��������ѧ����

����ժ Ҫ����Ŀ��:��ͻ�������Ժ���ͻ����̽�����������˴���ģ��ʹ�������Ļ��ơ�����:��������������ѹ�������� (chronic compression injury, CCI) �յ���������������ʹ, ���CCI������е��ʹ���������䴥�շ�ʹ�Լ���ʹ��������ָ�ꡣʵʱ����PCR���CCI��14 d��21 d�Ĵ���ͬ��L4-6��������GAP-43��NCAM��PSD-95��Syp��kinesin��dynein�ı�����:CCI����14 d����Ļ�е��ʹ���������䴥�շ�ʹ�Լ���ʹ������ָ������� (P��<0. 01) ��CCI������14 d GAP-43��NCAM�ı������� (P��<0. 01) , PSD-95��Syp��kinesin��dynein�ı�������� (P <0. 05) ������21 d GAP-43��NCAM�ı�����½� (P <0. 01, P <0. 05) , PSD-95��Syp��kinesin��dynein�ı�������� (P <0. 05) ������:����������14 d��, ��Ҫ������Ԫ�ṹΪ����������14 d��, �˷ܵĴ����Լ���ͻ����������
�����ؼ��ʣ���ʹ������; ͻ��������; ��ͻ����; ʵʱ����PCR; ����������;
����Abstract����Objective: To explore the mechanism of hyperalgesia with sciatic nerve injury rats model in terms of synaptic plasticity and axonal transport. Methods: Peripheral neuropathic pain in rats were induced by chronic compression injury ( CCI) of sciatic nerve. The indexes of mechanical hyperalgesia, cold allodynia and thermal hyperalgesia in rats after CCI were detected. The expressions of GAP-43, NCAM, PSD-95, Syp, kinesin and dynein in the L4-6 dorsal root ganglion at 14 d and 21 d after CCI were detected by realtime quantitative PCR detection. Results: The indexes of mechanical hyperalgesia, cold allodynia and thermal hyperalgesia in rats in 14 d after CCI were reduced ( P < 0. 01) . The expressions of GAP-43 and NCAM in 14 d after CCI were increased ( P < 0. 01) , and the expressions of PSD-95, Syp, kinesin and dynein were reduced ( P < 0. 05) . The expressions of GAP-43 and NCAM in 21 d after CCI were reduced ( P < 0. 01 or P < 0. 05) , and the expressions of PSD-95, Syp, kinesin and dynein were increased ( P < 0. 05) . Conclusion: The repair of the neuron structure is predominant in the 14 d after sciatic nerve injury. After 14 d of injury, transmission of excitement and axon transport are initiated.
����Keyword����Hyperalgesia; Synaptic plasticity; Axonal transport; Real-time quantitative PCR; Sciatic nerve injury;
����ʹ����������������ʹ����ʹ����, ָ�������̼�������������[1]����ʹ�ж����ұ�����Σ��ʶ��Ϳ����˺����ӳ���������Ҫ��Ȼ��, ��֯���ˡ�������֢������ѧ�����յ��˺�����Ԫ������, �Ӷ����²�������ʹ������, ��ʹ��Ϊ��������, ʧȥ��ά�����������Եı�������[2]�������������µ�����ѹ���������ѱ��㷺�����о�����ѧ��ʹ�IJ��������, ���о���������������ѹ�������� (chronic compression injury, CCI) ��Ϊʵ�鶯��������ʹ�յ�ģ��[3], ���CCI�������Ļ�е��ʹ���������䴥�շ�ʹ�Լ���ʹ��������ʱ��ı仯��Ŀǰ, �о��Ѿ�ȷ���˴���������ʹ������ϸ���ͷ����ź�, ����������ʹ�������������ںܴ�̶��ϻ���δ֪�ġ��о�[4]����, ͻ��ǿ�Ⱥ�Ч������������, ���յ�������ʹģ�Ͷ�������Ի�е�Դ��շ�ʹ������, ��ͻ�����ϰ����ܵ�����ͻ��е�����Ժͻ�е������Ӧ[5], �˻���Ϊ��ʹ���о��ṩ���µ�˼·�����о���Ҫ��ͻ���������Լ���ͻ����������̽��CCI����ʹ�������Ļ���, ̽��CCI����������ص��� (GAP-43) ������άճ������ (NCAM1) ��ͻ�������ܵ��� (PSD-95) ��ͻ������ (Syp) ���������� (kinesin) �Լ��������� (dynein) ��Ӱ�졣
����1 ���Ϻͷ���
����1.1 һ������
����ѡ��180~221 g��������Sprague-Dawley (SD) ����40ֻ, ����������ͨ��ѧҽѧԺʵ�鶯�����ġ�ʵ�鶯����241�������������ιʳ����ˮ, 12/12����/�ڰ�ѭ���������������Ϊ����:Sham�� (��������, n=20) , CCI�� (������������, n=20) ��
����1.2 ��������������ģ��
��������CCI�յ���������������ʹ[3]�������ض���������Ĥ��ע��40 mg/kg����ͱ����ƽ����������ڹɼ����·���3~4 mm������, ��¶������, ������4-0������4������λ���������, ���Ϊ1 mm����������:ֻ��¶��������������֮��, ���������˿�߷�ϼ����Ƥ����, ��Ӧ�þֲ������ء�����������������������½��С�
����1.3 ��ʹ������Ӧ���
�����ֱ�����ǰ1 d������1 d��3 d��7 d��14 d��21 d�Դ��������ʹ������Ӧ���ԡ�
����1.3.1 ��е��ʹ������
��������von Freyϸ˿������е��ʹ������[6]�����������ܽ�����������, ͨ������von Frey˿��������������֫��ױ���ʩ��ѹ������¼�������������֫����ʱ��ѹ��ֵ, ��Ϊ���㷴����ֵ (PWT, g) ��ÿ�����3��, ���15 min, ȡ3��ƽ��ֵ��
����1.3.2 �䴥�շ�ʹ
�������ñ�ͪ�����������䴥�շ�ʹ[7]�����������ܽ�����������, ������צ��ױ�������100μL��ͪ����¼�����������ʱ�� (c-PWL, sec) ��ÿ�����3��, ���15 min, ȡ3�ε�ƽ��ֵ��
����1.3.3 ��ʹ������
��������Eddy’s�Ȱ�������ʹ������[8]�������¶��� (52.51.0) ��, ��¼���غ�צ����צ��ʱ��, ��Ϊ��צ�����㷴����ֵ (hPWL, sec) ��Ϊ�����צ����, ����15 sΪ��ֹʱ�䡣ÿ�����3��, ���15 min, ȡ3�ε�ƽ��ֵ��
����1.4 ʵʱ�����ۺ�ø��ʽ��Ӧ (qRT-PCR) ����
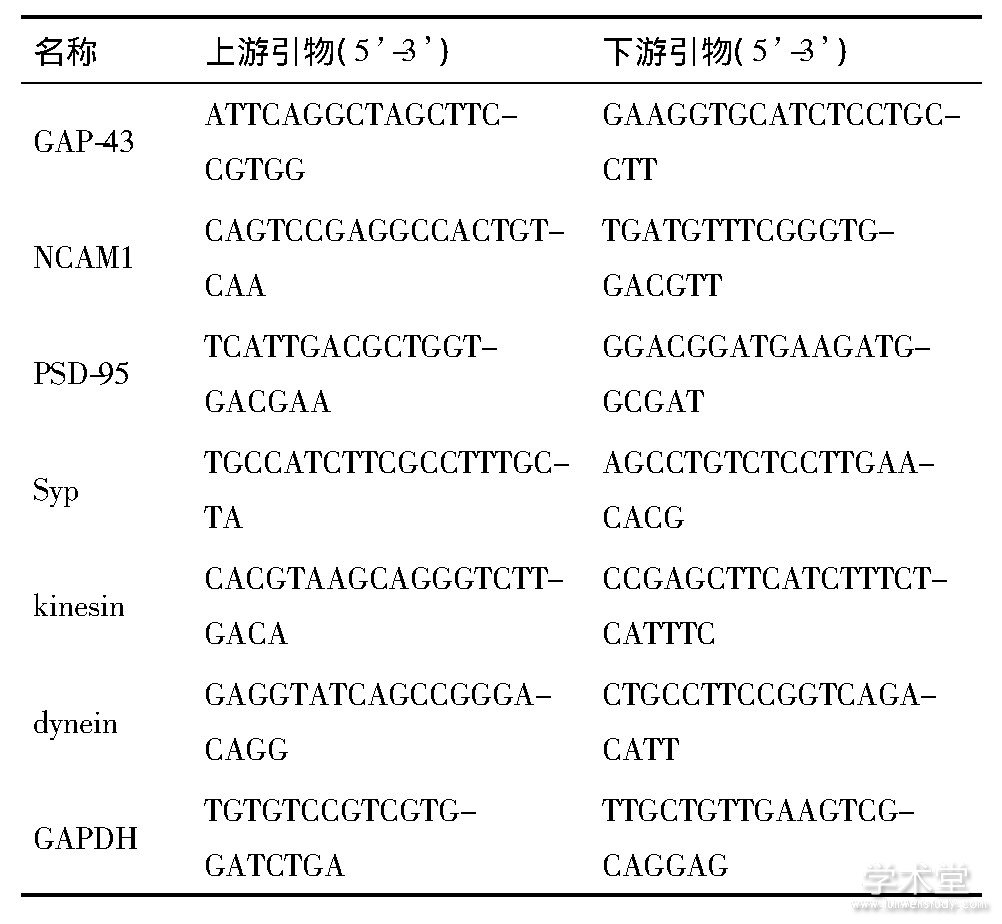
����CCI��14 d, �ֱ�ȡ�����10ֻ����ͬ��L4-6������ (DRG) ����qRT-PCR������ʣ��������������Ϊѧ���, CCI��21 d, ȡ�����ʣ���10ֻ����DRG����qRT-PCR��������Trizol�Լ� (Invitrogen, USA) ��DRG�з�����RNA��2μg��RNA����High-Capacity RNA-to-cDNA�Լ��� (Applied Biosystems, USA) �ϳ�cDNA������˵����, ʹ�����������SYBR Green I�Լ���RT-PCR�Լ���, ��Bio-Rad i Q5����PCRϵͳ (Takara, �й�) �ϼ��mRNAת¼���ˮƽ����Ա���ˮƽͨ��2-DΔCt�������㡣�����ڲλ���GAPDH�����л�����Primer-BLAST����������������������, ������Ƽ���1��
������1 �������
����1.5 ͳ��ѧ����
��������SPSS 19.0�����Աȷ���, ���������� (��±s) ��ʾ��Sham���CCI��PWT��c-PWL��hPWL�ڸ�ʱ���ĶԱȲ���t���顣ͻ��������ָ���Լ���ͻ����ָ����÷���������ж����Ƚ�, LSD���رȽϡ���P<0.05Ϊ������ͳ��ѧ���塣
����2 ���
����2.1 CCI��ʹ��������Ӱ��
����ͼ1A��ͼ1C��ʾ, ��ǰ1 d, �����PWT��h-PWL�������Ų��졣������1 d��ʼ������14 d, CCI������PWT��h-PWL����ʱ������Ӷ�����, ����ÿ��ʱ�������Sham�����, �������ͳ��ѧ���� (P<0.01) ��CCI������PWT��h-PWL������21 d��������14 dʱ���ӡ�
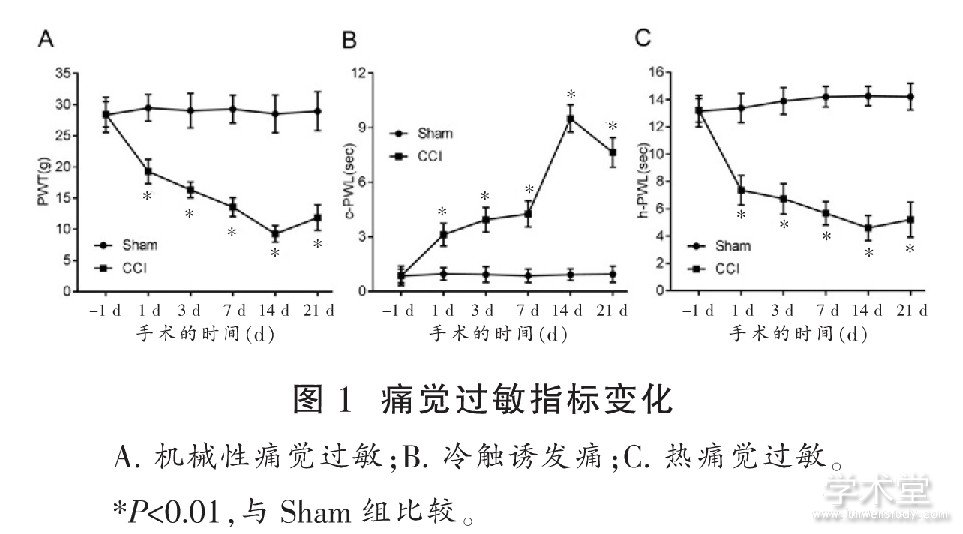
����ͼ1B��ʾ, ��ǰ1 d, �����c-PWL������ͳ��ѧ���塣������1 d��ʼ������14 d, CCI������c-PWL��ʱ������Ӷ�������, ����ÿ��ʱ�������Sham�����, �������ͳ��ѧ���� (P<0.01) ��CCI������c-PWL������21 d��������14 dʱ���͡�
����2.2 CCI��ͻ�������Ե�Ӱ��
����ͼ2A��ͼ2B��ʾ, ��Sham-14 d�����, CCI����14 d DRG��GAP-43��NCAM1�ı������� (P<0.01) ��������21 d, ��CCI-14 d���, GAP-43��NCAM1�ı�����½� (P<0.01, P<0.05) , ��δ�ָ�������ˮƽ��
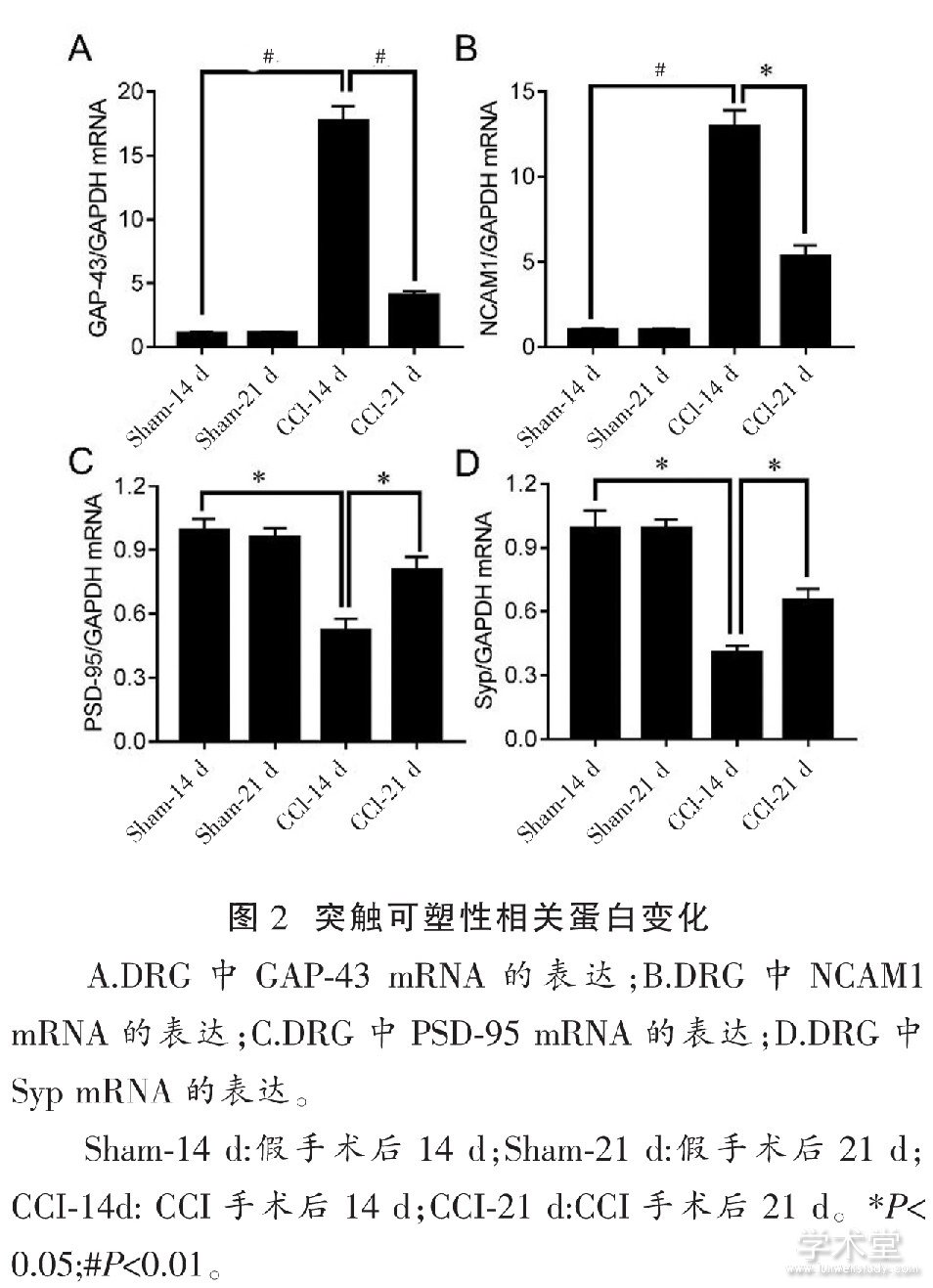
����ͼ2C��ͼ2D��ʾ, ��Sham-14 d�����, CCI����14 d DRG��PSD-95��Syp�ı�������� (P<0.05) ��������21 d, ��CCI-14 d���, PSD-95��Syp�ı�������� (P<0.05) , ����δ�ָ�������ˮƽ��
����2.3 CCI����ͻ�����Ӱ��
����ͼ3��ʾ, ��Sham-14 d�����, CCI����14 d DRG��kinesin��dynein�ı�������� (P<0.05) ��������21 d, ��CCI-14 d�Ա�, kinesin��dynein�ı������� (P<0.01) , �Ҹ�������ˮƽ��
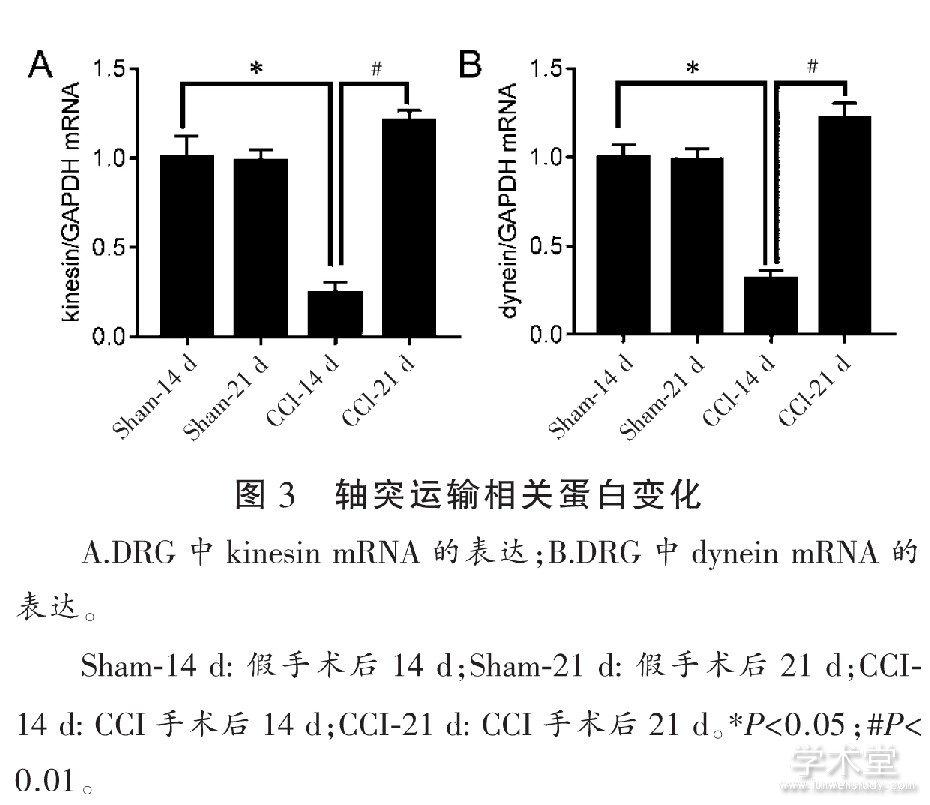
����3 ����
�������о�ּ�ڴ�ͻ�������Ժ���ͻ����������̽�ִ���������CCI������ơ����о�[3]����, CCI�յ������������˵�������̬ѧ������ѧ���Ÿı�, �������������˺�Լ2�ܹ۲쵽���ЧӦ�����, ���о�������CCI��21 d��ʹ������ָ��, ���䴥�շ�ʹ�ļ��, ������ͪ��, �����������Ӧ�ǿ����˻�צ��, Ѹ�ٷ�������˿�ϡ�����������ʹ������, ���צ���ز���������˿�ϵij���ʱ��ϳ�, ���ͨ�������������ʱ�� (cPWL, sec) ������צ�Ա�ͪ���䴥�շ�ʹ�������ʾ, ����������1 d��ʼ����PWT��h-PWL��ʱ������Ӷ�����, ������14 dʱ�����, 21 dʱ��ʼ�ָ�����������������˺����Ի�е���˺������г̶���ǿ, ���ȴ̼������ܽ��͡�����������1 d��ʼ����PWT��h-PWL��ʱ������Ӷ�����, ������14 dʱ�����, 21dʱ��ʼ�ָ���c-PWL������������1 d��ʱ������Ӷ�����, ������14 dʱ�����ֵ, 21 dʱ��ʼ���͡���������������˺���������̼��շ�����ʹ��ά��ʱ�����ӡ�
�����о�����, DRG������ʹ������״̬�����������֮һ, �������˿����յ�DRG�л���͵����ʱ���ĸı�, ����ʹ�������ķ�չ[9]�����, ���о���Ҫ��DRGΪ�о�����, ̽��CCI��14 d��21 d DRG��ͻ�������Ժ���ͻ����ָ��ı仯��
������������ص���GAP-43����Ҫ��������������ͻ��ǰĩ�ҵ�һ��ϸ����Ĥ��ص��ס�����øC�������ữGAP-43��˿����41 (Ser41) λ��, ���ữ��GAP-43ͨ���ȶ���������˿��������������ѿ�Լ�ͻ������[10]���о�[11]����, GAP-43 mRNA�ı�����������Ϻ�2���ڴﵽ��߷�ֵ, Ȼ�����͡����뱾�о����һ��, ��CCI��14 d GAP-43 mRNA�ı�����������, ��CCI��21 d���Ž���, ����CCI 2�ܺ�, �����������������ɡ�����άճ������NCAM1�ǵ���ͻ����������ͻ������ϸ��������ú�ͻ�������Եĵ�����[12]���о�����, NCAM1������Χ�������յ�����ǰ�۴�Ƥ����ض�ͻ������ת�ij�����ǿ��һ����, ��NCAM1�鵼��������, ��������Ϊ��������Χ������, ͻ�����ܶȲ��ֵ�NCAM1ˮƽ������ǿ[13]�����о���ʾ, ��CCI��14 d NCAM1 mRNA�ı�����������, ����ʱʹ������֢״����Ϊ����, ��CCI��21 d NCAM1 mRNA�������Ž��͡�
����ͻ�������Ե�����Ϊ�˷���ͻ��, ���˷���ͻ����ͻ����λ������ͻ�������ܵ��� (PSD) ������ͻ����Ĥ���γɵ�[14,15]���о�[16]����, ������Ԫ��PSD-95�Ĺ�����ɴٽ�ͻ������, �ر��ǹȰ�����ͻ���ij��졣���о���ʾ, ��CCI��14 d PSD-95 mRNA�ı������Ž���, ������������������, �����˺�14 dֻ������PSD-95������ͻ����Ĥ, �����˷ܡ���CCI��21 d PSD-95 mRNA������������, ��ʾ������������, ����PSD-95������ͻ����Ĥ, �γ��˷���ͻ����ͻ����λ��, �����˷ܡ�Syp��һ�ָƽ���ǵ���, ��Ϊ�������ʵ�ͻ��ǰС�ݵ������Ĥ������ֱ�����[17], ����Ҫ����ͻ��С���ںϺ������ͷŹ���, �ѱ��㷺��Ϊ��ͻ���ܶȵı��[18]���о�[19]����, ��������CCI��, Syp���ı����ֵ���������˺�14 d, ��Ȥ����, ���о������ʾ, CCI��14 d Syp mRNA�ı������Ž���, ��CCI��21 d��������, ����CCI 14 d��ͻ��С�ݵ��ں��Լ����ʵ��ͷŵ�����Ϣ���ݹ����ܵ�����, ��14 d��, �˹����ָ������, ����CCI��Syp������ʱ��ı仯, ���д���һ����ϸ�о���
�����Ὤ����ķ������������ﵰ���е��������� (kinesin) �Ͷ������� (dynein) ˮ��ATP������������[20]��Kinesin��Ӫ�����ʺ�dynein�Ӱ������䵽�������ͻԶ�� (˳������) , dynein������ͻԶ�˺�, ��ֱ�ӻ���Ӻ�Ӫ�����ʽ��, ��Ӫ���������������� (��������) [21]�����о������ʾ, CCI��14 d kinesin��dynein mRNA�ı������Ž���, ���������������˺�14 d��, ��ͻ���䷢���ϰ�����CCI 14 d��, ������������صĵ�����GAP-43��NCAM1������, ���������Ԫ, Ϊ��ͻ�����ͻ�������ṩ�ṹ������������ͻ��С�ݵ���Syp�Լ�ͻ����Ĥ���ܲ�λ����PSD-95��CCI��14 d�ڵĽ���, ������ʱͻ����Ĥ�Ľṹ����������, ͻ��С�ݵ��ں��Լ����ʵ��ͷŻ������ϰ�, ���²��ܴ����˷ܡ���Щ���������ڴ�ʱkinesin��dynein���ױ��オ��, û���㹻��Ӫ������, ��˲�����ά����Ԫ���ܵ��������ġ���CCI��21 d kinesin��dynein mRNA��������, ͬʱSyp��PSD-95�ı���������, ������ʱ��Ԫ����������, ����Ԫ�ṹ�����ѽӽ�β��, ���GAP-43��NCAM1�ı��オ�͡�
������֮, ���о�����CCI��14 d, ����Ļ�е��ʹ���������䴥�շ�ʹ�Լ���ʹ�������ﵽ���ЧӦ, 14 d��ʼ���⡣GAP-43��NCAM1��CCI��14 d����������, ����14 d��, ��Syp��PSD-95��kinesin��dynein��CCI��14 d�����Ž���, ����14 d�����ӡ�����ʾ����������14 d��, ��Ҫ������Ԫ�ṹΪ����������14 d��, ͻ��С�ݵ��ںϡ����ʵ��ͷš��˷ܵĴ����Լ���ͻ��������, ����ʱ��Ԫ�ṹ�����ѽӽ�β����
�����ο����ף�
����[1] Jaggi AS, Jain V, Singh N. Animal models of neuropathic pain[J].Fundam Clin Pharmacol, 2011, 25 (1) :1-28.
����[2] Colloca L, Ludman T, Bouhassira D, et al. Neuropathic pain[J].Nat Rev Dis Primers, 2017, 3:17002.
����[3] Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man[J]. Pain, 1988, 33 (1) :87-107.
����[4] Jaken RJ, Joosten EA, Knuwer M, et al. Synaptic plasticity in the substantia gelatinosa in a model of chronic neuropathic pain[J].Neurosci Lett, 2010, 469 (1) :30-33.
����[5] Dilley A, Richards N, Pulman KG, et al. Disruption of fast axonal transport in the rat induces behavioral changes consistent with neuropathic pain[J]. J Pain, 2013, 14 (11) :1437-1449.
����[6] Chaplan SR, Bach FW, Pogrel JW, et al. Quantitative assessment of tactile allodynia in the rat paw[J]. J Neurosci Methods, 1994, 53 (1) :55-63.
����[7] Choi Y, Yoon YW, Na HS, et al. Behavioral signs of ongoing pain and cold allodynia in a rat model of neuropathic pain[J]. Pain, 1994, 59 (3) :369-376.
����[8] Jain V, Jaggi AS, Singh N. Ameliorative potential of rosiglitazone in tibial and sural nerve transection-induced painful neuropathy in rats[J]. Pharmacol Res, 2009, 59 (6) :385-392.
����[9] Chen WH, Chang YT, Chen YC, et al. Spinal PKC/ERK signal pathway mediates hyperalgesia priming[J]. Pain, 2018.[Epub ahead of print].
����[10] Benowitz LI, Routtenberg A. GAP-43:an intrinsic determinant of neuronal development and plasticity[J]. Trends Neurosci, 1997, 20 (2) :84-91.
����[11] Yao GL, Kiyama H. Dexamethasone enhances level of GAP-43 mRNA after nerve injury and facilitates re-projection of the hypoglossal nerve[J]. Brain Res Mol Brain Res, 1995, 32 (2) :308-312.
����[12] Dallerac G, Rampon C, Doyere V. NCAM function in the adult brain:lessons from mimetic peptides and therapeutic potential[J].Neurochem Res, 2013, 38 (6) :1163-1173.
����[13] Ko HG, Choi JH, Park DI, et al. Rapid Turnover of Cortical NCAM1 Regulates Synaptic Reorganization after Peripheral Nerve Injury[J]. Cell Rep, 2018, 22 (3) :748-759.
����[14] Konno D, Ko JA, Usui S, et al. The postsynaptic density and dendritic raft localization of PSD-Zip70, which contains an N-myristoylation sequence and leucine-zipper motifs[J]. J Cell Sci, 2002, 115 (23) :4695-4706.
����[15]����, ����ΰ, �ƾ�, ��.ͻ����������ѧϰ�����ϵ���о���չ[J].����ҽѧԺѧ��, 2012, 27 (1) :89-92.
����[16] El-Husseini AE, Craven SE, Chetkovich DM, et al. Dual palmitoylation of PSD-95 mediates its vesiculotubular sorting, postsynaptic targeting, and ion channel clustering[J]. J Cell Biol, 2000, 148 (1) :159-172.
����[17] Wiedenmann B, Franke WW. Identification and localization of synaptophysin, an integral membrane glycoprotein of Mr 38, 000 characteristic of presynaptic vesicles[J]. Cell, 1985, 41 (3) :1017-1028.
����[18] Eastwood SL, Burnet PW, Mc Donald B, et al. Synaptophysin gene expression in human brain:a quantitative in situ hybridization and immunocytochemical study[J]. Neuroscience, 1994, 59 (4) :881-892.
����[19] Chou AK, Muhammad R, Huang SM, et al. Altered synaptophysin expression in the rat spinal cord after chronic constriction injury of sciatic nerve[J]. Neurosci Lett, 2002, 333 (3) :155-158.
����[20] Caleo M. Different rates of horseradish peroxidase transport in the optic nerve of neonatal and adult rats[J]. Neuroscience, 1996, 72 (3) :725-730.
����[21] Qian JJ, Cheng YB, Yang YP, et al. Differential effects of overexpression of wild-type and mutant human alpha-synuclein on MPP induced neurotoxicity in PC12 cells[J]. Neurosci Lett, 2008, 435 (2) :142-146.

ͻ������Ч��������ͻ����������ص���
ͻ����������ָͻ������̬�����ϵĸı䣬��ѧϰ�ͼ�����������ѧ����������ϵͳ�ķ��������켰ѧϰ����������Ҫ���á�ͻ���Ĵ��ݿ������Ǿ�������ͻ����������ؼ��IJ��֡������о�������ͻ���������밢���ĺ�Ĭ����AD��������֪�����½�������...
��ؼ��ضԼ����˶���Ԫͻ�������Ե�����
ͻ���������ձ������������ϵͳ�У���δ������ϵͳ�ķ�����������ѧϰ��������Ե����������ܻ������ء����������ڷ���ϵͳ���ӵ������õ���Ҫ����������ʣ�������-�ڷ���-��������ϵͳ���о���������Щ����Ҳ����ϵͳ�в��ɻ�ȱ�ĵ�...