
�������������˺��ܹ�����������ʵ�������٣��������������������������ġ������༲�������£����ڸ��ֲ������ص�Ӱ�죬������������������ȫ������ϸ�����ι��ܵ�ȱʧ����ʹ����ֲ�������Ʊ����Ը�˥��ĩ�����Ըβ�������ѡ���ڸ�Դ������ȱ�Ľ����о��������dz���Ҫ��
����1 ����������
����������������һ�����ӵĹ��̣��������ɵĽṹ���ӣ����Ҳ���������ϸ�����Ͷ����������������У���ϸ�������Ƚ��з��ѵ�ϸ�����ͣ��ڸ������ӵĴ̼��£�ϸ����������������йصĻ����� G0 �ڽ�������ڣ�ͨ���� 2 �� 3 d�ھ������ 1 �� 2 ��ϸ���������ڡ������״ϸ����Kupffer ϸ���͵�����Ƥϸ���Ⱥ����ϸ���������ڡ�ͬʱ��Ѫ����Ƥϸ����ֳ������Ѫ���������������ؽ������Ѫ�ܽṹ�����⣬��������ἤ�����ϸ��( hepatic progenitor cell��HPC) �������ܹ��ֻ�Ϊ��ϸ������Ƥϸ��������ϸ������������������������������ʱ�������ϸ���⽫�����������ԭϸ��������״̬����ԭϸ���������٣�����һ���������Щλ��������Χ����Բϸ����������ֳ�������о�֤ʵ�����ָ��г���Լ 22 d����ԭϸ�����ɵĸ�ϸ���͵�����Ƥϸ�����ָ�ԭ�еĸ���֯�������������Ĺ����ܵ��������ص�Ӱ�죬���� Notch �ź�ͨ·�����漰����ϸ������ֳ�ͷֻ������� Notch �ź�ͨ·�ڸ������е����ý������ܵ�Խ��Խ������ӡ�
����2 Notch �ź�ͨ·
����Notch ָ��Ӭ��Եȱ��( notch) ���ͣ����ִ��Ŵ�ѧ�ĵ����֮һ Morgan �ڹ�Ӭ�Ĵ��ģͻ���о������ȼ������ִ���������ѧ�о���������Ӭ Notch Ϊһ����Է�������( Mr) Լ 300 000 �ĵ��� 1 �Ϳ�Ĥ���嵰�ף��� 2 ����ͨ�������������ɡ�Notch �ź�ת��ͨ·�����塢����� DNA ��ϵ��� 3 ������ɡ�Notch �źŴӷ��ڵ��ͷų� Notch ���ڶι����� 3 ���ѽ⣬�ֱ𱻳�Ϊ S1��S2 �� S3 �ѽ⣬�ڷ��� S1�ѽ���γɵ���Դ�����壬���� 4 ������( Notch 1 �� 4) ��Notch ������( Delta ���嵰��) ���� 5 �࣬�ֱ�Ϊ Dll-1��Dll-3��Dll-4��Jagged-1 �� Jagged-2��Ҳ���ڿ�Ĥ���ס�Notch �����������Ϻ�������������������α ת��ø/ȥ������-��������ø( tumor necrosis factor-α converting enzyme/a disintegrin andmetalloprotease��TACE / ADAM) ������ S2 �ѽ⣬�γɵĺ����Ȼ�ĩ �� �� �� �� Notch ϸ �� �� �� �� ( Notch extracellulartruncation��NEXT) �� γ ����ø( γ-secretase) �����·��� S3 �ѽ⣬�Ӷ��ͷų� Notch ���ڶ� ( Notch intracellular domain��NICD)��NICD ���ж�����ܽṹ��: �ӽ�Ĥ������Ϊ�����ź����н�ϵ��� Jkappa ( recombination signal sequence-binding protein Jkappa����BP-J ) ��ط��� ( ��BP-J associationmolecule����AM) �ṹ��CDC10 / ankyrin �ظ����У�ת¼����ṹ���Լ�������-�Ȱ���-˿����-�հ��� ( proline-glutamate-serine-threonine��PEST) �ṹ��ȡ���AM �ṹ��鵼 NICD��ת¼���ӣ�BP-J/Su( H) �Ľ�ϣ��� CDC10/ankyrin �ظ����к�ת¼����ṹ���� NICD ��ת¼���������йء�Notch �źŵ����ΰе������������-������( basic helix-loop helix��bHLH) ���ף��緢״���������ǿ�� 1 ( hairy and enhancer ofsplit-1��HES-1) �� HES-5�������ܹ�ͨ�������� bHLH ���ӵȷ�ʽ�谭ϸ�������Էֻ�ЧӦ����ı������Ӱ��ϸ���ķֻ�����ֳ�͵�������Ϊ ��BP-J �������ź�ͨ·��������ѧ������ģ�BP-J���������ź�ͨ·Ŀǰ��û��������о���
����3 Notch �ź�ͨ·�������
����о���֤ʵ Notch �ź�ͨ·��������������ɣ������ڸ������о�����Ҫ����( ͼ1) �����ȣ�Notch �ź�ֱ�Ӳ�����������ڸ�����ֲ�� 2-����������( 2-acetylaminofluorine��2AAF) �������ٽ���70% �IJ��ָ��г�( partialhepatectomy��PH) ���Ʊ� 2AAF-PH ģ�͡����ø�ģ�ͣ��о��߷��� 2AAF-PH ��11 d�����������Բϸ����ͬʱ���� Notch-1 ��������ߡ����ʹ�� γ ����ø���� Notch �źţ���Բϸ���ķֻ����ܵ����ƣ��������ɵĸ�ϸ������Ҳ�ܵ�Ӱ�졣Wakabayashi�ȷ���С�ָ��г��� 3 h�����ɳ��� Notch1 ת¼�����ߣ���Ϊ���� Notch1 �ı����������������½���K��hler���Լ� Wang �ȵ��о�Ҳ֤ʵ Notch ͨ·�ڸ������о�����Ҫ���á����⣬Notch �ѽ��İ������ͨ����ʽ���̵ķ�ʽ�����ϸ�����Ӷ���߸�ϸ���ڵ� Notch ����������Ͷ����о���֤ʵ Notch �ź��ڵ��ܰ����ܹ����е���������Ҳ������Ҫ���á���Σ�Notch �źŲ��������������ܡ��о����֣�Notch �ź�ͨ���ı�θ���ת¼���ӵı�����Ƹ�ĸϸ���������ϸ����ϸ����ת�ֻ�����������ϸ������� Jagged-1 �����ܹ�ʹ��Χ��ĸϸ���еĵ��������Ա����( �� Sox-9 �� HNF1β ��) �ϵ���ͨ������ NICD ���Ƹ���ϸ�����ϸ������ֻ���ʹ��Щϸ���������͵ĵ�����Ƥϸ����ʹ�� Notch �ź����Ƽ������Ƹ���ϸ���γɵ���ϸ�����͡��ٴ����֣����������ܼ���ʱ HPC �ͷ�Ӧ�Ե���ϸ���е� Notch �źŷ�����������ȫ���� Notch�źţ������������ϸ���ֻ��� HPC����� Notch ͨ·��������ϣ����п��ܲ�����ϸ���ֻ��� HPC�����Dz������γ������ĵ��������⣬����Ѫ����Ƥϸ��Ҳ����Notch ���壬�����ڸ����������в������ѭ�������ܡ�
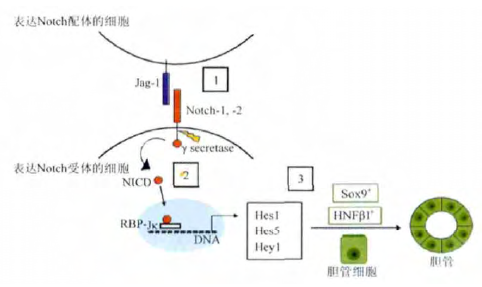
����Notch �� 4 �������ڸ����ж��б��Notch-1 �� Notch-2��Ҫ�����ڵ���ϸ���� HPC������������ʱ�������������ߣ�Notch-3 �� Notch-4 ��Ҫ��������ϸ��������ѧ���ԣ���Ƥϸ����Ҳ�б�����������������ΧѪ����Ƥϸ���ϵ� Notch ����( ��Ҫ�� Notch-2 �� Notch-3) �����������ߣ��������쳣������Ѫ�������йء�Notch �źŵ� 4 ������Ը�ϸ�����ܵ�Ӱ���кβ�ͬ��Ortica ���״ζԴ˽������о���������� Notch �ź�ͨ·��δ�ֻ�״̬�µ� HPC ��ֳ����Ҫ���ã����� Notch1��2��4 ���������ֳ�дٽ����ã���Notch3 �������������á���Щ���������Notch �źŵı���������������͵IJ�ͬ�����ֲ�ͬ�ĸı䣬���� Wnt ��Hedgehog �������ź����ϣ��Իָ��������̬���ܡ�
������Ѫ���Ƥϸ��( liver sinusoidal endothelial cells��LSEC)�Ǹ�Ѫ�����ڲ����е���Ƥϸ�������˲������ѭ�����³´�л֮�⣬��ͨ�����ϸ��֮�临����ϵ֧�ָ���ķ�����������С�ָ��г�ģ����ʾ���ڸ������У�һ�����ϸ����ֳ�γ���Ѫ�ܵ�ʵ�ʵ���ͨ��Ѫ����Ƥϸ����������( vascular endothelial growth factor��VEGF) ������ LSEC����һ���棬LSEC ����ͨ�� VEGF ���� 1( vascular endothelialgrowth factor receptor1��VEGF��1 ) �źŶԸ�ϸ�������������ã���������������Ѫ�������ء��о�֤ʵ����BP-J �鵼�� Notch �ź�ͨ·��ͨ������ LSEC Ӱ�������������Wang �ȵ��о��У�����ʹ�� ��BP-J �ó�С��ģ�ͽ����о������� ��BP-J ȱʧ��ʹ�ø�������ྲ����������( ��Ѫ�������ά���ʶѻ���Disse ��϶ˮ��ϸ����������) ����Ѫ�ṹ�쳣������ VEGF��1 ���オ�ͣ����� LSEC ����ֵ���ӣ�����Ϊ��ϸ���ṩ��ϸ���������� ( hepatocytegrowth factor��HGF) ��Ӫ�����ӣ�����������������ı䡣
�����ɴ˿ɼ���Notch/��BP-J �ź�ͨ·�Ը���������Ӱ����ܲ��ֹ鹦�� LSEC �е� VEGF��1-HGF �źš�VEGF�� �� LSEC ��ϸ�����е������ã�VEGF��2 ������� LSEC ����ֳ�źţ��� VEGF��1 ���� VEGF��2�����ҽ鵼��ϸ����ϸ������ 6( interleukin-6��IL-6) �� HGF �IJ�����
������һ���棬�������о���ʾ ��BP-J �鵼�� Notch �ź�ͨ·�� 2 �ֲ�ͬ��Ⱥ����Ƥ��ϸ��( endothelial progenitor cells��EPC) ������ EPC( early endothelial progenitor cells��EEPC) �ͺ��� EPC( endothelial outgrowth cells��EOC) �ݾ��в�ͬ�ĵ������ã��Ӷ�Ӱ������������о�����ʾ����BP-J ��ȱʧ��Ӱ��EEPC �Ĺ鳲���� PH ���С����� EEPC ��ע���������EEPC �� PH �յ��ĸ������нϺõ����á�EEPC �� EOC ��PH �յ��ĸ������в�ͬӰ�죬EEPC �ܹ��ٽ���ϸ����ֳ�����ٸ�ϸ��������EEPC ����Щ�����ܵ� Notch-��BP-J �ź�ͨ·�ĵ��ڡ�
����
����4 �������� Jagged-1 ;�����о�
���������е� Notch ����ֻ�� Jagged-1 �� Dll-4�����������Jagged-1 ������ HPC �͵���ϸ���Լ��ž������ʵ�ƽ����ϸ�����ڵ��ܰ��������У������ϸ��ͨ�� Jagged-1 �����Notch �ĸ�ĸϸ���Э�����ڵ������������У����ż����ϸ������ Jagged-1�����ĸϸ���ϵ� Notch-2 ��������ã��������뵨�������йصĻ����������ϸ���ֻ��йػ��Ӷ��ٽ������������� Jagged-1 ����� Notch-2������ͻ�䣬�������������� 1β( hepatic nuclear factor1β��HNF1β) ȱ�������µ���ϸ����Ӧ���ε�ϸ�����ۼ���Ӱ�쵨���������Ӷ��γ� Alagille �ۺ���( Alagillesyndrome��AGS) ���ٴ�����Ϊ���صĵ��ܷ��������͵�֭�ٻ��Լ����ָ�����֡���������� Jagged-1 �ı����������ߣ��Ǽ��� Notch �źŵ���Ҫ���塣K��hler ���ڶ�С����в��ָ��г����� 4 d �� Notch1 �� Jagged-1 ���ڸ�ϸ����������ڣ�NICD �ĺ�Ǩ���� 15 min �����Ӳ��ﵽ��ֵ���ڼ����� Notch ͨ·��HES-1 �ı����� 30 �� 60 min ���߲���60 minʱ�ﵽ��ֵ�������ָ�ϸ��ͬʱ���� Notch �� Jagged���ֵ��ף����߿���ͨ�������ڻ�·���Ƽ���ijЩ����ͨ·��Ҳ����Ӱ���ϸ������������ϸ�����ڸ�ϸ���� Jagged ����̼��£����ָ��г� 72 �� 144 h ���Ѫ���Ƥϸ����ʼ���������ĸ���֯�ڡ�������ͨ���о�Ҳ֤ʵ�ڸ��౻�����г���Notch/Jagged �ź�ͨ·���Դٽ����ܵ��γɺͽṹά�֣�����������Ѫ�ܵ��γɼ���ϸ������ֳ�������о����֣�Jagged-1 �Ǹ�ϸ������˿����ԭ���ڲ��ָ��г� 2 d�������Ĭ Notch �� Jagged-1�����������Ƹ��г����2 ��4���ϸ������ֳ���ڶ� Notch �����������о���ʾ��Notch ����� HES-1 �ı������ڼ����ߣ����� 60 min �ﵽ��ֵ��12 h �ָ�����ˮƽ��HES-5 �� 1 �� 6 h �������ߣ���12 �� 48 h֮������ 85% �����ѡ���Գ�Ĭ Notch-1 �������ϸ���������������ܵ�Ӱ�졣EPC �������ڲ�����Ѫ�����ɺʹٽ���֯���������������֮���н�Ϊ���еĹ�ϵ������ Notch �ź�ͨ·�� Jagged-1 �� EPC �ĵ������˺������á����о���ʾС�ָ��г���ĸ����������� Jagged-1�����н�Ϊ���Եı仯���� Jagged-2 �������Ҳ�������ˣ�Notch �� Jagged-1 ;��һ����ͨ���ٽ���ϸ������˿���Ѻ���ֳ�ٽ�����������һ��������п���ͨ���� EPC �ĵ��ڴӶ��ٽ���������
����5 С����չ��
�����������ڽ�����༲�����漫��DZ���������ܵ����ӣ����� Notch �ź�ͨ·�����еĹ�ϵ��Jagged-1 ;��ͨ�����Ӹ�ϸ������˿���Ѻ���ֳ���� EPC ��������ڴӶ��ٽ�����������BP-J ;��������� LSEC �Լ�ͨ�� CXC��4 �������EEPC ��ԱǨ�ƴٽ���������Ȼ������һ������о�Ŀǰ���������Σ�EPC ��Ѫ�ܷ�������֯�����еľ�����ӻ��Ʋ�����Notch ͨ·��ε��� CXC��4 �ķ��ӻ�����δ�������������о�δ�����롣��ˣ���δ�����о��У������о��������룬�������� Notch �ź�ͨ·���������ӻ��Ʊػ�չ�֣�����������ٴ�Ӧ�ôӶ���ø���ļ�ֵ�����档
���������:
������1��DeLeve LD�� Liver sinusoidal endothelial cells and liver regeneration��J�ݣ� J Clin Invest��2013��123( 5) : 1861 ��1866��
��2��Fiorotto �ң���aizner A��Morell CM��et al�� Notch signaling regulatestubular morphogenesis dur-ing repair from biliary damage in mice��J�ݣ� J Hepatol��2013��59( 1) : 124 ��130��
��3��Newsome PN��Hussain MA��Theise ND�� Hepatic oval cells: Helpingredefine a paradigm in stem cell biology��J�ݣ� Curr Top Dev Biol��2004��61: 1 �� 28��
��4��Greenwald I��Kovall �ң� Notch signaling: genetics and structure��J/OL�ݣ�Worm Book��2013: 1 �� 28��
��5���� �࣬�� �裮 Notch �ź�ͨ·��Ѫ�ܷ�����J�ݣ� ϸ�����������ѧ��־��2008��24( 12) : 1225 ��1227��
��6��Baker NE�� Patterning signals and proliferation in Drosophila imaginaldiscs��J�ݣ� Curr Opin Genet Dev��2007��17( 4) : 287 �� 293��
��7��Darwiche H��Oh SH��Steiger-Luther NC��et al�� Inhibition of Notchsignaling affects hepatic oval cell response in rat model of 2AAF-PH��J/OL�ݣ� Hepat Med��2011��3: 89 ��98��
��8��Wakabayashi N��Shin S��Slocum SL��et al�� ��egulation of notch1 signaling bynrf2: implications for tissue regeneration��J / OL�ݣ� Sci Signal��2010��3( 130) : ra52��
��9��K��hler C��Bell AW��Bowen WC��et al�� Expression of Notch-1 andits ligand jagged-1 in rat liver during liver regeneration ��J�ݣ�Hepatology��2004��39( 4) : 1056 �� 1065��
��10��Wang L��Wang CM��Hou LH��et al�� Disruption of the transcriptionfactor recombination signal-binding protein-Jkappa ( ��BP-J) leads toveno-occlusive disease and interfered liver regeneration in mice��J�ݣ�Hepatology��2009��49( 1) : 268 �� 277��
��11�� Parks AL��Klueg KM��Stout J�ң�et al�� Ligand endocytosis drivesreceptor dissociation and activation in the Notch pathway ��J�ݣ�Development��2000��127( 7) : 1373 �� 1385��
��12��Zong Y��Stanger BZ�� Molecular mechanisms of bile duct development��J�ݣ� Int J Biochem Cell Biol��2011��43( 2) : 257 ��264��
��13��Zong Y��Panikkar A��Xu J��et al�� Notch signaling controls liverdevelopment by regulating biliary differentiation��J�ݣ� Development��2009��136( 10) : 1727 �� 1739��
��14��Yanger K��Zong Y��Maggs L�ң�et al�� ��obust cellular reprogrammingoccurs spontaneously during liver regeneration ��J�ݣ� Genes Dev��2013��27( 7) : 719 �� 724��
��15�� Boulter L��Govaere O��Bird TG��et al�� Macrophage-derived Wntopposes Notch signaling to specify hepatic progenitor cell fate inchronic liverdisease��J�ݣ� Nat Med��2012��18( 4) : 572 �� 579��
��16��D’Amore PA��Ng YS�� Won’t you be my neighbor? Local induction ofarteriogenesis��J�ݣ� Cell��2002��110( 3) : 289 �� 292��
��17��Ortica S��Tarantino N��Aulner N��et al�� The 4 Notch receptors playdistinct and antagonistic roles in the proliferation and hepatocyticdifferentiation of liver progenitors��J�ݣ� FASEB J��2013 Oct 21����Epub ahead of print��
��18��Diehl AM�� Neighborhood watch orchestrates liver regeneration��J�ݣ�Nat Med��2012��18( 4) : 497 �� 499��
��19��Fausto N��Campbell JS����iehle KJ�� Liver regeneration��J�ݣ� Hepatol��2006��43( 2 Suppl 1) : S45 �� S53��
��20��LeCouter J��Moritz D�ң�Li B��et al�� Angiogenesis-independent endothelialprotection of liver: role of VEGF��-1��J�ݣ� Science��2003��299( 5608) :890 �� 893��
��21��Chen JY��Feng L��Zhang HL��et al�� Differential regulation of bonemarrow-derived endothelial progenitor cells and endothelial outgrowthcells by the Notch signaling pathway��J / OA�ݣ� PLoS One��2012��7( 10) : e43643��
��22��Morell CM��Fiorotto �ң�Fabris L��et al�� Notch signalling beyond liverdevelopment: Emerging concepts in liver repair and oncogenesis��J�ݣ�Clin ��es Hepatol Gastroenterol��2013��37( 5) : 447 �� 454��
��23�� Lemaigre F��Zaret KS�� Liver development update: new embryomodels��cell lineage control��and morphogenesis��J�ݣ� Curr OpinGenet Dev��2004��14( 5) : 582 �� 590��
��24��Fabris L��Cadamuro M��Guido M��et al�� Analysis of liver repairmechanisms in Alagille syndrome and biliary atresia reveals a role fornotch signaling��J�ݣ� Am J Pathol��2007��171( 2) : 641 �� 653��
��25�ݣ�oss MA��Sander CM��Kleeb TB��et al�� Spatiotem-poral expressionof angiogenesis growth factor receptors during the revascularization ofregenerating rat liver��J�ݣ� Hepatology��2001��34( 6) : 1135 �� 1148��
��26������꣬��ܣ�����壬�ȣ� �β����г���������� Notch/Jagged �źż� TNF �� IL-1 �ı����J�ݣ� �л��ε������־��2008��14( 10) : 732 �� 734��
��27�� Croquelois A��Blindenbacher A��Terracciano L��et al�� Inducibleinactivation of Notch1 causes nodular regenerative hyperplasia in mice��J�ݣ� Hepatology��2005��41( 3) : 487 ��496��
��28�� Xie J��Wang W��Si JW��et al�� Notch signaling regulates CXC��4expression and the migration of mesenchymal stem cells��J�ݣ� CellImmunol��2013��281( 1) : 68 �� 75��


