��������ѧ����


������ƭ��һ��������ձ���ڵ�����һֱ�������ܵ��㷺ѧ������Ĺ�ע[1,2], �����ſ�ѧ�����ĸ���, ��ƭ�о�Ҳ�����������ķ�չ. ����Ĵ�ͳ�о���Ҫ���öർ�Ǽ���, �ü���ͨ����¼�����(��Ƥ�����衢Ѫѹ������Ƶ��)�ı仯�������ƭ��Ϊ. ���������������ƭ����ֱ����ϵ, ��һ������������. �����¼���ص�λ(event relatedpotential, ERP)����֪��ѧ�е�Ӧ��, �о�������ʼ��ע��ƭ��صĴ��Ի, ������ ERP �ĵͿռ�ֱ�������, ��ƭ���Ի�����Ȼ������ȷ. ���������ֵĹ��ܺ˴Ź���(functional magnetic resonanceimaging, fMRI)����ͬʱ�߱����Ժ߿ռ�ֱ�������, ������ֱ��̽������ƭ�йصĴ��Ի[1], Ϊ��ʾ��ƭ�������ṩ����Ҫ����.
����Langleben ����[3]�״β��� fMRI ����̽����ƭ������ . ����ʹ�ø������GKT(guiltknowledge test)��ʽ�շ���ƭ��Ӧ�ͳ�ʵ��Ӧ, ���Ƚ����ַ�Ӧ�Ĵ��Լ���ģʽ. �������, ���ʵ��Ӧ���, ��ƭ��Ӧ������ǰ�۴��غͶ��ϻصȲ���ִ�п��ƹ��̵������ĸ���, ֤ʵ�� fMRI �ź��ڱ����ƭ�ͳ�ʵ��Ϊ�ϵĿ�����. ���ͬʱ, Spence����[4]�� Lee ����[5]�ֱ���ù������Ӧ��ʽ�ͼ�װʧ�䷶ʽҲ֤ʵ�� fMRI �ź��ܹ�������ƭ��Ӧ�ͳ�ʵ��Ӧ. ����Щ�о��Ļ�����, �����о���ʼ���첻ͬ���͵���ƭ��Ϊ�������ϵ���ͬ[6~10]; ��������̽����Ϊ��������ƭ�ɷ�(�������ɷַ���)[11~15]; �����Խ�ʾ�Է���ƭ��Ϊ����������ƭ��Ϊ�ĸ������[16~19]. ��Щ�о��Ľ����ձ���Ϊ,��ʵ��һ�ֻ���ʽ�ķ�Ӧ, ����ƭ����Ҫ�����ִ�п��ƹ��ܵIJ���, ��˻��漰����ִ��ϵͳ�ļ���, ��Ҫ����ǰ��Ҷ����Ҷ��ǰ�۴��ص�[20]. ����,���һЩ�о����˼ʽ���������龳�¿�����ƭ��Ϊ, �������, �������֪����й���(�罱��Ԥ�ڡ����������������жϺ��������۵�)��ص�����Ҳ������ƭ[16,21,22], ˵���ڸ�Ϊ��ʵ������龳��, ��ƭ���漰���Ӹ��Ӷ�������������[23], ����֮, ��ƭ�����ƻ��ܵ�����龳���ص�Ӱ��[20,23,24].
�������ܻ��� fMRI ����ƭ�о��Ѿ�ȡ����һЩ�ɹ�,��Ҳ������������. ����, ��Ȼ�����о���ʶ��������龳����ƭ(����)������ҪӰ��[20,23,24], ����, ���������Ӱ����ƭ���̵ľ�����Ʋ������. ����,��ƭ����������ƭ������龳�ı�Ҫ����, ������ƭ���̵�Ӱ������ȴ�ܵ��ܴ�ĺ���. ����ƭ����Ҫ���ٽ�����龳��, ����������й���ƭ����ʵ����[23]. Ŀǰ���������о����þ�����ƭ�������ʵ�鷶ʽ(���н�ɫ�ֹ�����Ϸ��ʽ)̽����ƭ���̵�����[16,17,21], �������, ���˾���Ĵ���ִ��ϵͳ�м�����, �����⼤�����������֪����й�����ص�����[16,21,22]. ����Щ�о���û�в�����ƭ���������, ������϶���Щ���������ļ����Ƿ�������ƭ����ļ��������. ��֮��Щ�о�ʹ����������ƭ�о���ȫ��ͬ����Ϸ��ʽ, ����Ҳ��ͨ�����о��ıȽϷ������ƭ�������ƭ���̵�Ӱ��ЧӦ. Ϊ�˽����Щ�ɻ�, ���о�ֱ�Ӳ�����ƭ�����һ����, ��ϸ�����С�����ƭ�������ƭ���̵����Ƶĵ�������.
�������о�����һ��"ɱ����Ϸ"ģ��ɱ���龳, Ȼ������鱻��("ɱ��")�ֱ�ʹ���С������� GKT��ʽ���в��, ������ָ����߶Ա���ÿ����ƭ��Ӧ���ʵ��Ӧ���ж����(����һ��Ӧ�ж�Ϊ��ʵ����ƭ), ��ͨ���ӷֻ�۷ֵķ�ʽ��ʵ������е���ƭ��Ӧ���ʵ��Ӧ�����������(�ۻ�������ܷ�ֱ�Ӿ�������߶Ա������ݵ������ж�), �ڿ����γ�������Ե���ƭ���. ͨ���Ƚ����鱻�Խ�����ƭ��Ӧ�ͳ�ʵ��Ӧ�Ĵ��Լ���ģʽ, ��ʾ��ƭ�������ƭ���̵����Ƶĵ�������.
�������о�Ԥ��: (��) �����������, �з����鱻����Ҫ�������ٲ���ߵķ���������������״̬,�Լ�ʱ����ӡ������IJ��ԡ��ٳɳɹ���ƭ, ���ʵ��Ӧ����ƭ��Ӧ��������̶ȵؾ����������ۼӹ�����, ��˿��ܻᵼ����������ĸ���, �米��ǰ�۴�Ƥ��(dorsal anterior cingulate cortex, dACC);(��) �����������, ����Ҫ������ƭ������з�������, ��ƭ���̽�����̶ȵ��漰������Ԥ�ںͷ�����������, �������ƭ�������漰���ڶ�������, ������Ԥ�ںͷ�����������״��(striatum,STR)���Ե�(insula, INS)�������ܵ��С�����(��ƭ���)���صĵ�������.
����1 ����
����1.1 ����
������ij��ѧ��ļ 31 ����ѧ��������о�, ����������䵽������(15 ��, ���� 6 ��, Ů�� 9 ��; ƽ������(21.63±1.54)��( x ±SD, ��ͬ))���з�����(16 ��,���� 8 ��, Ů�� 8 ��, ƽ������(20.94±1.24)��). ���б��Ծ�Ϊ������, û���κ���������������ʷ, �����������������. �ڻ�֪����ʵ����̵ĸ���ϸ�ں�, ÿ��������Ըǩ����֪��ͬ����.
����1.2 ʵ������
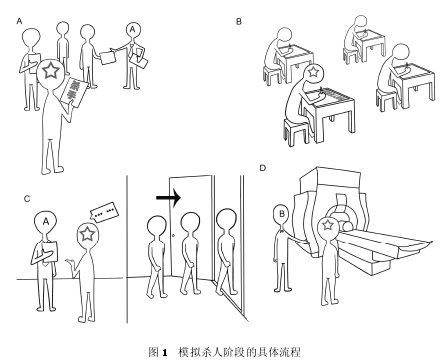
����(1) ģ��ɱ�˽�. ģ��ɱ�˽εľ���������ͼ 1 ��ʾ. ����, ʵ���� A(������)��֯ 1 �����Ժ����� 3 ���ٱ���һ���ǩ, ��ȷ��������ɱ����Ϸ�е�������"ɱ��"����"����"(ͼ 1A). �����˸�֮,���� 4���н����� 1��"ɱ��"������ 3��Ϊ"����".��ʵ���е�ǩ����"ɱ��", �ɴ�ȷ���汻�Գ鵽������"ɱ��"ǩ, ����Ϊ��Ϸ�е�ɱ��. �����˻�����,���� 4 �˵����ݳ����Լ���������֪������, ���� 3λ�����ߺ������ʵ���߶���֪��, ÿ�������߶���Ҫ���Ⱪ¶�Լ�������. ����ȷ���Ժ�, 4 �������߶�Ҫ���һ���ʾ�(ͼ 1B). "ɱ��"(�汻��)��Ҫ���һ��ģ��ɱ���ʾ�(����渽¼1), ���ʾ����� 7������: ����Ҫ����д�� 2 �����ѵ�����, Ȼ��ѡ������һ����Ϊ"ɱ��"����, ������ѡ������ʱ�䡢���������������岿λ��װʬ��ڴ�����ɫ���Լ���ʬ�ص�. �������ѵ�������Ҫ����������д����, ����������ֻ��Ҫ���Դ� 2 ��������ѡ���н���ѡ��(��ѡ���� 6 ��ѡ�����"ɱ��"���̵�ɱ��ϸ��, ��Щѡ���Ϊ GKT ��ʽ�е�"̽��̼�"���������IJ�ѳ�����). ��"����"ֻ��Ҫ��дһ�ż�¼���˻�����Ϣ���ʾ�. ��д�ʾ�ʱ, 4 �������ߵ���λ�䱣���㹻����, ʹ���������������������ߵ��ʾ�����. ����ʾ��Ժ�, 4 �������߱���ɢ����ͬ����. ����������"ɱ��"�ķ���, Ҫ��"ɱ��"ʹ��֮ǰȷ���� 6 ��ɱ��ϸ�ڱ���һ��������������ɱ�˹���, ��Ҫ��"ɱ��"�������ƺ�ϸ����һɱ�˹���,ʹ֮�������������ݾ���, ֱ������ӡ�����(���Էdz������ظ� 6 ��ɱ��ϸ��)Ϊֹ(ͼ 1C). Ȼ�������˸��豻����ϸ��ָ����, ָ��������ڲ�ѹ����н��а�����Ӧ(����渽¼ 2). ���ñ�����һ�α��� 6 ��ɱ��ϸ��. ���, �����˽����Դ���˴�ɨ����, ����һλʵ���� B(�����)������в��(ͼ 1D).
����(2) ��ѽ�. ��ѳ���ʹ�� GKT ��ʽ, ����Ĵ̼����źͷ�Ӧ��ʽ�����������о�һ��[13,25~27],���� 3 ��̼�: (��) ̽��̼�(probe, P). ����̼���ģ��ɱ�˽�ѡ���ɱ��ϸ�����, ÿ�����Ը�����ͬ, �������� 6 ������: ѡ��ɱ������������(��"����")������ʱ��(��"��ҹ")������(��"��ͷ")���������岿λ(��"����")��װʬ��ڴ�(��"�ڿڴ�")�Ͳ�ʬ�ص�(��"�¹�"). ������Ҫ��װû������Щ�̼�,��"û����"�ķ�Ӧ, ����ƭ��Ӧ; (��) �ش̼�(irrelevant, I). ����̼���ģ��ɱ�˹�����, ��ʵ��������ָ��, ��ÿ�����Զ���ͬ. �����ڴ�����������̽��̼�ƥ��, 1��̽��̼�(ɱ��ϸ��)����4���ش̼�(����渽¼ 3 �� 4). ��Щ�ش̼�����ɱ�ֺ�������˵����֮ǰû�м������´�. ����ֻ��Ҫ����ʵ�����������з�Ӧ, ��"û����"�ķ�Ӧ, ����ʵ��Ӧ; (��) Ŀ��̼�(target, T): ����̼���ģ��ɱ�˹���Ҳû���κι�ϵ, Ҳ��ʵ��������ָ��, ��ÿ�����Զ���ͬ, �ڴ���������Ҳ��̽��̼�ƥ��, 1 ��̽��̼����� 1 ��Ŀ��̼�(����渽¼3). ����, ������Ҫ�ڲ��ǰ������Щ����, Ȼ����A: ʵ���� A ������Ϸ������, ����DZ�ǵ����汻��, ���� 3 ��Ϊ�ٱ���, ��ǩ����"ɱ��"����; B: "ɱ��"���ģ��ɱ���ʾ�; C: 4 ����������ɢ����ͬ����, ɱ�ָ���ģ��ɱ���ʾ���ȷ�����װ�ϸ�ڱ������; D: ����һ��ʵ���� B ��������߲�ʹ�ú˴Ź���Ա��Խ��в�Ѳ���а���ʵ�����������"����"�ķ�Ӧ, ����ʵ��Ӧ. ���ڶ�Ŀ��̼��ķ�Ӧ("����")��ͬ����������̼�(��Ϊ"û����"), ��Ŀ��̼������ɢ�������̼��������ά�ֱ���ע����������[13,25,26]. GKT��ʽ�IJ��ԭ����: ���б���(����ɱ�ֺ�����)����̽��̼����ش̼�������ͬ����Ϊ��Ӧ(�ش�"û����"), ��ֻ��ɱ�ֿ���ʶ��̽��̼�, �������Ƕ���, ̽��̼���С���ʵġ�������Ĵ̼�, ������"û����"��Ӧ��Ҫ�漰��ƭ����, ���շ����صĴ��Լ���ģʽ; ������������̽��̼����ش̼�, ����̼���������ȫ��ͬ�Ĵ��Է�Ӧ[26]. ���,ͨ���۲챻�Զ�̽��̼����ش̼��Ĵ��Է�Ӧ�Ƿ���ڲ���, ��ɵ�֪�����Ƿ�����, �ж������Ƿ�"����".
�����ڴ̼�������, ������ֻ�з�Ӧ��, Ҫ���Զ�ÿ������ͨ��������Ӧ�ش��Ƿ����, ���з����黹��������ķ�����: �ڶ�ÿ��������з�Ӧ��, ���Ի�õ����Բ����(�����)�ķ���. ������"+2"��" 2"����ʽ����, �ֱ�������ϻ��ȥ 2 ��.
��������߸��߱���, ���Ŀ��̼��ķ��������������Ƿ���ȷ��Ӧ������, �������ȷ�ش�"����"����"+2"(positive feedback following target, TP), ������ �� �� " û �� �� " �� �� �� " 2"(negative feedbackfollowing target, TN). ���ĵ÷ֽ���������Ƕ�Ŀ��̼��ļ�������Ͳ�ѹ����н��з�Ӧ������̶�. ��̽��̼����ش̼��ķ�����������߶��䷴Ӧ���ж����, "+2"��ʾ�����ǵķ�Ӧ�ж�Ϊ��ʵ,��" 2"���������ǵķ�Ӧ�ж�Ϊ��ƭ, ���ĵ÷�Խ��Խ���ܱ��ж�Ϊ"����", Խ��Խ���ܱ��ж�Ϊ"ɱ��". ʵ����, ����Ŀ��̼��ķ���ȷʵ�ǻ��ڱ����Ƿ���ȷ��Ӧ������, ��̽��̼����ش̼��ķ����������趨�õ�. ��ÿ�ִ̼�����, "+2"��" 2"���ֵĸ��ʸ�ռ 50%, �ɴ��γ� 4 �����͵��ж����: ��̽��̼�������ƭ��Ӧ������Ϊ��ʵ(̽��̼�������Է���(positive feedback followingprobe, PP))����ȷ�ж�Ϊ��ƭ(̽��̼���ĸ��Է���(negative feedback following probe, PN)); ���ش̼����г�ʵ��Ӧ����ȷ�ж�Ϊ��ʵ(�ش̼�������Է���(positive feedback following irrelevant, IP))������Ϊ��ƭ(�ش̼���ĸ��Է���(negativefeedback following irrelevant, IN)). ���ÿ��̼��ķ�Ӧ��ʽ������������μ�ͼ 2A.
�����̼�����ͨ�� E-prime 2.0 ����. �ޡ��з�����Ĵ̼����̼�ͼ 2B �� C. ÿ�� trial ��ʼʱ, ��Ļ�������һ������ 1 s �Ĵ���, ������һ������ 0.5 s �ķ�Ӧ��ʾ�ź�"*". Ҫ�����ڴ�����ֽ�ֻ����Ӧ��, ֱ����ʾ�źų��ֺ������Ӧ. ���Ե��������ж��Ƿ�����ô���. Ϊ��ƽ����ָ�İ���ЧӦ, ��ÿ�鱻����, 1/2 ����ͨ������Ĵָ��"1"���ش�"����", ����Ĵָ��"3"���ش�"û����", �� 1/2 ����ʹ���෴�İ���ģʽ. ����������, ��ʾ�ź���ʧ��, ���ֳ���ʱ���� 0.5/2.5/4.5 s �������ע�ͷ���"+", ��������һ�� trial; �����з�������, ��ʾ�ź���ʧ��, ����һ������ʱ���� 0.5/2.5/4.5 s ������ĺ���, ��֪���Ե��Դ�ʱ���ڷ������ǵĴ����ź�, ������ʧ���ֳ��� 1.5 s �ķ���, ���ų��ֳ���ʱ���� 0.5/2.5/4.5 s �������ע�ͷ���"+", ��������һ�� trial. ������Խ����˴���, ��ֱ�ӷ���" 2", ������Բ�����0.5 s��������Ӧ, ����"��Ӧ".
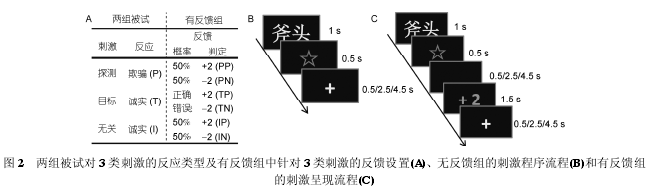
����ɨ��ǰ, �Դ�������ʱ���̽��̼�������Ӧ��Ŀ��̼����ش̼���Ϊʵ������ñ��Խ��г����ϰ, �ﵽ 90%���ϵ���ȷ�ʺ��ٽ�����ʽʵ��.��ʽʵ�����5��ʵ�����, ÿ������һ��̽��̼����м��(������ϰ���ϵ�"����ʱ��"��������), 5 ��ʵ�������ֵ��Ⱥ�˳���ڱ��Լ����.ÿ��ʵ�������� 1 ��̽����1 ��Ŀ������Լ�4 ���ش���. ̽������Ŀ�����ֱ��ظ����� 14��, 4 ���ش����е� 2 ������ֱ��ظ� 11 ��, ����2 ������ֱ��ظ� 10 ��, ���ÿ��ʵ�������� 14��̽�� trial, 14 ��Ŀ�� trial �� 42 ���� trial, ����70 �� trial. 70 �� trial ��ÿ��ʵ��������������.
����(3) �º����۱���. ɨ������Ժ�, ÿ��������Ҫ�ٴα���̽��̼���Ŀ��̼�, ��ȷ�����Ƕ�����̼��ļ������. ���, ������Ҫ���һ���º����۱����ʾ�, ���ʾ�ͨ��ѯ�ʱ��Զ��ӹ����(����������ɱ��)��ϣ���̶Ȳ������Բ��������"����ˮƽ"; ͨ��ѯ�ʱ��Կ���ÿ��̼�ʱ�Ľ��ų̶Ȳ������ԶԲ�ͬ�̼���"���۾���̶�"; ��ѯ���з����鱻�Զ�ÿ��̼�������"��ע�̶�". ����,���챻�ԶԱ��о���Ƶ������������µ��������������ʹ�����. ͨ���Ѽ����鱻�Ե���Щ��������, �˽���ƭ���������ƭ���̵���Ҫ��������, ��Ϊ fMRI ����Ľ����ṩָ��.
����1.3 fMRI ���ݲɼ�
�����Գ�������ʹ�������� 3T ȫ���Ź����������(Siemens Magnetom Trio Tim, �¹�)�Ѽ�, ���� 8 ͨ�������ͷ��Ȧ��ȡ���ԵĴ��ԴŹ����������.Ϊ�˷�ֹͷ��, ʹ�óĵ�̶�ͷ��, �����߱�����ʵ������о�������ͷ����ֹ. �Ӿ��̼�ͨ���Ź����������õ�רҵͶӰ�豸����, ����ͨ��ͷ����Ȧ�ϵĵ��Ӿ����йۿ�. Ϊ�˼���ɨ���Dz����������Ա���ִ��ʵ�������Ӱ��, ÿ���������ר�õķ���������.
����ȫ�Թ�������ö�Ѫ��ˮƽ����(blood oxygen-ation level dependent, BOLD)���������Ե� T2*��Ȩ���μ����ݶȻز��� EPI(echo planar imaging, EPI)���л��, �������ÿ�����Թ��ɼ� 880 �� TR ��ȫ�ԵĹ���Ӱ��, ���з������ÿ�����Թ��ɼ� 1360�� TR(repeat time)��ȫ�ԵĹ���Ӱ��. ÿ����������32 ��, ���ø���ɨ��ķ�ʽ, ��� 3 mm, ����1 mm, ɨ�跽��ƽ���� AC-PC ƽ��. ɨ���������:
����TR=2000 ms, TE(echo time)=30 ms, ɨ �� �� ҰFOV(field of vision)=220×220, ɨ�����(matrix)Ϊ64×64, ��ת��(flip angle)Ϊ 90°. ÿ�� trial ���ֵ�ʱ��ͨ�� E-prime ��������ɨ��ͬ��. ����, ʹ�����²�����ȡÿ�����ԵĽṹ������: TR=1900 ms,TE=2.52 ms, ��� 1 ms, ɨ����Ұ FOV=256×256, ��ת��Ϊ 90°, ����ÿ�����صĴ�СΪ 1×1×1 mm3.
����1.4 fMRI ���ݷ���
������ �� �� �� �� �� �� �� Ԥ �� �� �� �� �� �� ��SPM8(Wellcome Department of Cognitive Neurology,Ӣ��)���߰�, �ù��߰����� MatLab(Math works, ����) ƽ̨����. ��Ԥ������, ���Ȳ��� slice timing�������ʱ����, �ԶԸ�ɨ����ڻ�ȡʱ���ϵIJ�����н���, �ٽ���ͷ������(realigned). Ȼ��,��ͼ�����ݻ���SPM8�ṩ��EPIģ����пռ��������(normalize), ͬʱ�������������ز���Ϊ3×3×3 mm3�ֱ��ʴ�С. ���, ʹ�� 8 mm ��ƽ���˽��и�˹ƽ������(smooth). ���б��Ե�ͷ��С��3 mm.
������һˮƽ���������¼����ģʽ. ���ÿ�����Խ����̼�����, �����鱻�Զ���, ��������Ӧ�ε� 4 ������(regressors): ���Զ� 3 ��̼�(P, T �� I)������ȷ��Ӧ(��̽��̼�����ȷ��Ӧ��ָ���������ƭ��Ӧ)�� trial ���ɵ� 3 ������Ȥ regressor. ����Ӧ����Ӧ�� trail �ķ�Ӧ�ι����� 1 ������Ȥregressor(incorrect response, INR). �� 4 �� regressor �ľ���ʼʱ���Ӧ�ڴ̼�����ʱ��, duration ����Ϊ0. ���з��������, �������˶��� 7 �������ε�regressors, ����������ȷ��Ӧ����ֵ� 6 ���ж����(PP, PN, TP, TN, IP �� IN)���ɵ� 6 ������Ȥ regressor,�Լ��ɴ������Ӧtrial�ķ����ι��ɵ�1������Ȥ regressor(incorrect judgment, INJ). �� 7 �� regressor�ľ���ʼʱ���Ӧ�ڷ�������ʱ��, duration ����Ϊ 0. ����, ÿ�����Ե� 6 ��ͷ������Ҳ��Ϊ����Ȥregressor ������̼�����, ���ų�ͷ�����źŵ�Ӱ��.
����ʹ�� GLM(general linear model)�� HRF(hemodynamicresponse function)�������и���ˮƽ��ͳ�Ʒ���, ���� 128 s �ĸ�ͨ�˲�������Ƶ���������ݵ�Ӱ��. ���о���ע�С�����ƭ�������ƭ���̵ĵ�������, ���ֻ���췴Ӧ�εĴ��Լ���ģʽ, �������������ε�����. ����, ��������Ŀ��̼�(T)������ά�ֱ��ԶԴ̼���ע��[25], ���Ҳ����Ŀ��̼����з���. ʹ�õ�����t����, �ֱ�P(̽��/��ƭ)��I(��/��ʵ)�����������������, ���� P ������ I �������, ��� 3 �����(contrast)��� contrast �Ժ� beta��ͼ��, ��Щͼ���������ĵڶ�ˮƽ����.
�����ڶ�ˮƽ��������һˮƽ�����еõ��� P �� I������Ӧ�� contrast ��ͼ��(con �ļ�)����һ�� 2(��/�з���)×2(̽��/��)��ȫ����ģ��(full factorialanalysis). ��ЧӦ������� FWE(family wise errorrate)<0.001, ����ش��� 50 ������(K>50)����ֵУ��.
����Ϊ��̽���������صĽ�������, �ſ���ֵ�� P<0.005,K>10, ��Ȼδ��У��, �� P<0.005 ����ֵ����������ѧ������ձ��Ͽ�[28,29], �ڸ���ֵ�»�õĽ� �� �� �� �� �� �� �� �� �� �� �� . �� �� �� �� �� ��AAL(anatomical automatic labeling)ģ����ж�λ.
�������ڷ������ͷ�Ӧ����֮��Ľ���������˫����״�弰����Ե�������, ���ݽ������õĽ�������� 3 ��������Ȥ����(region of interest, ROI), Ȼ����ȡÿ�������� 3 �� ROI ��̽��̼����ش̼���Ӧ�ļ���ǿ�Ȳ���(parameter estimates, PEs), �ٽ������һ��2(��/�з���)×2(̽��/��)���ظ������������, �Թ۲콻�����õľ���ģʽ.
����2 ���
����2.1 �����
����(1) ��ȷ�ʺͷ�Ӧʱ. ���о�����Ŀ��̼���ά�ֱ��ԶԴ̼���ע��, �ޡ��з����鱻�Զ�Ŀ��̼��ķ�Ӧ��ȷ�ʶ��ϸ�( x �ֱ�Ϊ(89.9±0.111)%��(95.6±0.030)%), ˵�����鱻�Զ�����ִ���˰�������. ���鱻�Զ�̽��̼����ش̼��ķ�Ӧ��ȷ�ʺͷ�Ӧʱ���� 1. ���������ȷ�ʽ��� 2(��/�з���)×2(̽��/��)���ظ������������, P ֵ����Greenhouse Geiser ��У��. �������, �����ЧӦ������, F(1, 29)=1.855, P=0.184. �̼�������ЧӦ������, F(1, 29)=0.000, P=0.989. ������֮��Ľ�������Ҳ������, F(1, 29)=0.128, P=0.723. �Է�Ӧʱ����ͬ�����ظ������������, �������, �����ЧӦ������, F(1, 29)=2.548, P=0.121. �̼�������ЧӦ������,F(1, 29)=1.514, P=0.228. ���ʹ̼����������ؼ�Ľ�������Ҳ������, F(1, 29)=1.259, P=0.271. ��Щ���˵��, ����̽��̼����ش̼���Ҫ��ȫ��ͬ�İ�����Ӧ, �����շ�����ͬ����Ϊ��Ӧģʽ.
����(2) �º����۱���. ɨ�������, ���б��Զ���ȷ����ر���Ŀ��̼���̽��̼�, ˵��������ʵ�������, ���Զ�����̼��ļ�������ȷ���ι̵�.�º����۱���Ľ����� 2 ��ʾ. �������, ���鱻��ϣ��ͨ����ѵĶ���ˮƽ���ϸ�, ������ 4(5������), �Ҷ������� t ����Ľ������, ����Ķ���ˮƽû�����Ų���, t(29)=1.158, P=0.256. ��˵�����о����õ�ģ��ɱ���龳�շ������б��ԽϸߵĶ���ˮƽ.
����ʹ�ñ��ԶԴ̼������۾���̶Ƚ��� 2(��/�з���)×2(̽��/��)���ظ������������, P ֵ����Greenhouse Geiser ��У��. �������, �����ЧӦ����, F(1, 29)=18.081, P=0.000, �����������, �з���������д̼������۾���̶ȶ�����. �̼�������ЧӦ����, F(1, 29)=32.592, P=0.000, ̽��̼����ش̼������˸��ߵ����۾���̶�. �����ؼ�Ľ�����������, F(1, 29)=6.083, P=0.020, ��ЧӦ�����Ľ������, ��������, ̽��̼������ش̼������˸��ߵ����۾���̶�(�з����� P=0.000, ������ P=0.027), ����, �ش̼����з��������շ������۾���̶�����ǿ��������(P=0.000), ��̽��̼����������շ������۾���̶�û�����Ų���(P=0.602).
����ʹ���з����鱻�Զ�̽��̼����ش̼������Ĺ�ע�̶Ƚ������ t ����Ľ������, ���Զ�̽��̼���ķ����ȶ��ش̼���ķ�����Ϊ��ע,t(15)=4.869, P=0.000, ˵�����Բ�����������̽��̼����ش̼�, �������Ǹ���ע��̽��̼�������ƭ��Ӧ�Ľ��.����, ���б��Զ�������������ʵ������ʹ�õ�"�������"ȷʵ�߱������Ե�����. ���˱���"����ɱ��ϸ�ڴ���ʱ���������侲"��һ�ձ������, û�б��Ա�������ʹ�����κ��������������.
����2.2 fMRI ���
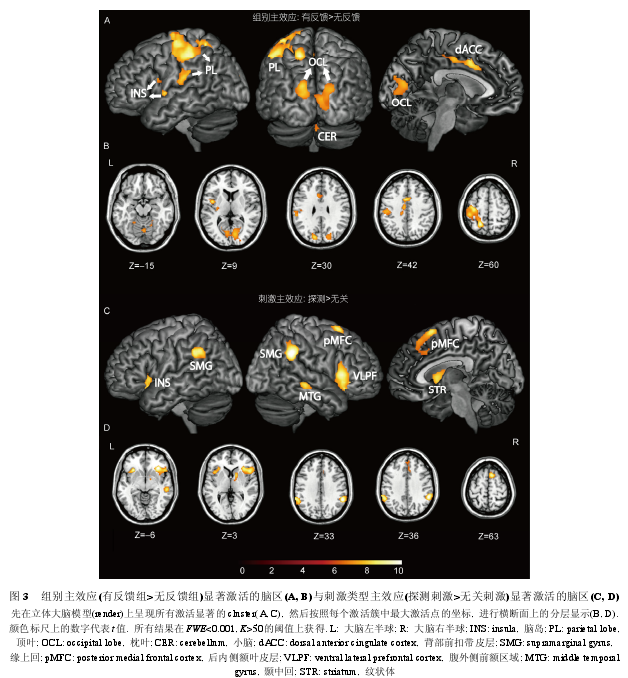
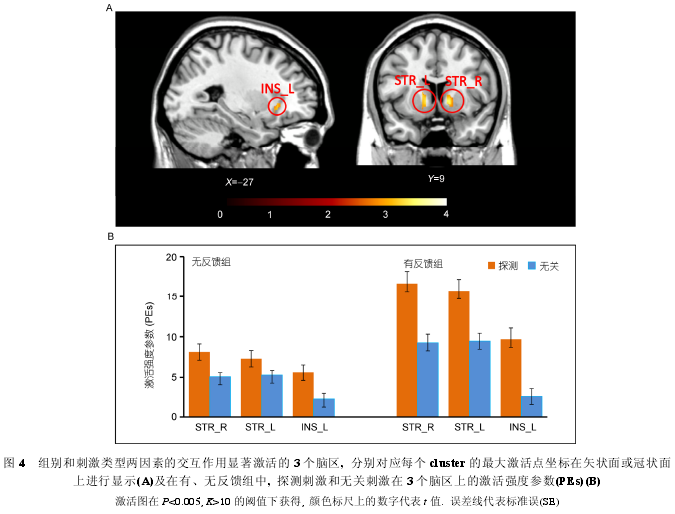
����ȫ����ģ�ͷ����Ľ�����3��ʾ. �����ЧӦ����, �ڷ�Ӧ��, �з�����������鵼���˸�������ǰ������������ඥҶƤ�㡢��� dACC����� INS �Լ�˫���Ӿ�Ƥ��Ĺ㷺������Ҳ�С�Եĸ���(ͼ 3A); ���������֮�з�����û�е����κ������ĸ���. �̼�������ЧӦ����, ̽��̼����ش̼��������Ҳ����Ƥ��(λ�ڸ����ǰ������, ventral lateral prefrontal cortex, VLPF)��˫��Ե�ϻ�(supramarginal gyrus, SMG)(BA40)����� INS���Һ��ڲ��ҶƤ��(posterior medial frontal cortex,pMFC)���Ҳ���л�(middle temporal gyrus, MTG)�Լ��Ҳ� STR �ĸ���(ͼ 3B); ���ش̼���֮̽��̼�û�е����κ������ĸ���. �ڸ����ɵ���ֵ��(P<0.005, K>10)����, ���ʹ̼������������صĽ���ЧӦ(�з�����(̽��>��)>������(̽��>��))��˫��STR�����INS������(ͼ4A��B).
������Խ���ЧӦ���ŵ� 3 �����������幦�� ROI, ��ʹ���� 3 �� ROI �ļ���ǿ�Ȳ���(PEs)�ֱ���� 2(��/�з���)×2(̽��/��)���ظ�������������Ľ����ʾ, 3 ��������ȷʵ�������ŵĽ���ЧӦ(��� INS:F(1, 29)=8.05, P<0.01; ��� STR: F(1, 29)=8.66,P<0.01; �Ҳ� STR: F(1, 29)=9.28, P<0.01); ��һ����ЧӦ�����Ľ������, (��) ���С�����������,̽��̼��� 3 �� ROI �ϵļ��ǿ���ش̼�(������������, ̽��̼����ش̼������ STR �ϵļ�������Ե����, P=0.067, ����Ϊ P<0.01); (��)��̽�����������, 3 �� ROI ���з������еļ��ǿ��������(����� INS ���ش̼��ļ���ǿ�����С��������û�в���, P=0.81, ����Ϊ P<0.05);����, ̽��̼����ش̼�����Щ�����ϵļ���������з������б����������и�������. Ҳ����˵, ���з�����, ��̽��̼����з�Ӧʱ�������ƭ���̸���̶ȵ��漰���� 3 �������IJ���.
����3 ����
�������о���һ��ģ��ɱ���龳��, �����鱻�Էֱ�ʹ���ޡ��з���(��ƭ���)�� GKT ��ʽ���в��,ͨ���Ƚ����鱻�Խ��г�ʵ��Ӧ����ƭ��Ӧʱ�Ĵ��Լ���ģʽ����, ���������ص�����������Ӧ��(������ʵ��Ӧ����ƭ��Ӧ)������; ����ƭ�������漰���ڶ�������, ������Ԥ�ںͷ����������̵���״����Ե���������ܵ����������صĵ�������.
����3.1 ����������������Ӧ�ε�����
����fMRI ����е������ЧӦ(�з�����>������)�����˷�����������Ӧ�εĵ�������. �����ЧӦ��ʾ, �з�������������ڷ�Ӧ�ε����˸�������ǰ����ص���ඥҶƤ�㡢��� dACC�����INS��˫���Ӿ�Ƥ�㡢�Լ��Ҳ�С�Եĸ���, ˵�������Է�Ӧ�β����˹㷺��Ӱ��, ��һӰ��ͬʱ�漰��ʵ��Ӧ����ƭ��Ӧ, ʹ�����ַ�Ӧ���շ��˸�Ϊ���ӵ���֪����. ����, dACC ��Ϊ��������ϵͳ�ĺ����������д�������[30~32]. dACC ������֪�Ե�������������[30,33], ����������������״̬�Ĵ�����������[31]. �ڴ�������������, ������Ҫ�����˵�֪ʶ״������, �������������ʵ��������, ���ջ�ö���������ľ�֪������. Ȼ��, ������������������һ����֪������(perception-based)[34~36], ��ֻ�е����˵�����ͨ��һ��������������¶���������������֪��ʱ, ����ſ���������������������״̬������. �ڱ��о���, ������䵱����һ֪�������Ľ�ɫ: ���з�������, ���۱�������ʵ��Ӧ������ƭ��Ӧ, ��Ӧ���ò���߶���һ��Ӧ���ж����, ������һ�ж�, ���Ե������۲���߹����Լ�����(ɱ��/����)�ĵ�ǰ����, �����ڶԲ���ߵ�ǰ�������������������ķ�Ӧ����, ����һϵ�з����ƽ����������ۼӹ�����, �㷺������������Ӧ�κͽ���ӹ���; Ȼ��, ����������, ����������йز���ߵ�ǰ������κ�������Ϣ, �����������ۼӹ���������ֻ�ܺ�С�̶ȵĽ���. ���, ���鱻�Զ��������ۼӹ����̵���֪����̶��������Բ����, ��һ�������������� dACC �ļ��������.
�����з�����������黹�����˸�������ǰ��صĶ�Ҷ����С�ԡ��Լ��Ӿ�Ƥ��ĸ���. ����ǰ��غ�С�����������˶�Ƥ��, ���ǵĸ������˵���з����龳�µķ�Ӧ�����շ��˸���̶ȵ���֪����[37,38]. ������ע��ϵͳ�Ķ�ҶƤ�㡢�Ӿ�Ƥ���²�����ص��������з����龳�µĸ�����ܷ�ӳ�����۾����ע����̵���ǿ����[39].
����3.2 ������������ƭ���̵�����
����fMRI ����еĴ̼�������ЧӦ(̽��>��)��ӳ����ƭ�������漰�Ĵ��Լ���ģʽ. ��������, ̽��̼����ش̼��������Ҳ� VLPF��˫�� SMG�����INS���Ҳ� pMFC���Ҳ� MTG �Լ��Ҳ� STR �ĸ���, ��Щ������������ƭ�о��ж��ѻ�ù㷺�ı���[3,18,20,25,40~43], ���о��ķ����������о�����һ��, ֤�����о���ʵ�������Ч���շ�����ƭ��Ϊ.
����fMRI ����е����ʹ̼����͵Ľ���ЧӦ(�з���(̽��>��)>����(̽��>��))�����˷���(��ƭ���)����ƭ���̵ĵ�������. ����ЧӦ��˫��STR ����� INS 3 ������������, �����Ҳ� STR ����� INS �������̼���ЧӦ����е��Ҳ� STR �����INS �ص�(peak �������<3 mm), ����, ��������ֵ�� FWE<0.005 ʱ, �̼���ЧӦ����� STR ��Ҳ����������(����渽¼ 5), ��ζ���� 3 ����������ƭ����. ��һ����ЧӦ�����Ľ��֤ʵ����һ���۹۲�: ��������, 3 ������ȷʵ�����ֳ����ŵĴ̼�����ЧӦ, ����̽��̼��ļ��ǿ�ڶ��ش̼��ļ���.
��������, ��ЧӦ�������������, ��̽��̼����ش̼�������, 3 ���������з������еļ��ǿ��������, ˵�������ձ���ǿ����Щ��������ƭ��Ӧ�ͳ�ʵ��Ӧ�еļ���ˮƽ; ��Ϊ��Ҫ����, ̽��̼����ش̼��� 3 �������ϵļ���������з������б����������и�������, ˵���з����龳�µ���ƭ���̸���̶ȵ��漰����Щ����IJ���. ���д����о�֤ʵINS�������������[44,45], ����״����뽱��Ԥ�ڹ���[46~50], ������ STR ��Ԥ�ڽ��͵����ۼ�ֵ����[50~52]. ������Ϸ���, ���о���Ϊ,(��) ����������ƭ���ʱ(������), ��ƭ�Ѿ��漰һ���̶ȵķ������ͽ���Ԥ�ڹ���, �����ʵ���, ��ƭ����Ϊ���߷���, ���ͬʱ, ���ʵ��������ȷ�ж�Ϊ��ʵ(����ԩ��)��������, ��ƭ��ȴ����������Ϊ��ʵ(�ɹ���ƭ)��һ����Ե�Ԥ�ڽ��Ͷ���ƭ�߾��и�������ۼ�ֵ; (��) ����Ӧ��Ҫ���ټ��̵��ж����ʱ(�з�����), ���ԶԷ�Ӧ���(����)�ĵ��Ǻ��ڴ���ǿ, ��˳�ʵ��Ӧ����ƭ��Ӧ���漰�˸�ǿ�ķ��������ͽ���Ԥ�ڹ���; ����, ��Щ�������̵���ǿ������б����ƭ��Ӧ, ����˵����Ҫ�����ƭ������з����鱻������ƭʱ�ر��ı�����, ͬʱ���ر�ϣ���ӹ�����������⽱��, ��Щ����ʹ������(�������鱻�����)����ƭ�����в�������ǿ�ҵķ���������鲢�Գɹ���ƭ�Ľ���(����ֻ������ԵĽ���)�����˸��ߵ����ۼ�ֵ. �����з����鱻�Ե����۱�������һ���̶���ӡ֤����������, ���۱���Ľ������, �з����鱻�Զ�̽��̼�����ƭ��Ӧ�ܷ�ɹ��Ĺ�ע�̶�Ҫ���Ÿ��ڶ��ش̼��ij�ʵ��Ӧ�ܷ���ȷ�ж�Ϊ��ʵ(����ԩ��)�Ĺ�ע�̶�, ˵����ƭ��ķ���(��ƭ���)�Ա��ԵĶ�����Ҫ��, ��˲����������Щ�����Ԥ�ڼӹ�Ҳ������ǿ��.
������INS��STR��, ��ƭ�����漰����������, ��˫�� SMG���Ҳ� VLPF���Ҳ� MTG ���Ҳ� pMFC ��û�г������ʹ̼����͵Ľ���ЧӦ, ˵�����Ƕ���ƭ�IJ���û���ܵ�������(��ƭ���)�ĵ���. �����о���Ϊ, ��Щ��������ƭ�в���ִ�п��ƹ���[1,20], ���Ǹ������Ƴ�ʵ��Ӧ����, ��������ƭ��Ӧ�IJ���[20]. ���������о��ͱ��о����, �ƲⷴӦ���ƹ����ڲ�ͬ���͵���ƭ�ж�����Ϊ��������֪�ɷ�, ������������ܵ���ƭ�����������龳���ص�Ӱ��. �෴, INS �� STR ������ķ��������ͽ���Ԥ�ڹ�������ƭ�е������֪����гɷ�, ���ǽ������ܵ�����龳��Ӱ��, �����ܵ���ƭ����ĵ���.
����4 ����
�������о�ʹ���ޡ��з���(��ƭ���)�� GKT ��ʽ,���ڴ����ƵĽǶȿ�����ƭ�������ƭ���̵ĵ�������. �о��������, �����������, �з����鱻������ƭ��Ӧ�ͳ�ʵ��Ӧ�϶�����̶ȵؾ������������ۼӹ�����, ��ӳ�� dACC �ĸ�����;���ҷ���, �з����鱻�Ա������鱻������ƭ�����и���̶ȵؾ����˽���Ԥ�ںͷ�����������,��ӳ������������������������ص����� STR ��INS �ĸ�����. ��ƭ�������ƭ�����ƾ������Եĵ�������, ����Ҫ����ƭ�������֪����гɷֲ���Ӱ��.
����5 ����IJ���
�������о����õ� GKT ��ʽ��һ��"ָ����"����ƭ��ʽ, ��Ҫ������ʵ���ߵ�ָ���¶��ض��Ĵ̼�������ƭ��Ӧ, ������ʵ�и����Է�����ƭ��Ϊ��һ�����. ָ������ƭ��ʽ���ھ������ڿ��Ʊ����������Ʋ�ͬʵ���������źŵ��Ӵ������ŵ�, ������̽����ƭ�е��±���ЧӦ���۽��������̽�����о�, �����Ȼ���������ƭ�Ͳ���о���ʹ��[6,7,10,11,18,25,41,42,53~58]. ����, ָ������ƭ��ʽ�Ͼ�ȱ����̬ѧЧ��, �������о��б�Ҫ�����Է���ƭ��ʽ, �罫ָ�����Ϊ"�����ӱܲ��", Ȼ���ñ����Է���ѡ���ʱ������Щ�̼�����, �ڸ�Ϊ��ʵ����ƭ��Ϊ�п�����ƭ�������ƭ���̵ĵ�������.
���������
����1 Abe N. How the brain shapes deception. Neuroscientist, 2011, 17: 560–574
����2 Ford E B. Lie detection: historical, neuropsychiatric and legal dimensions. Int J Law Psychiat, 2006, 29: 159–177
����3 Langleben D D, Schroeder L, Maldjian J A, et al. Brain activity during simulated deception: an event-related functional magneticresonance study. NeuroImage, 2002, 15: 727–732
����4 Spence S A, Farrow T F D, Herford A E, et al. Behavioural and functional anatomical correlates of deception in humans. Neuroreport,2001, 12: 2849–2853
����5 Lee T M C, Liu H L, Tan L H, et al. Lie detection by functional magnetic resonance imaging. Hum Brain Mapp, 2002, 15: 157–164
����6 Ganis G, Morris R R, Kosslyn S M. Neural processes underlying self-and other-related lies: an individual difference approach using fMRI.Soc Neurosci, 2009, 4: 539–553
����7 Abe N, Suzuki M, Tsukiura T, et al. Dissociable roles of prefrontal and anterior cingulate cortices in deception. Cereb Cortex, 2006, 16:192–199
����8 Abe N, Suzuki M, Mori E, et al. Deceiving others: distinct neural responses of the prefrontal cortex and amygdala in simple fabricationand deception with social interactions. J Cogn Neurosci, 2007, 1

�����ش���ظ�ԭ���أ�ƽ��������4000m���ϣ�2010��4��14�յ���Ե��ػ�����ʩ����˾���ƻ�������ڳ����ؽ��Σ�ȫ������½���д���ʩ����Ա��������������ǿ�����Ͷ����д���Ϊƽԭ�˳�����ԭ��������ϰ��ʷ���ʽ����ԭ�������׳���һϵ...
�����ڸ���֬�ʴ�л�Ĺ���������Ҫ�ĵ������á���ʳ��������ҩ������ؿ�ͨ����������;�����ڸ���֬�ʴ�л����Щ����ʹ֬�ʴ�л���ң����¼�������;��Щ����ʹ���ҵ�֬�ʴ�л�ָ�ƽ�⡣...
���о�ͨ��ȫϸ��ĤƬǯ�����ķ�����������ϸ������������Ĥ���Ժ�ͻ���ȶ��Եľ���仯�ص�, �˽����ֵ��������Ըı���������ڽ�һ������RGC���Ӿ���Ϣ�з��ӵ����á�...