Σģ���β��������������� ���ͣ��ģΣ����ȣ£ǣ������棬���������� ������ ������ѭ����������������Ƭ�γ���Ϊ���Σģ�����������𣬣Σģ����������β�����������������𣬣ͣ��ģΣ�����������𣬣ȣ£ǣ���������𡣸��ݸ����������� �ã�ֵ������ ���������ã���������Ի����������
������������ͳ��ѧ�������������ݲ��� �ǣ�������У����������ͳ�����������������������Ƚϲ��õ����ط��������ʵ�����������Ƚϲ��� �ģ���������������飬���������� ����±���ʾ���У���������Ϊ������ͳ��ѧ���塣
���������ᡡ��
���������������У�ͬ���� �ͣģӪ���ϸ����⡡���Уý�ϸ��ͬ������ �ǣ����ǣ��ڣ�ϸ�������� ���ڡ���ͼ ���ɼ���ʵ���龭 ���Уô�����ϸ������ ���ڼ�����ϸ�����ڣ���δ�� ���Уô����Ķ����� �ͣģӪ���ϸ������������ϸ�����ڽ��̣����ڿɼ��д���ϸ�����ڡ�
����
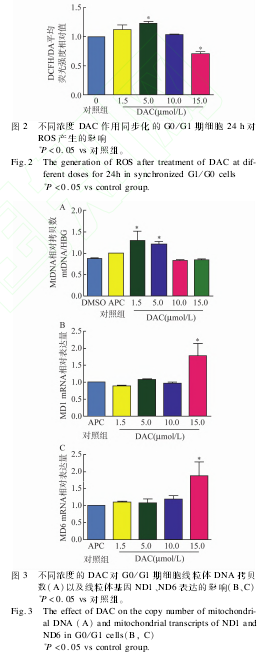
��������������ͬŨ�� �ģ��ö�ͬ���� �ǣ����ǣ���ϸ�� �ңϣӲ�����Ӱ�졡��ͼ ������ͼ ���ɼ���ʵ���� �ͣģӪ���ϸ���ڣ�μ����죯�̣ģ������ã��������Դٽ�ͬ������ �ͣģӪ���ϸ������ �ңϣӣ����������Ȳ�����ͳ��ѧ���壨�У��������������� �ģ���Ũ��Ϊ ����μ����죯��ʱϸ���� �ңϣӲ����϶��������Խ��٣��У�������������
��������������ͬŨ�� �ģ��ö�ͬ���� �ͣģӪ���ϸ��������ģΣ��������������������� �Σģ����Σģ�ת¼��Ӱ�졡���������ȣ�ͬ���� �ͣģӪ����ڵ�Ũ�ȣģ��ã�����������μ����죯�̣������¿ɴٽ������� �ģ�A���ͣ������������ģΣ����ͣ��ģΣ������� �� �� �� ���У������������������� �ģ���Ũ�����ӣ��ģ��ö� �����ģΣ�������Ӱ�첻��ͼ�������ڸ�Ũ�� �ģ��ã�����μ����죯�̣������£�ͬ���� �ͣģӪ���ϸ�� �Σģ����Σģ�����ת¼�������ӣ�����Ũ�� �ģ��ö� �Σģ����Σģ���ת¼Ӱ�첻��ͼ ���¡��á�
����
���������֡���
�������о��������Ŵ��쳣������������չ�в�����Ҫ�����á��ģΣ�ȥ����ҩ��ij��ָ� �ͣģ������ṩ���µIJ��ԡ������������ģ��ã���Ϊ�����ƣģ��������������ٴ��� �ģΣ�ȥ����ҩ��֮һ���ֳ��Ϻõ�Ӧ��ǰ�������� �ͣģӻ��ߴ������档Ŀǰ���о���Ϊ���ģ��÷��ӿ��������þ���Ũ�������ԣ��ڵ�Ũ���� �ģ�����Ҫ���� �ģΣ�ȥ�������ã�ʹ����������Ĭ���ְ��������±������Ũ�� �ģ�����ͨ�������� �ģΣ����Ϸ���ϸ�������á�
������������һ�ִ����ڰ����ڵ�ϸ�����������������ѧ������������л�����ɻ�������������ƽ�⡢������ת¼�����Ρ�ϸ�������ȡ�������ģΣ�������������ȱʧ�Լ��������������ͻ��ɼ��ڶ���������֯�У����������ķ�����չ��Ԥ���йء��ͣģ���һ�ֶ�����껼�ߵ�ѪҺϵͳ������ͨ���� �ͣģӻ��ߵ������� �ģΣ������о����������� �ͣģӻ��������� �ģΣ�ͻ��Ƶ�����ӣ���ͻ�����ڷDZ���������������ģΣ����뵰�ı������Լ������� �ģΣ��Ŀ����������쳣Ҳ�ɼ��� �ͣģӻ��ߡ�
������ʵ��ͨ�������Ԥ��ʹ �ͣģӪ���ϸ��ͬ������ �ǣ����ǣ��ڣ����� �ģ��õ� �ģΣ��������ã��Դ�̽�� �ģ����Ƿ��ж�ϸ�����ӷǻ������������������á����У���һ�� �ģΣ��ۺ�ø���ƣ����������Խ�� �ģΣ��ۺ�ø������ �ģΣ����ƺϳɴӶ���ϸ������ͬ������ �ǣ����ǣ��ڡ���ʵ������ʾ�����Уÿ��Ժ���Ч�ؽ�ϸ��ͬ������ �ǣ����ǣ��ڡ���ͬ������ �ͣģӪ���ϸ�����費ͬŨ�� �ģ������ú��֣��ڵ�Ũ�ȣ�����������μ����죯�̣��� �ģ��ÿɴٽ�ͬ����ϸ���ңϣӲ�����������Ũ�ȵ����ӣ�ͬ����ϸ�������ģңϣ�ȴ���ڶ����顣�������������������Ӵ�����ϸ���� �ңϣ���Ҫ��Դ���Ʋ������ �ģ��øı������幦���йء�ͨ����ⲻͬŨ�� �ģ��ö�ͬ���� �ǣ����ǣ�ϸ�������� �ģΣ��������Լ���������� �Σģ����Σģ����Σ��ģȣ�������������壱�����������Ӱ�졣��ʵ������ʾ����Ũ�� �ģ��ã�����������μ����죯�̣����Դٽ������� �ģΣ����������ӣ�����Ũ�� �ģ���ȴ�����á������� �ģΣ����ƾ��а������ԣ����� �ģΣ��ۺ�ø γ���ǻ�������������������ģΣ��������������γ�������������������������������������أ�����������ļ����̶��������� �ģΣ��������ʸ���ء����⣬���о����֣�����άԭϸ�����ܵ���Դ�Ի���Դ���������ʴ̼����ɴ̼�������ģΣ����������ӡ���ʵ������ʾ���� �ģ���Ũ��Ϊ ��μ����죯��ʱ�������̶ȴٽ�ϸ���� �ңϣӲ���������Ũ�ȶ������� �ģΣ�������Ӱ�첢�������ԡ��ɴ˿ɼ�����Ũ�� �ģ�������������� �ģΣ����������ӿ����� �ģ���ȥ���������йء�����Ũ�����ӣ��ģ������������� �ģΣ����ˣ�ʹ������ģΣ����������٣������� �ģ���ȥ�����������µ������� �ģΣ����������ӵ����á���ˣ��ڸ�Ũ�� �ģ������ã�ͬ���� �ͣģӪ���ϸ�������� �ģΣ�������δ�����Ըı䡣�����о����������Уö������幦����Ӱ�죬����ʵ������ʾ�����Уô�������ģͣӣ϶�������ȣ������� �ģΣ������������Եı仯���Σģ����Σģ����ѱ��������������� �ģΣ�����ת��ø������ص����������Ũ�� �ģ��ö�������� �Σģ����Σģ�����������Ӱ�죬���֮�£���Ũ�� �ģ��ÿ��Դٽ����ߵı����������������������������װ���쳣��Ӱ���������������Ӵ��ݣ����Ը�Ũ�� �ģ���ͨ���ı���������� �Σģ����Σģ����ױ����������������װ���쳣���Ӷ�Ӱ�������������������Ӵ��ݣ��������ȣ����ͣңϣӵIJ�����
�����������������о�����̽���� �ģ��ô���ϸ����������������ã���ͬŨ�� �ģ��ÿ���Ӱ�������� �ңϣӵIJ������ı������� �ģΣ�����������������������������ã�Ϊ�����о� �ģ��õ����û����ṩ�������ݡ�
�����ο����ף�
�����ۣ��ݣΣ�����ӣģ��ͣģӣ����������������������������������������������������������������������������������������������������������ۣʣݣ��ȣ��������������� �ӣ��ȣ��������ţ����У���������������������������������飺������������������������������������������������
�����ۣ��ݣϣ�����ˣ��ӣ�����ã��ԣ��������ͣ������죮�ɣ�����������������������������������������������ãģ�����������������������������������������������������壨�ãģ������ãģ������ãģ������̣����������������������������������������������������ۣʣݣ��ӣ��������죬��������������������������������������
�����ۣ��ݣУ�����ˣ��ģ������ʣ��ģ��ӣ������죮�ԣ����������棵������᪲��′������������������������������������������������������ģΣͣԣ������������������������������������������������������ۣʣݣ��Σ�������������ң��������������������������������������������
�����ۣ��ݣ٣���������٣��ӣ����ó�У��ӣ�ü����ģ������죮�ң�����������������ģΣ�������������������������������ۣʣݣ��ţ���������������������������������������������
�����ۣ��ݣӣ����̣ӣ��ԣ������У֣��У��������ţʣ������죮�ģΣ����������������������壱�������������������������������������������������������������������� ������������� ���������������ۣʣݣ��УΣ��ӣ���������������������������������������������
�����ۣ��ݣ��������֣����֣��ԣ��������̣��ԣ������������������������������������������������������������������������������ۣʣݣ��Σ������������ɣ������������������������������������������
�����ۣ��ݣ̣���������������֣��ã�����������������������������������骲�����������������ۣʣݣ��ã���ţ�����������������������������������������������
�����ۣ��ݣƣ����ʣ��ʣ���������������������У£��ģΣ����������������������������������������������������������������������������者����������������������������������������ۣʣݣ��ţ�������ϣ���Ԫ�����У������������������������������������������������
�����ۣ��ݣӣ�����������ã��£���������£��ͣ�����ԣ������죮�ƣ�������������������������ģΣ� ���������������������������������������������������������ۣʣݣ��ã�����ң������������������������������������������
�����ۣ����ݣ٣��ͣ��ǣ���������������������������������������������骲�������������ģΣ����������������������������������������㪲����ۣʣݣ��̣���ӣ�飬��������������������������������������
�����ۣ����ݣʣ���ͣ��ˣ���ӣӣ��̣��ʣ��ã������������������������������������������������������������������ۣʣݣ��ţ���ͣ��ͣ�䣬��������������������
�����ۣ����ݣף��������ͣ���ü�������ã��ԣ�������ã������죮����������������������������ģΣ������������������������������������������������������ۣʣݣ��ţ���ȣ�������죬��������������������������������������
�����ۣ����ݽ��죬�����٣����������ȣ����������쳣�ۺ������������� �ģΣ��ģߣ������ͻ���о��ۣʣݣ�ɽ����ѧѧ����ҽѧ�棬��������������������������������������
�����ۣ����ݣɣ��������ӣ�����ӣ��У������ã������죮�ģ������������������쪲������������������������������� ��������������������������������������Σģգƣӣ�����Σģգƣӣ������������������ɣۣʣݣ��ʣ£骲���ã�������������������������������������������������������
�����ۣ����ݣ̣���У��ģ������£��ģΣ������������������������������������ͣ�����������������������������ۣʣݣ��ţ�������ͣ��ͣ���������������������������������������������
�����ۣ����ݣˣ�����ңģ��ͣ�����������ͣ�ˣ������ͣ������죮�ͣ�������������ģΣ���������������������������������������������������������ģΣ� ������������������������������������ģΣ�����������������������ۣʣݣ��Σ�������������ң������������������������������������������������
�����ۣ����ݣ̣��ȣã��٣��Уȣ��̣��ã٣������죮�ɣ��������������者���������������������������ģΣ������������������������������������������������ۣʣݣ��£�������ʣ�����������������������������������������
�����ۣ����ݣã��������骲�£���¨���ӣ��֣�����������ȣ��ã��������ţ������죮�ģ����������������������������������������������������������������������������������������������������ۣʣݣ��ţ���ã���ң����������������������������������������
�����ۣ����ݣģ����ƣ��ţ��í�������ʣ����ͣ�����ãԣ��ã�����������ң������������������������������������������������������������骲����������������������������������������ɣ���ɣ֣ۣʣݣ��ͣ��ã���£��죬������������������������������������.
