��������ѧ����

����ժ Ҫ����ѪС�����ֹѪ��ά��Ѫ�������Ļ������ܣ��������о�֤������ϸ����Ⱦ�е�����Ҫ�����߷�����ɫ�����������ϸ����Ⱦʱ��ѪС���һ��Ǩ�㵽��Ⱦǰ�ߣ�ʶ���������̲�ԭ�����ͷŶ���ɱ���ﵰ��ֱ��ɱ��ϸ����ͬʱ��ǿ������ϸ��������ϸ��������ϵͳ�Ĺ������߹��ܣ���Tϸ����Bϸ��������Ӧ�����߹��ܣ�Э����ͬ�������ϸ����
�����ؼ��ʣ���ѪС��; ϸ��; ��Ⱦ; ����;
����Abstract����Platelet has the basic function of hemostasis and maintaining vascular integrity, recent studies have proved that it plays an important role in immune defense against bacterial infection. When bacterial infection occurs, platelet firstly migrate to the infection frontline to identify, bind, endocytose pathogens or release a variety of platelet microbicidal protein, which can directly kill bacteria, strengthen the innate immune function of neutrophil, monocyte and complement system, promote T lymphocyte and B lymphocyte to initiate adaptive immune responses against bacteria.
����Keyword����platelet; bacteria; infection; immunity;
�����ܶ�������������ڼ���������һ��Ѫϸ��ͬʱ����ֹѪ�����߷������ܡ��ڲ��鶯��Ľ��������У�ϸ�����ͼ��书�ܱ�ø������죬��ѪС�塢��ϸ�����ܰ�ϸ���ֱ�ִ��ֹѪ����֢�����ߵ��ڹ���[1]����ΪֹѪϸ����ѪС�壬�����������ǹ�ע������������Ѫ��Ѫ˨�Լ����Ĺ�ϵ��������������о���չ���ࡣ������1901��Levaditi�ͷ��ֻ��һ�������ѪС��ۼ����������ѪС����ϸ��ֱ�ӽӴ�����ع۵�[2],��һֱ�������о��ƺ����༯����ϸ����μ���ѪС�岢�շ�Ѫ˨��ֱ��1997�꣬Yeaman ��[3]�ٴ������֤ʵѪС���Ա�������ЧӦϸ���Ĺؼ��ṹ�����߷������ܣ��й�ѪС����Ϊϸ����Ⱦ��“��һ�ڱ�”���۲��ٴ��ܵ����ӡ���֮��ʹ�ø�ѪС��Ѫ��(platelet-rich plasma, PRP)��ѪС�������ϸ����Ⱦ��ʼ��Ϊ��Щ�����˵��о����������о�֤ʵ��PRP�Խ��ɫ����������ͼ������ֽ��ɫ��������������ù�س�����Լ������ײ�����ͭ�̼ٵ������������������Ե���������[4,5],��ʾPRP��ΪԤ�������пڸ�Ⱦ�����������˿ڻ�Ǹ�Ⱦ����Ч�������ơ�PRP�뿹��ҩ�����Эͬ���ã������ڿ���ҩ����ҩʱ���ж�������[6,7]��������ϸ���������߰����ڹ�עPRP,�����Խ���1���ڰ���������[8]���ɼ�ѪС�����ϸ���ĵ�λ�õ��Ͽɣ����ľ�ѪС�������Ч����ϸ�����о�����������

����1 ��ѪС����ص�
����ѪС�����ɾ�ϸ����������ϸ����������ѭ��������ԼΪ10 d, �ڴ��ڼ���Ծ�Ϣ��״̬����[1]����ȻѪС���ˣ������о�ϸ������mRNA,�ɶ����ϳɵ�����[9]��ѪС�廹����������صĽṹԪ�أ���ϸ����Ĥ�������ĸ��ַ������壬Ĥ�����γɵĿ�����ϵͳ(open canalicular system, OCS)�Լ�ϸ���������������ܿ�����α��������ø�������[10,11],����ѪС������ʹ�����ѪС������еķ��Ӻܴ�̶ȿ�����ѪС��Ĺ��ܡ�
����ѪС����溬�д������ǵ��ף���Щ�ǵ��ײ�������Ѫ˨���γɺ�ֹѪ��������������֢��Ӧ����ѪС�屻�����������ͨ�����䡢��Ĥ��������ںϱ����µ����ϵ�[12,13] ��ѪС����ڴ������ֿ������ֱ�Ϊα���������ܿ�������ø�����[14](����1),��ѧ���������һ��DZ�ڵ�T����[15]�� α�������������(80��/ѪС��),������(ֱ��200��500 nm),����Լ��300�ֻ��Ե��ף�����ɱ���ﵰ��(platelet microbicidal protein, PMP),��������������CXCL4��CXCL7�������RANTES(CCL5)�ȣ���ѪС��𤸽����Ѫ��ֹѪ�����к�Ѫ˨��֢���������ϡ�����������Ѫ�����ɺͿ��������������Ȳ�ͬ���̷�������[16]�������α���������ܿ��������������������/С(3��9��/ѪС�壬ֱ��250 nm),�������������(ADP)������������(ATP)���������Ρ������Ρ��鰷��Ѫ���ء������ӵȡ�ADP��һ�ֽ�����ѪС�弤������������ѪС����״�ı䡢�����ͷź;ۼ�[17]����ø��������е���ø������ø�;���ɱ�����Ե������ӵ��ף�Э������ϸ������[18]��
����2 ��ѪС�����ϸ����Ⱦ�Ļ���
������֯���Ĥ��������������룬������ԭ������Ѫ�������;����ѪС����Ѫ������������ϸ��֮һ��ϸ��������ж��ֲ�ԭ��ʶ�����壬�ܿ���ʶ��ԭ��[1]��ϸ����ѪС���ֱ�ӽ�ϣ�ѪС����������Ƥ�½�ԭ�Ľ�ϣ�����������Ƥ������Ѫ;�����ɵ���Ѫø�����ܻѪС�壬��һ������ѪС����κ��ѿ��������ӵ���ϸ����Ⱦ�Ĺ��ܡ�
������1 ѪС����Դ�ķ��Ӽ������߹���
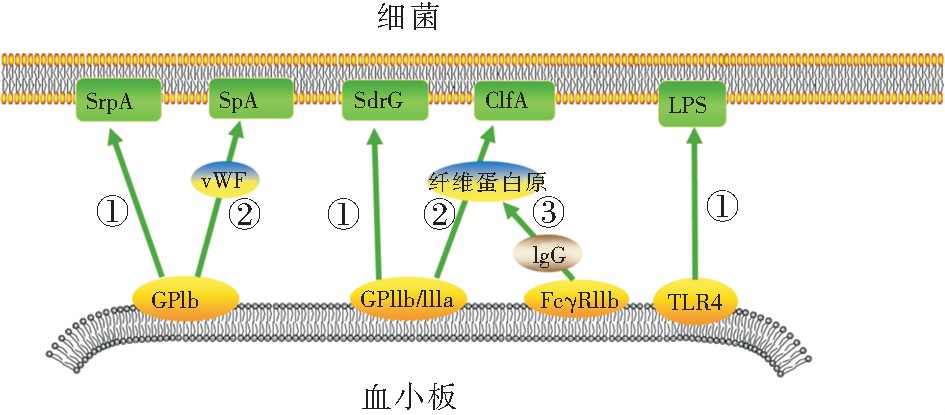
����2.1 ��ѪС��ʶ���ϸ��
����ѪС����ϸ��һ����ϣ����ն�����ѪС������������ѪС��ۼ����Ӷ�ʹѪС�����IJ�ԭ��ģʽ���������������������ǿ���������̼�ѪС��Ѹ��ʶ��ϸ����ϸ����ѪС����ϸ���Ľ��������������ʽ[19]:��ϸ���ͷŲ���Ӵ����ϵ�ѪС�塣��Ⱦ��ϸ���ͷ�һϵ��ϸ�����صȲ��ͨ��ֱ�ӽӴ���ı�ѪС��Ĥ�ṹ���ѪС�塣����ɫ��������ͷŵ�α���ؿɽ�ϵ�ѪС�����֬˫���Ӳ㣬������γɿ�Ĥ�ף�ʹ����Ca2+����������ѪС��[20];����߲�������������İ��װ��ᵰ������ѪС�����Ca2+���࣬��ʹѪС��Ѹ�پۼ�[21]����ϸ��ֱ����ѪС��������ϡ�ѪС�������ϸ��ʶ����ص�������GPIb��GPIIb/IIIa��TLR2��TLR4��gC1qR��FcγRIIa�ȡ�����ϸ��������ʶ���ֱ�ӽ�ϵ�ѪС��Ĥ�ϵ��������壬��ѪҺ���������˿���ᵰ��A(SrpA)�ɽ��ѪС��GPIb, ��Ƥ�������SdrG�ɽ��GPIIb/IIIa, ���������֬����B��������ɽ��TLR2,��ϣ����֬����(LPS)�ɽ��TLR4��[13,22,23,24]�� ��Ѫ��������ΪѪС���ϸ����ϵ���������ΪѪС����ϸ����ϵ����������� �о�������vWF����ά����ԭ�����������ͨ��vWF���ѪС��GPIb, ���������������A(ClfA)ͨ����ά����ԭ���ѪС��GPIIb/IIIa[23]����ͼ1��
����ͼ1 ѪС���ǵ�����ϸ�����浰�������ʾ��ͼ
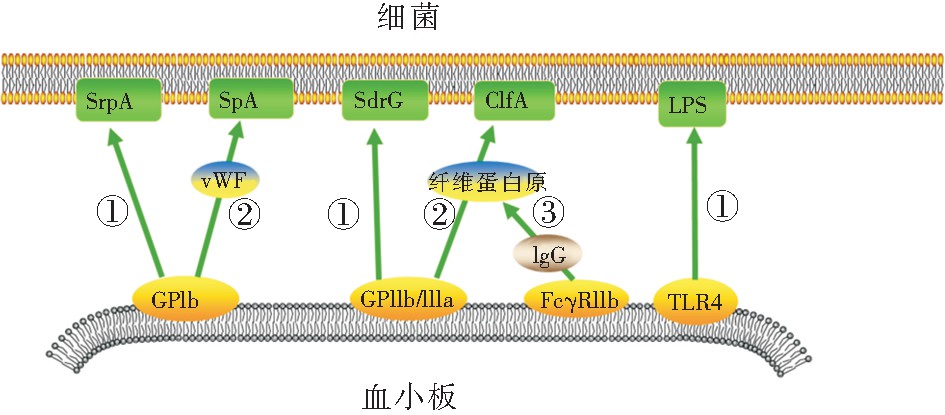
����ע����ѪС����ϸ��ֱ�ӽ�ϣ���ѪС��ͨ��vWF����ά����ԭ��ϸ����ӽ�ϣ���ѪС���FcγRIIa������IgG��Fc�ν�ϣ���ͨ����ά����ԭ��ϸ����ӽ�ϡ�
����2.2 ��ѪС������ϸ��
����ѪС��һ����ϸ����ϣ��ͻᷢ��һϵ�������ṹ���ܱ仯���������Ρ��������̡����ɣ����ɱ��ϸ��������1980��Clawson��[25]����ѪС��������̽��ɫ���������OCS��Youssefian��[26]��һ���о���֤ʵ����ϸ�����̣�������ѪС�����α�㽫ϸ����Χ�����ѪС����Ĥ���ݣ��γ������ݡ������ݿ���OCSĤ�ںϣ�ʹ����ϸ����������λ��OCS�С�Ȼ���������е�ϸ���ܷ�ɱ�𣬲����Ǻ������White��[27]�о���Ϊ��ѪС�������̽��ɫ���������OCS,������ɱ��ϸ������ΪѪС�岻�������������ø������ø��Ҳ��ѧ����ΪѪС��α���������������ںϣ����ͷſ�����������PMPs, �����������е�ϸ�������[26],�п���ɱ���������е�ϸ����Trier���������ѪС���ϸ�����л������������ѪС�����Ŀ��������������ɱ����������ѪС����ĿԶ���ڽ��ɫ�����������ʱ(1 000��),��ɱ��95%�İ���ϸ��[28],������ѪС������ɱ��ϸ���ġ����������Чɱ�������ڻ���ѪС��OCS�Ľ��ɫ��������Ƿ���Ϊ��ԭ����������ϵͳ��һ�ַ�ʽ��������Ϊϸ����“�ӻ���”ֵ�ý�һ���о���
����Ȼ������������ϸ����ѪС�嶼���������ڻ����á������о�֤��ѪС��������ϣ��(������ϼ������IgG),��ֻ�����̲�ɱ������IgG��ϵĴ�ϣ����������ѪС��FcγRIIa(��IgG����)�ͼ����������Ž鵼ѪС����ѽ��IgG�Ĵ�ϣ���������ɺ�ɱ�𣬸�ɱ��������������л������[29],˵��ѪС��FcγRIIa��ɱ��������ϸ�����������Źؼ����á�
����2.3�� ѪС���ͷ�PMPsֱ��ɱ��ϸ��
������ѪС�屻���ֵ�ϸ�������ѪС�弴�ѿ������ͷų�PMPs���������ӡ����������������ֱ��ɱ��ϸ����PMPs��һ��С�������Ӷ��ģ�ͨ����ѹ������ͨ��ֱ���ƻ���ԭ���ϸ��Ĥ������ϸ��Ĥ�Ķ����ϰ������Ըı�[30],�Ӷ�ɱ��ϸ���������о�[31]֤����PMPs��C��(���Ȼ���)�����ṹ�����ɱ�����ԣ���N�˾�����������(������);�������ֽṹ������Щ�������κı䣬���Ҳȷ����PMPs����ˮ����Ծ���ɱ�����ԡ�
����3�� ѪС����ǿ��������
���������Ⱦʱ��ѪС�����ȵ���Ⱦǰ�ߣ�һ��ʶ����ϸ����ϣ����������ã�ѪС���ɹ⻬Բ��״��ɶ�̵����Σ�ϸ���������ָ״��˿״α�㣬����Ч����ϸ������ʸ�������ϸ������������ϸ��[32](��ͼ3),ʹ����ϸ����������Ⱦ�ͬʱҲ�����ϵͳ���ӹ����������á�
����3.1�� ѪС��Эͬ������ϸ������
����ѪС��ͨ������������ͷ��������ӣ�������ļ��������ϸ������Ⱦ�ͬʱ�����ѪС�廹�ɴٽ�������ϸ���ͷŻ��������γɰ����ղ���(neutrophil extracellular trap, NET),������Ч�ղ�ϸ������ǿ���ɹ��ܣ���������“ϸ����������”ɱ��ϸ����
����3.1.1�� ѪС�弤���ļ������ϸ��
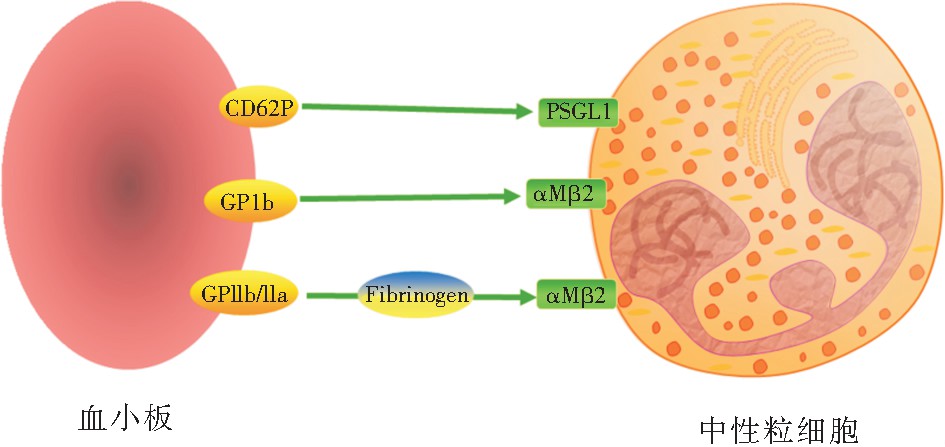
������ ѪС����Ҫͨ����������ͷ�����������ϸ�������(��ͼ2),����ѪС��CD62P�ɽ��������ϸ��Pѡ�����ǵ�������1(PSGL-1),ѪС��GPIb�ɽ��������ϸ����������αMβ2,�Լ�ѪС��GPIIb/IIIaͨ����ά����ԭ��ӽ��������αMβ2[33,34] ���� �ѪС���ͷ�CD40L���ϵ�������ϸ�������صı���ͷ�Ѫ��������������ϸ��ļ������֢��[35,36]�� �� ��Ѫ������ʱ��������������Ƥ������������ϸ��ļ�������𤸽����ȱʧ����������£�ѪС��GPIb-IX-V�������GPVIͨ��Ѫ������vWF𤸽����Ƥ�µĽ�ԭ��һ��𤸽��ѪС��������Pѡ���أ�Ϊ������ϸ��PSGL-1�ṩ���λ��[37],��ʱ������ϸ������������Ҳ���Խ�ϵ�ѪС��������������ӣ��Ӷ�ʹ������ϸ����Чļ����Ѫ�����˴�����ˣ���ȱ�����������𤸽��������£�ѪС��ɳ䵱������ϸ��ļ����“��½��”,��������������ϸ���������źš�
����ͼ2 ������ϸ��𤸽������ѪС��𤸽���Ӽ������ʾ��ͼ
����3.1.2�� ѪС����������ϸ�����������е�����
�������ͷŻ�������������ϸ������Ҫ�����Dz������ͷŻ�����(reactive oxygen species, ROS),ROS��ϸ�����Խϴ���ɱ��ԭ��[38]��ѪС����������ϸ���Ľ���ܴ������ROS������Ч�ʣ����ڹ��̵ĺ�������������˸ù����ֳ�Ϊϸ����������[39]�����������á���������ϸ���������ܲ�ԭ��(���߾�������߲��������)���о��з��֣�������ϸ��������Ч��������ѪС����������ϸ����������γɣ� 20 min�ڣ����ѪС��鵼������ϸ��������ϸ��Ч��Լ����20%[40]����ˣ�ѪС��ɴ����ǿ������ϸ�������ɹ��ܡ� ��NETs���γɡ�NETs��������ϸ���ͷŵ�Ⱦɫ�ʡ�����ˮ��ø����������������ɣ������ղ���ֱ��ɱ��ϸ��[41,42]����������ղ����IJ���ʹ������ϸ��ʧȥϸ���ˣ���ϸ���Ծ���Ѫ�������к�תλ�ȹ���[43];���DZ�������Щϸ���˶����ܣ�������NETs��ϸ���IJ��������������о���������ѪС�弤����(ADP����ԭ����Ѫø��������ϩ�ᡢTLR����)����ʱ���ɹ۲쵽ѪС������NETs���γ�(��ͼ3);ȱ����ЩѪС�弤����ʱ��NETs�����γ�[44],��ˣ������ѪС��������NETs�γɵı�Ҫ������������ѪС��Ļ�Ƿ�Ҳ��NETs�γɵ��Ⱦ��������в������
����3.2 ��ѪС��Эͬ���ո�ϸ������ϸ��
����ѪС����Ϊ�����ڱ�Ѳ����Ѫ����[45]���λ��������ʾ����״̬ʱ���ڸ�Ѫ����ڵ�ѪС��GPIb����ո�ϸ������vWF���ݽ�ϣ���ѭ�����γɶ��ݵ� “�Ӵ�-�ƶ�”����(����1 s);��ϸ�������Ѫ��ʱ�����ո�ϸ���ͻ�Ѹ�ٲ���ϸ������Ѹ�ٴ���ѪС����“�Ӵ�-�ƶ�”�Ķ���𤸽״̬ת��ΪѪС��GPIIb/IIIa����ո�ϸ������vWF��ϵij����ι�𤸽״̬���Ӷ�ʹ����ѪС��Ѹ�ٽ���Щϸ����Χ�������γ�ѪС��ۼ�Ȧ[46] (��ͼ3)������ѪС��ۼ�Ȧ���ȿ�ֱ���к�ϸ��[45],Ҳ�ɷ�ֹϸ��������ո�ϸ������һ����ǿ����ϸ�����������á����ƺ��Ǹ����һ�ּ��ӻ��ƣ���ѪС��ͨ��“�Ӵ�-�ƶ�”�������Ѫ���ھ���ϸ���Ļ�����ڲ�������ϸ���Ĵ��ԡ����⡢Ƥ����������֯�ں��ٿ���ѪС������ֶ��ݽ���[46]��CD40L������ϸ����һ����Ҫ�����������ǿ����ϸ�������ɹ��ܣ���ѭ���дֵ�CD40L�ɻ��ѪС�����[47],��ˣ����ѪС�����ǿ����ϸ������ϸ����Эͬ���ӻ���Ĺ������ߡ�
����ͼ3 ѪС������ո�ϸ����������ϸ���ڸ�Ⱦ�е������ʾ��ͼ
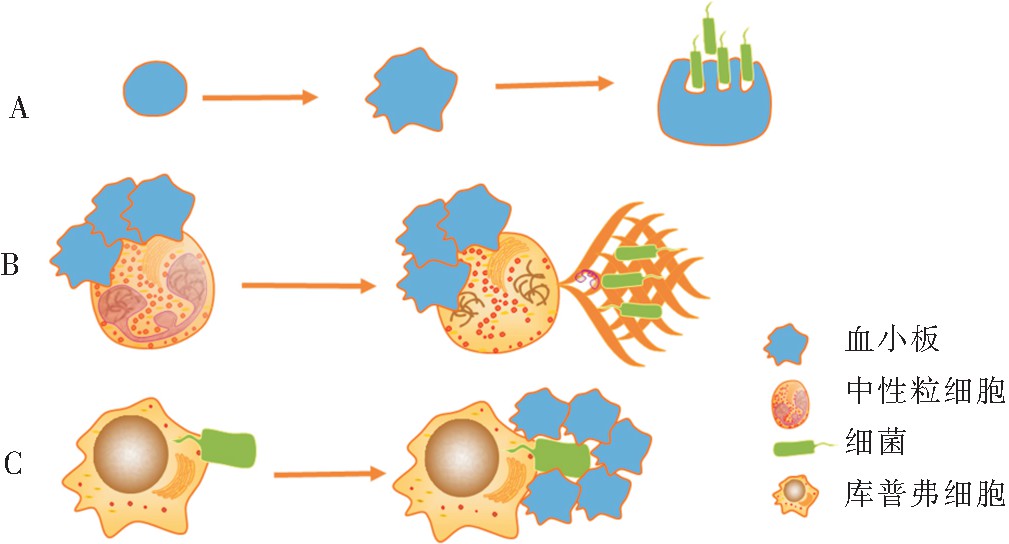
����ע��AΪѪС����ѪС��������α�㣬����ϸ��������ϸ�����������ϸ��������ϸ���������ɣ�BΪ���ѪС����������ϸ����Ϻ�NETs���γɣ�NETs��Ч���ղ�ϸ������ɱ��ϸ����CΪϸ������Ѫ�����ո�ϸ������ϸ�������ѪС��Ѹ���ڿ��ո�ϸ������Χ��ϸ����Χ������
����3.3�� ѪС����ǿ���������ЧӦ
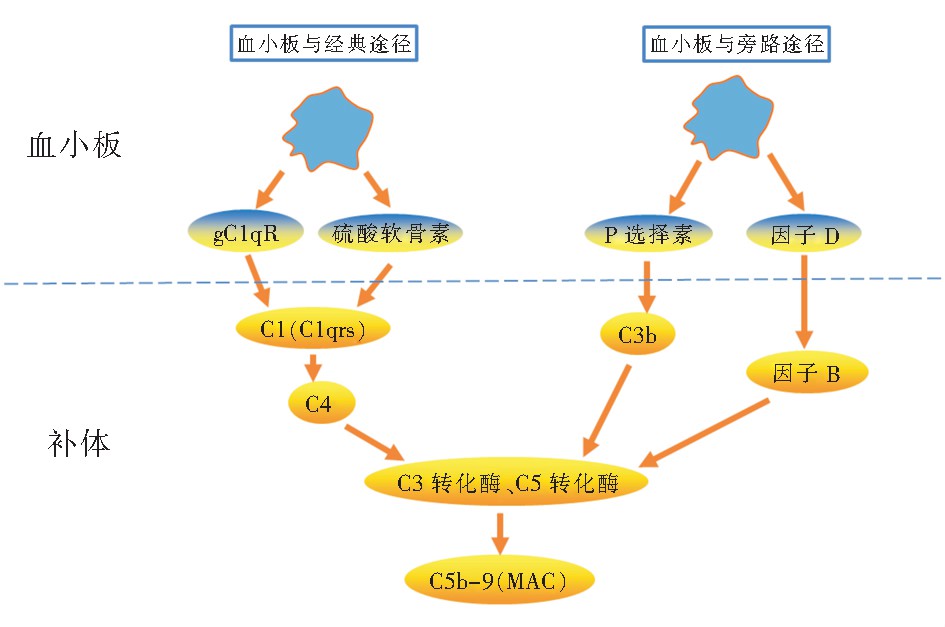
��������(complement, C)�ǹ�������ϵͳ����Ҫ��ɲ��֣�Ѫ���еIJ�����Էǻ��ʽ���ڣ�ֻ���ڱ�������������ѧ���ܡ����ѪС����ﲿ�ֲ������弰�������ӣ���ϲ��壬����ͨ����������;������·;������壬�ٲ��巢�ӿ������á�
����3.3.1 ��ѪС��ͨ������;�������
��������C1ͨ����C1q(C1r)2(C1s)2���ϴ������ʽ����Ѫ���С��о��������ѪС�����IJ�������gC1qR�ɽ�ϲ�����C1q, ���������C4(����������;�������)������ѪС����ڵ����������أ�ͨ�����C1q, ͬ��Ҳ��������;�������[48]����ͼ4��
����3.3.2 ��ѪС��ͨ����·;�������
������·;���ǴӲ���C3��ʼ��C3�ɱ�C3ת��ø�ѽ�ΪC3a��C3b����Դ�ѪС���Pѡ������C3b�����壻Pѡ������C3b��ϣ��ɴ̼���·;���ļ���ø�ٷ�Ӧ�����[49]�����ѪС��α������������D,����D�ܼ���ѽ�����BΪBa��Bb [50],�ٲ���ת��ø�γɣ�ͬʱ����C3ת��ø���������ѽ���ಹ��C3,��ǿ��·;������塣��ͼ4��
����3.3.3 ��Ĥ������������γ�
��������;������·;���������γ�C5ת��ø����C5�ѽ��C5a��C5b������֤��ϴ�ӡ����ѪС����ͷ�C8��C9,С������C5��C6��C7��C5b��C6��C7��C8��C9��ϣ��γ�C5b-9������ֳ�Ĥ����������(membrane attack complex, MAC)[50](��ͼ4)��MACͨ����Ĥͨ�����γ�ʹϸ���ѽ⣬�Ӷ��ѽ�ϸ����MACҲ��ʹ��Ƥ�ͷ�TF��vWF������Ѫ;�����շ���Ѫø��Ѫ˨���γɣ����ռ���ѪС�岢��ѪС���ѿ���[51] ����ˣ����ѪС����ǿ����ϵͳ�ѽ�ϸ���������������IJ���Ҳ��ǿѪС��Ļ���ѿ���������������ЧӦ������ǿ����ɱ�����á�
����ͼ4 ���ѪС���벹���������ʾ��ͼ
����4 ��ѪС����ǿ��Ӧ������
������Ӧ������(������Һ���ߺ�ϸ������)������T��B�ܰ�ϸ��������Դ���ʵĴ̼������������ֳ���ֻ�ΪЧӦϸ�������������ԭ�������ѧЧӦ�����ѪС���ֱ�ӻ���������T��B�ܰ�ϸ������������ǿ��Ӧ�����߷�Ӧ��
����4.1�� ѪС����T�ܰ�ϸ��
����һ����ѪС�����ʶ��ͽ��ϸ�������䴫����Ƣ���粹��C3���ε���˹�ؾ�������ѪС����GPIb��ϣ����ϸ����ѪС����Ƣ��CD8α+��ͻ״ϸ��ʶ�𣬽����ź�(��ԭ)��ʸ�Tϸ��������ϸ�����ߣ���һ���棬ѪС�屾����CD40L�ܴٽ���ͻ״ϸ�����������T�ܰ�ϸ�������߷�Ӧ[52];ͬʱ��ѪС���ͷŵ�Ѫ���غ�RANTES(CCL5),Ҳ�ܽ鵼Tϸ���ļ���ͷֻ�[53]��
����4.2�� ѪС����B�ܰ�ϸ��
��������ѪС����B�ܰ�ϸ���������3 d���֣�B�ܰ�ϸ��������������(IgG1��IgG2��IgG3)����[53],����ѪС���ܼ�������ѪB�ܰ�ϸ���ʹ������IJ�����ͬʱ��ʾѪС������дٻ��������Һ���ߵ�DZ�����á����⣬ѪС������Fcγ���壬�ܽ��IgG����[54],Э��������Ӧ�����߷�Ӧ��ɱ��ϸ����
����4.3�� ѪС�����ܰͽ�
�����ܰͽ���T��B�ܰ�ϸ�����ӵij��������ѪС�����C����������2����(CLEC-2),�ɽ���ܰ���Ƥϸ����ƽ�㵰�ף���ά���ܰͽᡢ�ܰܵ�������[55,56],������Ӧ������Ӧ��ķ�����
����5 ���ܽ�
����ѪС�������˵�ϸ����Ƭ������ϸ����Ⱦ���ӹؼ������߷������á����Ŷ�“ѪС����Ϊ������ϸ���ĵ�һ�ڱ�”��������о������룬��Ϊ�ٴ�����Ⱦ�ṩ�µ�˼·��
���������
����[1] Yeaman MR.Platelets:at the nexus of antimicrobial defence[J].Nat Rev Microbiol,2014,12(6):426-437.
����[2] Clawson CC,White JG.Platelet interaction with bacteria.I.Reaction phases and effects of inhibitors[J].Am J Pathol,1971,65(2):367-380.
����[3] Yeaman MR.The role of platelets in antimicrobial host defense[J].Clin Infect Dis,1997,25(5):951-968.
����[4] Hannachi N,Grac L,Baudoin JP,et al.Effect of antiplatelet agents on platelet antistaphylococcal capacity:an in vitro study[J].Int J Antimicrob Agents,2020,55(3):105890.
����[5] R?za Ayta? ?etinkaya,Ercan Yenilmez,Patrizio Petrone,et al.Platelet-rich plasma as an additional therapeutic option for infected wounds with multi-drug resistant bacteria:in vitro antibacterial activity study[J].Eur J Trauma Emerg Surg,2019,45(3):555-565.
����[6] Zhang WH,Guo Y,Kuss M,et al.Platelet-rich plasma for the treatment of tissue infection:preparation and clinical eva-luation[J].Tissue Eng Part B Rev,2019,25(3):225-236.
����[7] de Oliveira BGRB,de Oliveira FP,Teixeira LA,et al.Epidermal growth factor vs platelet-rich plasma:activity against chronic wound microbiota[J].Int Wound J,2019,16(6):1408-1415.
����[8] Mirzaei M,Daneshpajooh A,Farsinezhad A,et al.The therapeutic effect of intravesical instillation of platelet rich plasma on recurrent bacterial cystitis in women:a randomized clinical trial[J].Urol J,2019,16(6):609-613.
����[9] Rondina MT,Weyrich AS.Regulation of the genetic code in megakaryocytes and platelets[J].J Thromb Haemost,2015,13(S1):S26-S32.
����[10] Gremmel T,Frelinger AL 3rd,Michelson AD.Platelet physiology[J].Semin Thromb Hemost,2016,42(3):191-204.
����[11] Lam FW,Vijayan KV,Rumbaut RE.Platelets and their interactions with other immune cells[J].Compr Physiol,2015,5(3):1265-1280.
����[12] Montague SJ,Andrews RK,Gardiner EE.Mechanisms of receptor shedding in platelets[J].Blood,2018,132(24):2535-2545.
����[13] Deppermann C,Kubes P.Platelets and infection[J].Semin Immunol,2016,28(6):536-545.
����[14] Cristina Gomez-Casado,Alma Villase��or,Alba Rodriguez-Nogales,et al.Understanding platelets in infectious and allergic lung diseases[J].Int J Mol Sci,2019,20(7):1730.
����[15] Thon JN,Peters CG,Machlus KR,et al.T granules in human platelets function in TLR9 organization and signaling[J].J Cell Biol,2012,198(4):561-574.
����[16] Blair P,Flaumenhaft R.Platelet alpha-granules:Basic biology and clinical correlates[J].Blood Rev,2009,23(4):177-189.
����[17] Thon JN,Italiano JE.Platelets:production,morphology and ultrastructure[J].Handb Exp Pharmacol,2012,(210):3-22.
����[18] Rendu F,Brohard-Bohn B.The platelet release reaction:granules’ constituents,secretion and functions[J].Platelets,2001,12(5):261-273.
����[19] Hamzeh-Cognasse H,Damien P,Chabert A,et al.Platelets and infections-complex interactions with bacteria[J].Front Immunol,2015,6:82.
����[20] Arvand M,Bhakdi S,Dahlb?ck B,et al.Staphylococcus aureus alpha-toxin attack on human platelets promotes assembly of the prothrombinase complex[J].J Biol Chem,1990,265(24):14377-14381.
����[21] Lourbakos A,Yuan YP,Jenkins AL,et al.Activation of protease-activated receptors by gingipains from Porphyromonas gingivalis leads to platelet aggregation:a new trait in microbial pathognicity[J].Blood,2001,97(12):3790-3797.
����[22] �����࣬�����ƣ�������.B���������ͬ�����ѪС��Ļ����[J].�й�����������־��2016,32(2):333-338.
����[23] Kerrigan SW.The expanding field of platelet-bacterial interconnections[J].Platelets,2015,26(4):293-301.
����[24] Schattner M.Platelet TLR4 at the crossroads of thrombosis and the innate immune response[J].J Leukoc Biol,2019,105(5):873-880.
����[25] Clawson CC,White JG.Platelet interaction with bacteria.V.Ultrastructure of congenital afibrinogenemic platelets[J].Am J Pathol,1980,98(1):197-211.
����[26] Youssefian T,Drouin A,Massé JM,et al.Host defense role of platelets:engulfment of HIV and Staphylococcus aureus occurs in a specific subcellular compartment and is enhanced by platelet activation[J].Blood,2002,99(11):4021-4029.
����[27] White JG.Why human platelets fail to kill bacteria[J].Platelets,2006,17(3):191-200.
����[28] Trier DA,Gank KD,Kupferwasser D,et al.Platelet antistaphylococcal responses occur through P2X1 and P2Y12 receptor-induced activation and kinocidin release[J].Infect Immun,2008,76(12):5706-5713.
����[29] Riaz AH,Tasma BE,Woodman ME,et al.Human platelets efficiently kill IgG-opsonized E.coli[J].FEMS Immunol Med Microbiol,2012,65(1):78-83.
����[30] Yeaman MR,Bayer AS,Koo SP,et al.Platelet microbicidal proteins and neutrophil defensin disrupt the Staphylococcus aureus cytoplasmic membrane by distinct mechanisms of action[J].J Clin Invest,1998,101(1):178-187.
����[31] Yeaman MR,Yount NY.Unifying themes in host defence effector polypeptides[J].Nat Rev Microbiol,2007,5(9):727-740.
����[32] Gaertner F,Ahmad Z,Rosenberger G,et al.Migrating platelets are mechano-scavengers that collect and bundle bacteria[J].Cell,2017,171(6):1368-1382.e23.
����[33] Zuchtriegel G,Uhl B,Puhr-Westerheide D,et al.Platelets guide leukocytes to their sites of extravasation[J].PLoS Biol,2016,14(5):e1002459.
����[34] Zucoloto AZ,Jenne CN.Platelet-neutrophil interplay:insights into neutrophil extracellular trap (NET)-driven coagulation in infection[J].Front Cardiovasc Med,2019,6:85.
����[35] Rahman M,Zhang S,Chew M,et al.Platelet-derived CD40L (CD154) mediates neutrophil upregulation of Mac-1 and recruitment in septic lung injury[J].Ann Surg,2009,250(5):783-790.
����[36] Duerschmied D,Suidan GL,Demers M,et al.Platelet serotonin promotes the recruitment of neutrophils to sites of acute inflammation in mice[J].Blood,2013,121(6):1008-1015.
����[37] van Gils JM,Zwaginga JJ,Hordijk PL.Molecular and functional interactions among monocytes,platelets,and endothelial cells and their relevance for cardiovascular diseases [J].J Leukoc Biol,2009,85(2):195-204.
����[38] Rossaint J,Margraf A,Zarbock A.Role of platelets in leukocyte recruitment and resolution of inflammation[J].Front Immunol,2018,9:2712.
����[39] Jacek Miedzobrodzki,Tomasz Panz,Przemys?aw M P?onka,et al.Platelets augment respiratory burst in neutrophils activated by selected species of Gram-positive or Gram-negative bacteria[J].Folia Histochem Cytobiol,2008,46(3):383-388.
����[40] Assinger A,Laky M,Schabbauer G,et al.Efficient phagocytosis of periodontopathogens by neutrophils requires plasma factors,platelets and TLR2[J].J Thromb Haemost,2011,9(4):799-809.
����[41] Papayannopoulos V.Neutrophil extracellular traps in immunity and disease[J].Nat Rev Immunol,2018,18(2):134-147.
����[42] Zhang XZ,Zhuchenko O,Kuspa A,et al.Social amoebae trap and kill bacteria by casting DNA nets[J].Nat Commun,2016,7:10938.
����[43] Yipp BG,Petri B,Salina D,et al.Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo[J].Nat Med,2012,18(9):1386-1393.
����[44] Carestia A,Kaufman T,Rivadeneyra L,et al.Mediators and molecular pathways involved in the regulation of neutrophil extracellular trap formation mediated by activated platelets[J].J Leukoc Biol,2016,99(1):153-162.
����[45] Jenne CN,Kubes P.Platelets in inflammation and infection[J].Platelets,2015,26(4):286-292.
����[46] Wong CH,Jenne CN,Petri B,et al.Nucleation of platelets with blood-borne pathogens on Kupffer cells precedes other innate immunity and contributes to bacterial clearance[J].Nat Immunol,2013,14(8):785-792.
����[47] Soong L,Xu JC,Grewal IS,et al.Disruption of CD40-CD40 ligand interactions results in an enhanced susceptibility to Leishmania amazonensis infection[J].Immunity,1996,4(3):263-273.
����[48] Nording H,Langer HF.Complement links platelets to innate immunity[J].Semin Immunol,2018,37:43-52.
����[49] Deppermann C,Kubes P.Start a fire,kill the bug:the role of platelets in inflammation and infection[J].Innate Immun,2018,24(6):335-348.
����[50] Speth C,Rambach G,Würzner R,et al.Complement and platelets:mutual interference in the immune network[J].Mol Immunol,2015,67(1):108-118.
����[51] Carter AM.Complement activation:an emerging player in the pathogenesis of cardiovascular disease[J].Scientifica (Cairo),2012,2012:402783.
����[52] Iannacone M.Platelet-mediated modulation of adaptive immunity[J].Semin Immunol,2016,28(6):555-560.
����[53] Koupenova M,Clancy L,Corkrey HA,et al.Circulating platelets as mediators of immunity,inflammation,and thrombosis[J].Circ Res,2018,122(2):337-351.
����[54] Palankar R,Kohler TP,Krauel K,et al.Platelets kill bacteria by bridging innate and adaptive immunity via platelet factor 4 and FcγRIIA[J].J Thromb Haemost,2018,16(6):1187-1197.
����[55] Li JL,Zarbock A,Hidalgo A.Platelets as autonomous drones for hemostatic and immune surveillance[J].J Exp Med,2017,214(8):2193-2204.
����[56] Lax S,Rayes J,Wichaiyo S,et al.Platelet CLEC-2 protects against lung injury via effects of its ligand podoplanin on inflammatory alveolar macrophages in the mouse[J].Am J Physiol Lung Cell Mol Physiol,2017,313(6):L1016-L1029.

ѪС����ע���ٴ�����ѪС����ٻ��ܲ�ȫ�����³�Ѫ��Σ�ػ���ʱ,����ȱ�ٵ�һ�������ֶΡ�����ѪС��������Ч�á���Ѫ������Ӧ�����ٶ����ٴ�ҽ�������ѡ�á�����ѪС���Ӱ�����ؽ϶�,��Ҫ������Ѫ������ѪҺѧ�ĸı䡣Ϊ��һ���۲쳤�ھ���ѪС��Ի�...