本文主要综述OPG/RANK/RANKL的特点、作用机制,及其在骨代谢疾病、药物和相关营养因素干预效果评价等领域的应用现状,并展望将其引入实验动物骨骼发育营养评估领域的应用前景。大家在相关论文写作时,可以参考这篇题目为“骨代谢营养学实验中OPG/RANK/RANKL系统综述”的人体生理学论文。
原标题:OPG / RANK / RANKL系统在实验动物骨骼发育营养需要评估中的应用
摘要:OPG/RANK/RANKL系统是调节骨代谢的主要信号通路,在维持骨形成和吸收之间的动态平衡中发挥着重要作用。本文主要综述OPG/RANK/RANKL的特点、作用机制,及其在骨代谢疾病、药物和相关营养因素干预效果评价等领域的应用现状,并展望将其引入实验动物骨骼发育营养评估领域的应用前景。
关键词:OPG;RANK;RANKL; 实验动物;骨骼发育;营养需要
骨骼发育需要多种营养成分参与,这些营养成分( 如钙、磷等) 不仅是骨骼的重要组成成分,而且对维持正常骨代谢具有重要意义[1].骨形成和吸收之间的动态平衡是保持正常骨代谢必要条件,而这种平衡需要依靠相应的细胞通路来调节。OPG/RANK / RANKL系统是近年发现的调节骨代谢的重要通路[2 - 3],由肿瘤坏死因子家族的三个新成员-①护骨素(osteoprotegerin,OPG)、②核因子κB受体活化子(receptor activator of NF-κB,RANK) 和③核因子κB受体活化子配体(receptor activator of NF-κBligand,RANKL) 组 成。目 前,有 关OPG / RANK /RANKL系统在骨代谢疾病病因学及药物疗效评价研究等领域报道很多,而在营养干预效果评价方面则报 道 较 少。因 此,本 文 在 阐 述OPG/RANK/RANKL系统的特点和作用机制的基础上,对近年来该系统在骨代谢疾病及相关药物和营养因素疗效评价的研究进展作一综述,为将其广泛应用于与骨代谢相关的营养学研究提供理论依据。
1 OPG / RANK / RANKL系统概述
1. 1 OPG,RANK和RANKL的结构与功能
1. 1. 1骨保护素 (OPG)
OPG是由Simonet等[4]于1997年首先在大鼠上发现的一种新型肿瘤坏死因子,属于TNF受体(tumor necrosis factor receptor,TNFR) 超家族成员。大鼠、人、和小鼠OPG同源性大于85%[4].OPGmRNA可在小鼠皮肤、肝脏、肺和心脏等组织中表达[4].OPG蛋白属分泌型糖蛋白,无疏水跨膜区,氨基酸全长为401个残基,包含21个氨基酸的信号肽和380个氨基酸成熟肽[5],共7个结构域(D1 ~D7) :D1 ~ D4区为半胱氨酸含量丰富区域,位于N端,主要功能是抑制破骨细胞分化和骨吸收;D5、D6区是位于C端的高度同源的两个区域,为死亡域。D7区含有肝素结合位点,可通过N端糖基化,形成巯基化二聚体[4].
OPG主要功能是作为诱饵受体调节配体和受体之间的相互作用。如OPG与RANKL具有高度结合能力,是RANKL的诱饵样受体,对RANK信号起着负调控的作用。体外实验表明OPG可抑制破骨细胞(osteoclast,OC) 产生[4].
1. 1. 2 NF-κB受体活化因子(RANK)
RANK也属于TNF受体家族成员,是RANKL的信号受体,属I型跨膜蛋白。人的RANK蛋白是由616个氨基酸的组成的同源三聚体,其胞外结构由208个氨基酸组成,包含4个富含半胱氨酸的重复序列[6],为N-末端,主要功能是与RANKL的C-端结合并产生传递信号。RANK在细胞内的信号传递需要由细胞浆内的肿瘤坏死因子受体相关因子(TRAFs) 介导[7].RANK蛋白的胞内结构域有383个氨基酸,在细胞膜末端的氨基酸胞质尾区可与TRAF家族中的1、2、3、5因子结合,使其活性被激活; 在 细 胞 膜 近 端Pro-X-Glu-X-X-(aromatic /acidresidue) 结合部与TRAF6结合[8].RANK mRNA广泛存在于胸腺、肝脏、骨胳肌、结肠、胰腺、肾上腺、心脏、乳腺、骨髓、肺、脑、肾脏、皮肤等组织器官[9].
RANK信号区别于其他TNFR超家族成员的一个重要特征是它不仅可激活典型的NF-κB信号通路,还 可 以 激 活 非 典 型 的NF-κB信 号 通 路[10].RANK的主要功能是与RANKL结 合形 成RANK-RANKL信号,在相关信号转导因子的参与下,促进OC的形成和骨吸收功能的加强[11].
1. 1. 3 RANK的配体 (RANKL)
RANKL是由4个独立的研究团队分别发现并加以命名的肿瘤坏死因子超家族成员[12],因此还被称为肿瘤坏死因子活化诱导细胞因子(TRANCE) ,破骨细胞分化因子(ODF) ,护骨素配体(OPGL)。
RANKL蛋白属II型跨膜蛋白,与肿瘤坏死因子超家族(TNFSF) 成员中的TRAIL,Fas L和TNF-a具有 同 源 性[13].人 和 小 鼠 的RANKL蛋 白 具 有83%的同源性[11],小鼠的RANKL蛋白包含316个氨基酸,由细胞外C-端受体作用区域和跨膜区域组成。研究发现RANKL蛋白存在膜结合和分泌型两种形式[9].RANKL蛋白有3种不同的亚型,最短的亚型缺乏细胞内区域和跨膜区,以分泌型存在于细胞外基质中,对RANKL活性具有抑制作用[14].
RANKL mRNA可在多种组织表达,包括T淋巴细胞,成骨细胞,骨细胞,骨基质和肺[12].RANKL不仅可与功能性受体-RANK结合,还可以与诱饵受体-OPG结合[12].RANKL对骨代谢具有重要调节作用,它与RANK结合后产生的转录活化信号,可促进OC的分化成熟[13].
1. 2 OPG / RANK / RANKL系统对骨代谢的调节机制
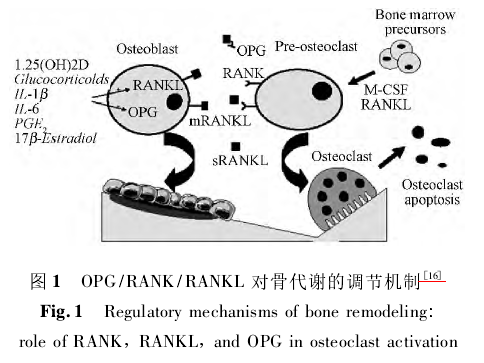
OPG / RANK / RANKL系统在维持骨吸收和骨形成两者动态平衡,防止骨的丢失,保证正常的骨转化更新等方面起着关键调节作用[15],并主导了破骨细胞(OC) 的形成和活化的调控[16].其作用机制是[16]: 破骨细胞(OC) 和成骨细胞(osteoblast,OB)分别介导骨的吸收和合成过程,RANKL主要由成骨细胞前体细胞及骨髓基质细胞表达,RANK主要由破骨细胞表达,RANKL与RANK结合,可促进破骨细胞分化、融合及成熟,并抑制破骨细胞凋亡,从而促进骨的吸收; 与此同时,成骨细胞也表达OPG,OPG实质上是一种受体诱饵,可与RANKL家族竞争性结合,抑制破骨细胞的形成、成熟。RANK与OPG竞争性地与RANKL的结合,使骨吸收和骨形成处于平衡状态( 图1)。因此,正常情况下,骨组织中RANKL、RANK与OPG三者相互间比值保持恒定,一旦由于某种原因使这种稳态被打破,就可能引起骨代谢紊乱,导致相应骨代谢疾病的发生。如OPG缺失小鼠具有骨质疏松的骨病变[17],其病因就是OPG缺失使体内OPG、RANK、RANKL相互间的正常比例被打破,RANKL与RANK结合的能力增强,从而促进了OC的增殖。