����ѧ����

����ժ Ҫ�����Ӿ���������ָ����ά�ֲ�����������Ϣ�����л�Ծ��֪�ӹ��Ĺ���, ���������֪��о��м�����Ҫ������.����, Ŀǰ��������������Ӿ���������ά�ֽζԶ��������ӿ��� (����, �����Ϣ�ͷ�����Ϣ) ��Ϣ����֪�ӹ����ƻ�û�еõ�������о�.����, �Բ�ͬ�Ŀ�����Ϣ����ά���Ƿ��Ӧ��ͬ����֪�ӹ�ģʽ, �Լ��Ӿ���������ά�ֽθ���ЧӦ��������û�еõ��㷺���о�.���, ���о��������Ϣ�ͷ�����Ϣ���Ӿ����������������, ������Ϊʵ����ܺ˴Ź�����̽���Ӿ���������ά�ֽ��жԲ�ͬ�������ݵ���Ϊ���֡����Լ���ǿ�ȡ��Ӿ�Ƥ�������Ƽ��为��ЧӦ, Ϊ���Թ��ܷ�������Ӿ������������ݵı����ṩ���м�ֵ��Ӱ��ѧ����.�о��������, �����ڶԸ��ӿ�����Ӿ��������������б��ֳ��Ӿ��������������������Ժ���ЧӦ, ����ͨ��˫�ຣ���ԡ�˫�ඥ�ڹ��ļ���ǿ�Ⱥ�V4��˫����״�ء�˫����Ϲ�����Ϣ������ȷ�������Ӿ���������ά������.����, ˫��Ǻ˵ļ���ǿ�����������Ӿ��������为��ЧӦ.
�����ؼ��ʣ����Ӿ���������; ά�ֽ�; ����ЧӦ; ��Ϊ����; ����ǿ��; �������;

����Abstract����Visual working memory (WM) refers to the maintenance and manipulation of visual information that is not in the visual field. Visual WM provides an essential connection between perception and higher cognitive functions. Previous f MRI studies with classification methods showed that early visual cortex could retain information of visual features maintained in WM. However, we still know little about the neural mechanisms of visual WM during maintenance period in higher visual cortex and the influence of memory load on such mechanisms. Therefore, the present study utilized a modified delayed match-to-sample paradigm to investigate the visual WM performance. Participants were required to maintain one or two target items (face or house) for 11 s. At the same time, function magnetic resonance imaging (f MRI) was applied to acquire the neural activities in visual cortical areas. Behavioral results showed that memory load but not stimuli significantly affected the accuracy of memory. Next, the amplitude of the neural activity during maintenance in each area was extracted. In order to reveal the areas sensitive to mnemonic stimuli, we compared the activity induced by face and house WM. The results showed that parahippocampal place area (PPA) and intra-parietal sulcus (IPS) were activated differently to different stimuli. Specifically, the neural response was stronger when house relative to face was held in memory. To further investigate whether other areas also retained visual information in WM, activation pattern analysis with decoding method was applied to the neural response in each area. The results demonstrated decoding performance in face selective areas including superior temporal sulcus (STS) , FFA and V4. Taken together, these results indicated that higher visual cortex could retain visual information held in WM. Specifically, PPA and IPS showed general stronger responses to mnemonic houses than mnemonic faces, while FFA, STS, and V4 contained different detailed representations of mnemonic faces and mnemonic houses. These findings were consistent with previous findings concerning visual features, and supported the view that the contents in visual working memory are represented in visual cortex. Notably, we did not find any effects of memory load on the neural activity in PPA and IPS. However, the activity pattern analysis about other areas showed significant effects only in high memory load condition, suggesting that high memory load might refine the representations in face selective areas without boosting the average activation level. In order to find the cortical areas of which responses were significantly modulated by memory load, we further performed a voxel-based whole brain analysis. Specifically, a 2 (stimuli: face, house) * 2 (memory load: 1 item, 2 items) repeated measures ANOVA was performed for each voxel on the neural response during the maintenance. No interaction was observed in each voxel. Interestingly, the main effect for memory load was significant in bilateral putamen. In summary, our findings revealed the crucial role of higher visual cortex in maintaining visual information in WM and the important role of putamen in modulating memory load.
����Keyword����visual working memory; maintenance phase; load effect; behavioral performance; activation intensity; decoding mechanism;
�����Ӿ��������� (visual working memory, v WM) ���ڹ��������һ��, ������ָ����������Χ�ڵ��Ӿ���Ϣ���ж��ݻ�Ծ�ӹ�����֪����[1~3].����ʵ������, �����ڽ��и�����֪���ʱ����Ҫ����ǰ���Ӿ���Ϣ���ж��ݵĴ洢�Խ������ǰ������ص���֪����.�����о�����, �Ӿ���������Ը�����Ϊ��������Ҫ��Ӱ��.�������Ӿ������������������������ע����ơ�����������ѧҵ�ɼ��ȷ���IJ���������[4~7].ͬʱ, �Ӿ�������������������֪���ӹ�����֪����֮��Ĺؼ�����[8~10].����, �Ӿ������������������֪������а����ż�����Ҫ������[11,12].�Ӿ���������ά�ֽε���֪�����о�Ҳ�������Ӿ����������о��漰�Ϲ㷽��֮һ, ͨ����ά�ֽν�����Ƶ��о�, �������ָ��Ԥ�������ά�ֽ���ά�ֵľ�������, ��ͬʱ̽���ض������ڽ��н���ʱ�ļ���ģʽ, Ϊ�������˽��Ӿ�����������֪ģʽ�Լ����������֪���̾��зdz���Ҫ������.
��������ʮ����, �о����Ƕ����Ӿ���������������о���ȡ����һ���ijɹ�, �Լ�������������ЧӦ����������Լ��洢���Ƶȷ������˸�������˽�[3,13,14,15].�ڹ����Ӿ��������为�ɵ��о���, �о������������һ��:��֪�ӹ�����Խ��, ���ԵĹ�������������ɼ�Խ��[14,16].һЩ�о��߹����Ӿ���������ά�ֽν�������о�̽���˳����Ӿ�Ƥ��ļ���ģʽ, ��ҪĿ����̽��V1~V4��Щ�����Ƿ����ض����� (content-specific) ��Ϣ, ��Ϊ�����Ӿ�Ƥ��ļ���ģʽ�������ֳ�����Ϣ[17,18].ǰ�����Ӿ���������ά�ֽε��о��еó�, ǰ��ҶƤ�� (prefrontal cortex) �����Ƥ�� (inferior temporal cortex) ����Ҷ-��Ҷ�������� (fronto-parietal areas) �������ļ���������Ӿ��������������жԿ�����Ϣ��ά��, ����, �ڲ�ͬ���о����漰�Ĵ̼������������쵼��ά�ֽμ����������ܻ���������[19].һЩ�о���, ���̼�����Ϊ����߷��ӵ�ʱ��, ά�ֽεĴ��Լ�����ܻ���Ҫ���������ѡ���� (��״��:FFA) �ͷ���ѡ���� (�����Ի�:PPA) [20,21].�ڶԵ���������ķ���������, ���о�����̽�������Ӿ�Ƥ���Լ�����Ƥ����ڶ��������� (����, ��ͷ��ӵ���Ϣ) �ļ���ģʽ, ���о��������, �����Ӿ�Ƥ��Ϳ���ѡ�������ԶԲ�ͬ������Ӿ���Ϣ��������[22].����Ҳ�й��ڳ����Ӿ�Ƥ����о�����, �����Ӿ�Ƥ����Ҫ�Ƕ��Ӿ���Ϣ����֪������ļӹ�Ȼ����Ϊ��Ϣ����;�����Ӿ���Ϣ���ݵ������������н�һ������֪�ӹ�����[23].
������������, ĿǰΪֹ, �����Ӿ�����Ƥ���ܷ�Զ��������ӿ�����Ϣ���н��뻹û�еõ�������о�, ��������������Ӿ���������ά�ֽζԶ��������ӿ��� (����, �����Ϣ�ͷ�����Ϣ) ��Ϣ����֪�ӹ����ƺ�ģʽҲû�еõ�������о�, ά�ֽθ���ЧӦ������ͬ����û�еõ��㷺���о�, ����, ���о����Զ���������Ϊ�̼�����̽�����Ӿ���������ά�ֽθ���Ƥ��Ľ������.��ʵ���Ŀ��Ϊ̽���ڶԾ��и�����������Ŀ���м����ʱ��, ���ԵĹ�������ɼ����ֵĸ���ЧӦ�Ƿ���̼������������, ���Եļ��������Լ�ͨ����Щ��������ļ����Ƿ����Ԥ�ⱻ����ά�ֵļ�������, �����Ƿ����ͨ���Ӿ�Ƥ��ļ���ģʽ�������Ӿ���������ά�ֽε��Ӿ������������, �����/�����Ӿ�����������, ����ЧӦ�ֱ��ڷ��Ӻ����������������ֵ�.
���������ڸ��ӿ�����Ϣ���Ӿ���������������Ҳ�������Ӿ������������������Ժ���ЧӦ, ���Ҳ����ܵ��̼����������Ӱ��.�����/�����Ӿ�����������, ǰ���о����Ѿ������漰�IJ�������, ������Ӿ�Ƥ�㡢���ѡ���� (FFA) ������ѡ���� (PPA) ��˫����Ϲ���˫�ඥ�ڹ���, ���������ڹ�������ά�ֽεļ���ǿ�Ȼ�ģʽ�ܹ�Ԥ�ⱻ����ά�ֵļ�������, ����Ԥ�����Ч�Ի����ż��为�ɵIJ�ͬ���仯.
�����仯���췶ʽ (change detection paradigm) [2]Ϊ�Ӿ����������о��춨�˼�ʵ�Ļ���.�ں��������Ӿ�����������о���, �����ǵ�������Ϊʵ���о����ǽ���Ϊʵ��ͬ���ܺ˴Ź�����������ѧ�о��������ϵ��о�����, �����漰����Ϊ��ʽ�����϶��ǽ����ڱ仯���췶ʽ����֮��, ������ʵ��Ŀ�����������ı�.�Ӿ���������ά�ֽ�ʱ�䳤�̵�ѡ��Ҳ���ݲ�ͬ��ʵ��Ŀ�Ķ�������ͬ.����, Ϊ��̽���Ӿ���Ϣά�ֹ����еĴ��Ի���, ��Щ����, ���о�����Ϣά��ʱ���ǰ���о��ļ��ٺ������ӵ�15 s����, ��ͨ����������ļ���֤��������Ӿ���Ϣ��ά��������ʱ���������Ч��[17].���ǵ��о����������ǵ�������ʾ���䷶ʽ, ֻ�Ǵ̼���Ϊ����ͷ��ݴ̼�.������ʾ���䷶ʽ�����������ʽ�ڹ�������ĴŹ����о��о��ж��ص�����.�÷�ʽ���Ա�֤�ڼ��䲻ͬ����ʱ����ͼ���ά�ֽ�ʵ�������̼���һ����, ���仯�Ľ����Ǽ��������.���, ����ڵͼ��Ӿ��̼����о�, �÷�ʽ�ر��ʺ������о���Ƥ��Ĺ����������.��Ϊ��FFA�ȸ�Ƥ��, �����Ļ�Դ̼����������ѡ����, ���ڱ���ν�����һ�ִ̼�, �������ֽ�����������������������̼��շ��Ļ���Ǽ��������շ��Ļ.������������ʾ���䷶ʽ֮��, ���ǿ��Կ��Ʋ�ͬ�����������̼��շ��Ļ���ֲ���, ����IJ���ɹ���ڼ������ݵIJ���.
����1�� �����뷽��
����1.1�� ����
����ʵ��ı�����16�� (7������, 9��Ů��) �������ϴ�ѧ�Ľ��������������о����μ��˱��ε�����̬���ܺ˴Ź������ݲɼ�, ���Ե�����Ϊ19~26��֮��, ƽ������Ϊ22.4��.���еı����ڽ��к˴Ź���ɨ��ǰ����д�����ұ����ʾ��Խ���ɸѡ�ų�����ʷ������ʷ���߷�����ҩ��ʷ�ı���.����, ÿ�����Ե������������߽�����������.���κ˴Ź���ʵ��ͨ�������ϴ�ѧ����ίԱ�����, ͬʱÿ��������ʵ�鿪ʼǰ����д����ص�֪��ͬ����.�ں˴Ź���ʵ����������ÿ��������Ӧ�ı���.
����1.2�� ���������
������Ϊʵ�����漰��ʵ����������ͼƬ�ͷ���ͼƬ, ���߸�60��.���ͼƬ��Դ���������ͼ��[24].���ڱ�ʵ����Ҫ̽�������Ӿ���������ά�ֽε���֪����, ��Ҫ�ų����������������ʵ�ͼƬ, ������е����ͼƬ��Ϊ�������ͼƬ.����ͼƬ��Դ�ڻ�����.���е�ͼƬ��ͨ��Matlab����ͳһ�ӹ�����, ������ͼƬ�ڳߴ��С (138 px×151 px) �����ȡ��Աȶȵ������϶�����һ��.����������Ϊʵ��̼����Ƿֱ���Ϊ1024×768, ˢ����Ϊ85 Hz���������߹���ʾ�� (cathode ray tube, CRT) .ʵ�����ͨ��Matlab�������б�д, ��ʵ���, ���������ͨ��ͬMatlab���ݵ�Psychtoolbox������������.��ʵ�鿪ʼ֮ǰ�Ա��Խ���ָ����Ľ���, ���ں˴������Ϊʵ����ͨ�����ݵ���ϰʵ���ñ��ԶԲ������ݺ�ʵ��Ҫ�������Ϥ�����������.�����ߵ��°ͱ�Ҫ��̶����Ӿ�ʵ��ר�õ�ͷ����, �۾�����Ļ�ľ����ԼΪ57 cm, ѡ��8������, ʵ����ϱ����������ѡ����������, �ӽ�Ϊ4.59°×5.54°.
������ʽʵ���ʵ��������ͼ1.��ͷ���ͼƬ������ij�����ѡ����4��λ���е�1����2��λ����, ÿ�ζ�����600 ms.�ڸ�����Ϊ2��ͼƬ��Ϣ�������, ���������1������߷���ͼƬ, ���Ա�Ҫ���ס����ͼƬ, 400 ms�����ٳ���1����һ���ͼƬ, ����ͬ����Ҫ���ס��ͼƬ (����ͼƬ) .���̼����ֽ���400 ms��������������, ����������Ϊ“1”��ʱ��, Ҫ����Ŭ��ά�ֵ�һ�����Ӿ���Ϣ, ����������Ϊ“2”��ʱ��, ������Ҫά�ֵڶ���ͼƬ���Ӿ���Ϣ.֮������1������ͼƬ, ����ͼƬ��50%�ĸ���������Ҫ���Լ�ס��ͼƬ (��ͼ) , 50%�ĸ�����Դ��ʣ�µ�ͬ������ͼƬ (��ͼ) .������ͼƬ�����ͼƬһ�µ�ʱ���ð������.�����Դ̼��ͼ���̼���һ�µ�ʱ��, ���Ҽ�.���Ա���֪��Ҫ���ֿ�ؽ��а�����Ӧ.�ڽ���������һ��12 s�Ĵ��к�ɫע�ӵ�Ŀ���, �ñ��ԵĴ��Եõ��㹻����ϢͬʱҪ������˼���ղŵ�ʵ����Ϣ.�ڸ���Ϊ4��ͼƬ��Ϣ�������, ǰ���������ֵĴ̼��ֱ�Ϊ2�������2�ŷ���ͼƬ��Ϣ, ǰ��˳�����.����֮�������IJ����������Ϊ2��ͼƬ��Ϣʱ��ȫһ��.
����ͼ1 (������ɫ) ʵ������ͼ.����θ���Ϊ2��ͼƬ (a) ��4��ͼƬ (b) ��Ϣʱ���Ӿ�����������Ϊʵ������ͼ.���ȳ��ְ�����ͷ��ݵļ���̼�, Ȼ�����������ʾ��������ͼƬ, ����11 s�ļ���ά�ֽκ�, ���ֲ��Դ̼�.������Ҫ�жϲ��Դ̼��Ƿ�ͼ���̼��е�ͼƬһ��Figure 1 (Color online) Experimental procedure.The procedure of the visual working memory behavior experimental when the encoding stage load is 2 pictures (a) and 4 pictures (b) information.First, present memory stimuli including faces and houses, and then present clues to remind which pictures to remember.After 11 s of memory maintenance phase, test stimulation is presented.Subjects need to determine whether the test stimulus is the same as the one in the memory stimulus

������ʵ���Ϊ20��run, ÿ��run����8��trial.ÿ��������40��trial, һ��160��trial.�ڱ��Խ���ʵ���ʱ��, ÿ��run������ʱ����Ϊ5 min.ÿ����1��run, ������3 min��Ϣʱ��, �Ա��´�ʵ����Ծ�������ͻ����Ӿ�ƣ��.�˴�ɨ���˳��Ϊ:�ṹ�����ݲɼ� (Լ8 min) , �ͼ��Ӿ�Ƥ�㶨λɨ�� (Լ15min) �Լ���ͷ�����Ȥ��ɨ�� (Լ15 min) , ��ʵ�� (Լ2 h) .�����ÿ��ʵ��֮��, �����ں˴�����һ������Ϣʱ�� (����ͷ������) .���ǵ�ʵ��ʱ��ij��̺�ÿ�����Եľ�����ע������������, ÿ�����Ե�ʵ�鶼�ֳ�3 d���� (�ڼ������ͬ�������) .��1�������Ȥ���Ķ�λ�˴Ź���ɨ�� (Լ50 min) , ��2�������ʵ��ǰ��10��run�ĺ˴Ź������ݲɼ� (Լ1 h) , ��3����ɵ�11~20��run�Ĵ��Թ��ܺ˴Ź���ɨ�� (Լ1 h) .����, ÿ��ɨ��ǰ��Ҫ�Ե���Ľṹ����вɼ��Ա���������������ܽ�����Ĺ�����ͬ����Ľṹ�����.
�����Եͼ��Ӿ�Ƥ�� (V1~V4) �Ķ�λ�õ��˾��������Ĥ����Ƥ�㶨λ���Ϻͷ�ʽ[25].��ɨ��һ������33�㸲��ȫ��.�̼�ͨ����˴ż��ݵ�ͶӰ��ͶӰ����������Ļ��, ����Ļ�ķֱ���Ϊ1024×768, ˢ����Ϊ60 Hz.�������ں˴���ͨ�������۾��Ϸ��ķ��⾵�۲�̼�����.����� (FFA, STS) ��������Ȥ�� (PPA) �Լ����ڹ� (IPS) ����Ķ�λʱ, ��Ļ�ϻ��������3�ִ̼�����:���ͼƬ������ͼƬ��ģ��ͼƬ.���ͼƬ��Դ���������ͼ��[24]������2��ͼƬ����һ�µ�������а�����Ӧ (����ʳָ) .ʵ�����������, ÿ��ͼƬ����200 ms, ͼƬ֮����300 ms, 24����ͬ����ͼƬ���һ�����.���֮����12 s.ÿ��run����3������5��.ÿ���������2��run�Ĵ���ɨ�貢��33�㸲��ȫ��, ����ʱ��ԼΪ15 min.�����ṩ������2������������ɰ�����Ӧ.ʵ��ȫ����Ҫ���Ա���ͷ����Ҫ��, ����ע������ɵ�ǰ��ʵ��, Ҫ���Բ�Ҫ˵��, ��Ϣ���ȶ���ͨ��������Ӧ�����.
����2�� �˴Ź������ݵIJ����ͷ���
����2.1�� �˴Ź���ɨ������Ķ����Ԥ����
����ÿ�ν���ʵ��֮ǰ������нṹ��IJɼ��Ա��ں������ݷ�����ʱ����Խ��ṹ���������ж���.�ṹ�������IJɼ�������������3.0 T�˴Ź���ɨ���� (Siemens Medical, �¹�) .���Ե�ͷ�������̶���ɨ���ǵĺ���Ȧ��.�ôŻ������ٲɼ��ݶȻز� (magnetization-prepared rapidacquisition gradient echo, MPRAGE) ���в������ɼ����и߷ֱ��ʵ�T1��.�ظ�ʱ�� (repetition time, TR) =2000 ms, �ز�ʱ�� (echotime, TE) =2.52 ms, ��תʱ�� (inversion time, TI) =1900 ms, ��ת��=9°, �������=256×256, ���=1.0 mm, ���ش�С=1 mm×1 mm×1 mm, ɨ��176�㸲��ȫ��.
��������������ݲɼ����õ���12ͨ����Ȧ, ����ΪT2��Ȩ�ݶȻز�ƽ��������м���.�ظ�ʱ�� (repetition time, TR) =2000 ms, �ز�ʱ�� (echo time, TE) =30 ms, ��ת��Ϊ90°, ���Ϊ3 mm, ��Ұ (field of view, FOV) =192 mm×192 mm, ����=0, ����Ϊ64×64, ���е�ɨ����Ҷ�ײ��������Ͽ�ʼɨ��.����������������Brain Voyager QX��������������Ԥ��������Ȥ���Ļ���.Ԥ�����IJ������:�ṹ��Ԥ�������ṹ���빦������롢ͷ�����������������Ƴ��Լ���ͨ�˲� (0.015 Hz) �ȹ���.ͷ�������Ĵ���������:6��ͷ�����������ܴ���3 mm, ���ᱻɾ��.��4λ����ͷ������, �Ͻ����, ���ܽ��к����о�, �������ݱ�ɾ��.���, �ں����йع��ܺ˴Ź������ݷ�����, ��ʵ�齫ֻ�漰12λ���Ե�����.
����2.2�� ���ݷ���
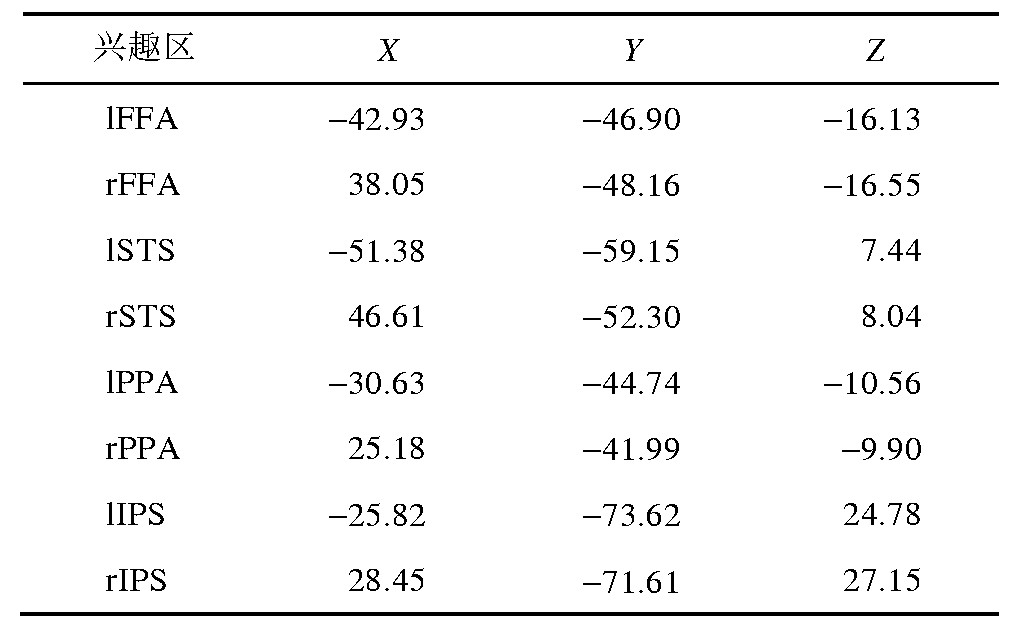
����(��) ������Ȥ��.�ڽ���ÿһ��������ݷ���֮ǰ, ���ǽ�ÿһ�����ԵĽṹ�����ݵ���ǰ���ϵ��ռ�ԭ��, �����ϵ���������ƽ�е�λ��.��ʵ������Ȥ�����������Ӿ�Ƥ���Լ���ס�������Ȥ��.��ʵ���ע�ĵͼ��Ӿ�Ƥ����Ҫ��V1, V2, V3, V4.������Ƥ����Ҫ��˫����״�� (bilateral fusiform face area, l FFA, r FFA) ��˫�ຣ���Ի� (bilateralparahippocampal place area, l PPA, r PPA) ��˫����Ϲ� (bilateral superior temporalsulcus, l STS, r STS) ��˫�ඥ�ڹ� (bilateral intraparietal sulcus, l IPS, r IPS) .���Ƚ�Ԥ����֮��Ĺ�����������ϳ�һ������ģ�� (general linear model, GLM) .�ڵͼ��Ӿ�Ƥ��Ķ�λ��, ���ö�λ�������ݰѼ�������ڱ�ƽ��Ƥ�ʱ���, Ȼ���������Ĥ��λͼ����ÿ���Ӿ�����.������, ͨ�������Ӿ������ʵ������ǿ�ȴ�С (P<0.001) , �Ѽ��������ŵ�������ΪV1~V4��IPS.�ڶ���ͷ�����Ȥ���Ķ�λ��, ���ⶨ��GLMģ����, ���о��������ͼƬ�ļ����ȥ����ͼƬ�ļ�����Ϊ�����Ȥ���Ķ�λ, �÷���ͼƬ�ļ����ȥ���ͼƬ�ļ�����Ϊ������Ȥ���Ķ�λ, ����ֵ�趨ΪP<0.001.����Ƥ��ÿ����Ȥ����ǿ����Talairach������ϵ�е��������1 (�ͼ���Ƥ��Ϊ����ϲ�����) .
����(��) ����ǿ�Ⱥ�ģʽ����.����ǿ�ȷ�������������������ᵽ��12����Ȥ�� (V1, V2, V3, V4, l FFA, r FFA, l PPA, r PPA, l IPS, r IPS, l STS, r STS) , ���õ���ƽ��������.��Ը�����Ȥ����4�ֲ�ͬ������, �Ե�һ�ż���ͼƬ��ʼ������Ϊ���, ��ȡÿ��TR��Ӧ��BOLD�ź�ֵ.ÿ��trial����16��TR.Ȼ���ٽ���ͬ�����µ�����BOLDֵ����TR��˳����е���ƽ��, �õ�ÿ�������ļ�������ͼ.
�����ڼ���ǿ�ȷ�����, ���Ƿ��ֵ�5�͵�6��TR�������ʺ���Ϊ�Ӿ���������ά�ֽ��о�����, ��Ϊ��5��TR��ʼ�պ��Ǽ���ǿ�Ⱦ�����ֵ֮��, �÷�ֵ�������Ǵ̼����ֽεĴ��Լ��� (��������������ǿ�ȷ������) .�ڵ�6��TR����֮��, ��Ȥ������ǿ��ֵ����ƽ��.���������ȡÿ��������ÿ��trial�ĵ�5~6��TR, Ҳ����10~12 s������, Ȼ�����߽���ƽ��, ��Ϊά�ֽεļ���ǿ��ֵ.
������1 ��Ȥ����Talairach������ϵ�е�����Table 1 The coordinates of the region of interest in the Talairach coordinate system
����Ϊ��̽�����Ӿ���������ά�ֽγ����Ӿ�Ƥ�����Ȥ���Լ������ݵĽ���, ���Dz����˶�����ģʽ�������� (multivoxel pattern analysis, MVPA) .�ڼ���ģʽ������, ���ݼ���ǿ�ȷ������ѡ���˵�5����6��TR��������Ϊ�Ӿ���������ά�ֽ��о�����.������ģʽ���������ڶ�������źż����ϲ����俴��һ����ά����, �ö�ά����������Щ���صĿռ�ģʽ.ͨ���������ķ�ʽ (������, classifier) �����ڲ�ͬ����֪״̬�¾��е��ض��ռ�ģʽ.������ģʽ�������������ڶԶ�����ؼ��ϵĿռ�ģʽ�ķ���, ��˱����˶Ե������ط���ʱ���е�����ȵ͵����ش������źż�������ȵ͵�����.
����������ģʽ�����������¼�������:��һ��Ϊ����ѡ��, ����ѡ����Է�Ϊ��ȫ������Ϊ��������Ȥ������Ϊ����.������ȫ������Ϊ�����ķ������й��������, ���Ա��о���ȡ�Ը���Ȥ����Ϊ�����ķ���.�ڶ����Ǵ�������, ����4��������ά�ֽμ���Ŀռ�ģʽ.������Ϊѵ��������, ָ���ǽ�ѵ������������������Ա�������ڶ���Щ����ѧϰ�����ֲ�ͬ�����.���IJ�Ϊ��������, ��Ϊѵ���ķ������, �����������δ֪�������ݿռ�ģʽ������������ܷ���ȷԤ��ģʽ���, ���о���ʹ�õ���leave-one-run-out�ķ���.�ڹ��ܺ˴Ź����о���, �о���һ��ϣ�������ź�ģʽ�н���Ȥ������֪״̬�������, �����������Ը���50%����ȷ�ʶԲ�ͬ��֪���̽��н���.���о��н��ж�����ģʽ�����Ĺ��߰�Ϊlibsvm (http://www.csie.ntu.edu.tw/~cjlin/libsvm/) [26].
����(��) ȫ�Է���.����Ȥ���ķ���������, ���о�û�з�����ֱ�������Ӿ��������为��ЧӦ���������������֪��ָ��.����, ���о�������ȫ�������ķ������������Ƿ����ijЩ�ض����������Ӿ��������为��ЧӦֱ�����.����, ���ÿ��voxel, ��ȡ��ά�ֽεļ���ǿ��ֵ (��5���͵�6��TR��ƽ��) .���, ��ÿ��voxel��4����������2×2�ķ������, �ҳ��������ú���ЧӦ���ŵ�����.ȫ������������ΪFDR���ؼ���, P<0.05, ����>200.
����3�� ʵ����
����3.1�� �����
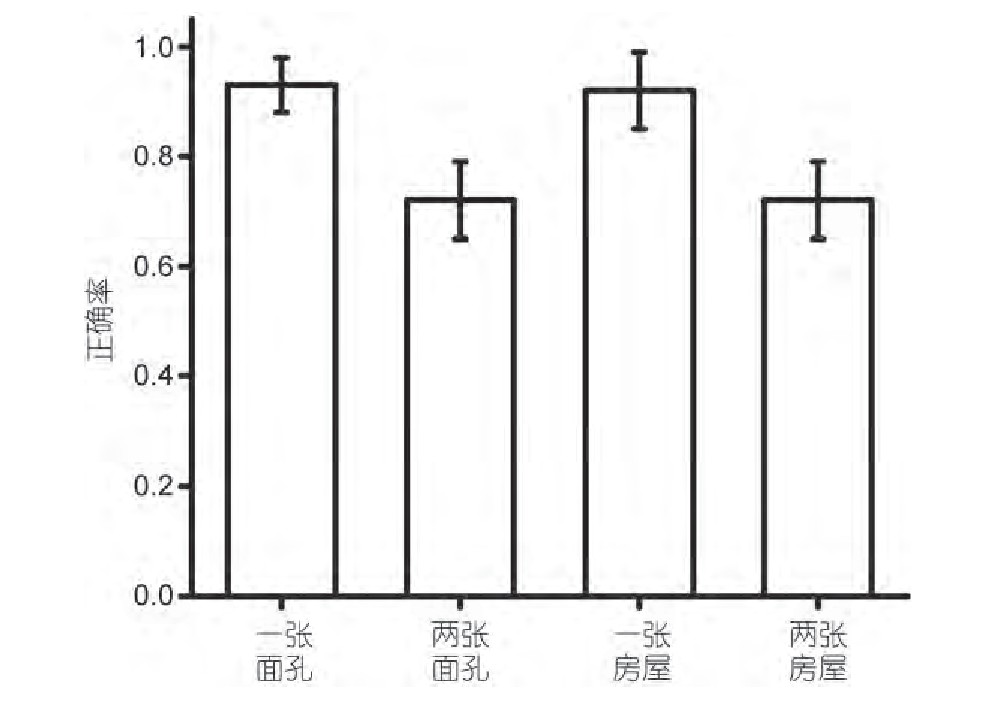
�������ǽ�ʵ������а����ķ�Ӧ�����˼�¼��ͳ�ơ�������ÿ�������µ���ȷ��, �����ͼ2.��Ϊ�����ʾ:ά��1�����ʱ��Ϣ��ƽ����ȷ��Ϊ0.93 (S D=0.0 5) , ά��2�������Ϣƽ����ȷ��Ϊ0.7 2 (SD=0.07) , ά��1�ŷ�����Ϣ��ƽ����ȷ��Ϊ0.92 (SD=0.07) , ά��2�ŷ���ͼƬ��Ϣ����ȷ��Ϊ0.72 (SD=0.07) .��4�������µ���ȷ�ʽ����ظ������Ķ����ط������, ����ЧӦ��ЧӦ���� (F (1, 11) =233.794, P<0.001) , ����������ЧӦ���������ò�����.����, ��4��������, ���е���ȷ�ʶ����Ÿ������ˮƽ, �ɼ����������DZ����ܹ�ʤ�ε�, ���ǵ�ʵ����������Ч�ط�ӳ�˱��Ե��Ӿ��������������.��Ϊʵ���ڸ���ЧӦ�ϴﵽ�����ʵ��ʱ��Ԥ��, ͬʱҲ�������Ӿ���������������������, ����ά�ֵĿ�����Ϣ��������ͻ������Ӿ���������ĸ���ЧӦ.
����ͼ2 �Ӿ���������˴�����Ϊʵ����ͼ.��������ЧӦ����, �̼�������ЧӦ������, �������ò�����Figure 2 Visual working memory behavior experimental results.The main effect of load was significant, the main effect of stimulating mate-rial was not significant, and the interaction was not significant
����3.2�� ����ǿ�ȷ������
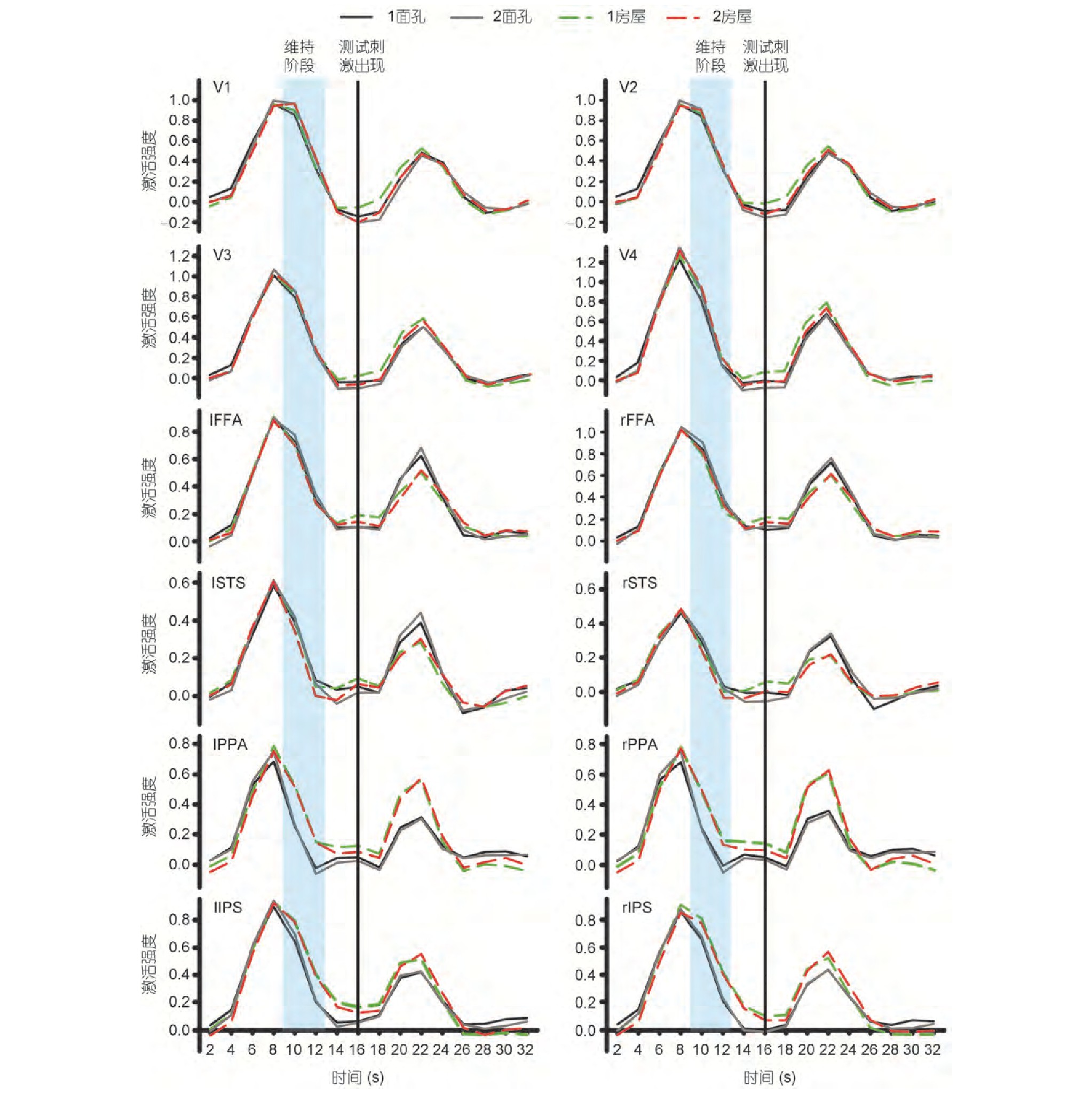
��������������ȡ�˼���̼����շ����źŷ�ֵ���бȽ�.��ͼ3��ʾ, ����̼��շ����ź��ڵ�4��TR����8��ʱ�ﵽ��ֵ.���, ������ȡÿ�����Եĵ�4��TR��BOLD�ź�ֵ������2 (�̼�����) ×2 (����) �������.�ظ��������������ʾ, ���������Ľ������þ�������, �̼�������ɵ���ЧӦҲ��������.��һ���˵��, �źŵķ�ֵ��Ҫ��ӳ���Ƕ������̼�����Ӧ, ���Ƿ�ӳ�������������.������һ�Ʋ����������, ���ָ����Եļ���̼���ÿ��trial�ﶼ�Ǽ�����״̼�, ���з��ݴ̼�, ���DZ��Լ���ά�ֵ�����ֻ������һ��.�����ֵ��ӳ�˼���ά�ֵ����ݵĻ�, ��ô�ڼ��䲻ͬ����Ĵ̼�ʱ, ��������������ͷ��������������ķ�ӦӦ�ò�һ��.���, ���ǵ��о�����ֵ�ź��ų����˼���ά�ֽε��о���Χ֮��, �����÷�ֵ�����Ҳ��Դ̼�����֮ǰ���ź����о�����ά�ֽε��.����, �����ų��˵�7��TR������, ��Ϊ��TRʱ�ź��Ѿ�����������ˮƽ, ���ܴ��ڵذ�ЧӦ.������������ѡȡ�����о�ά�ֽε��ź�Ϊ��5, 6��TR (��10~12 s) ���ź�.
����ͨ����4������ά�ֽμ���ǿ�ȵķ�������Ա�, ����:��ά�ֽ�, ˫�ຣ���Ի� (PPA) ��˫�ඥ�ڹ� (IPS) �ļ���ǿ���ڶԷ�����Ϣ��ά�������Ÿ��ڶ������Ϣ��ά��, ��������ЧӦ���� (F (1, 11) l PPA=41.66, P<0.001;F (1, 11) r PPA=29.58, P<0.001;F (1, 11) l IPS=29.09, P<0.001;F (1, 11) r IPS=28.75, P<0.001) , ����������δ���ֳ��̼����͵���ЧӦ.����, ���������ĸ�����ЧӦ�Լ��̼����ͺ��ɵĽ������þ�������.����ǿ�Ƚ������, ���ǿ���ͨ�������� (PPA) �غͶ��ڹ� (IPS) �ļ���ǿ�����жϱ������Ӿ���������ά�ֽεļ�������, ���ǽ����Ӽ���ǿ�ȷ��滹�����ж��为����.������״�� (FFA) ����Ϲ� (STS) �Լ��ͼ��Ӿ�Ƥ����˵, �������ݲ��ܸ����伤��ǿ�����ж�.
�������, ���ǶԲ��Դ̼��շ����źŽ����˷���.��ͼ3���Կ���, ���Դ̼��շ����ź��ڵ�11��TR (��22 s) ʱ�ﵽ��ֵ, ���������ȡ�������ֵ�źŽ�����2×2�������.�������, �ͼ��Ӿ����Ľ������ú���ЧӦ��������.���ѡ�����ͷ���ѡ�����Ĵ̼�������ЧӦ���� (F (1, 11) l FFA=27.07, P<0.001;F (1, 11) r FFA=11.90, P=0.005;F (1, 11) l STS=15.66, P=0.002;F (1, 11) r STS=20.66, P<0.001;F (1, 11) l PPA=49.36, P<0.001;F (1, 11) r PPA=57.47, P<0.001;F (1, 11) l IPS=18.46, P=0.001;F (1, 11) r IPS=13.03, P=0.004) , ���Ǹ��ɵ���ЧӦ���������þ�������.��һ����������ǵ�Ԥ��, �����ѡ��������״̼��ķ�Ӧ���ڶԷ��ݴ̼��ķ�Ӧ;��֮, ����ѡ�����Է��ݴ̼��ķ�Ӧ���ڶ���״̼��ķ�Ӧ, ˵�����ǵ�ʵ����ƺ���Ȥ����ѡȡ����Ч�ĺ���ȷ��.
����3.3 ����ģʽ�������
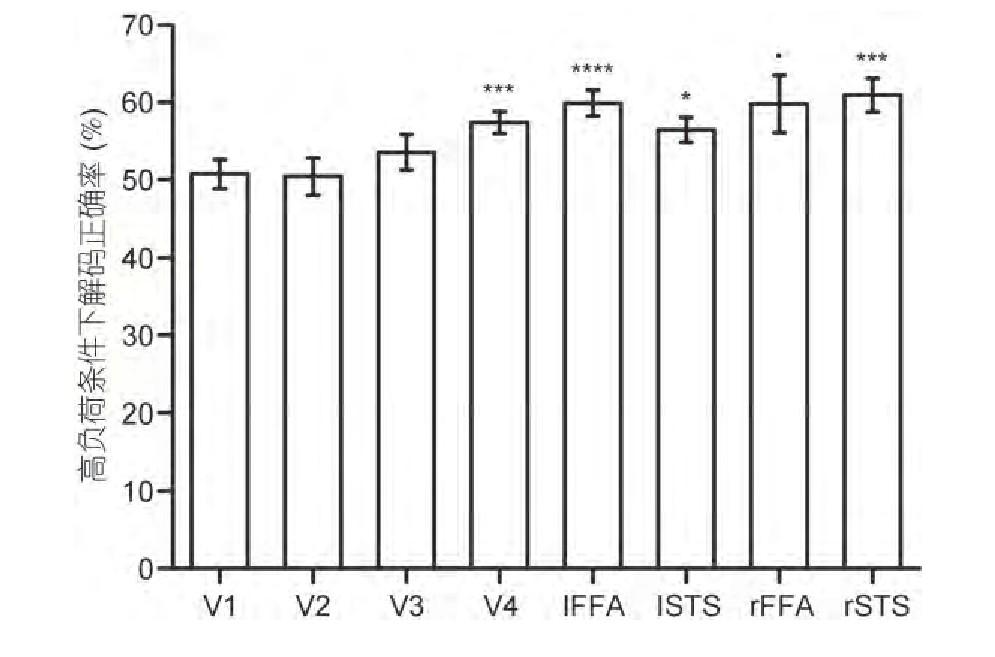
��������˫���PPA��IPS��4�������ڼ���ǿ�ȷ������Ѿ����ڶ���ͷ��ӵ����Ų���, ���������м���ģʽ����.����ģʽ�������漰����Ȥ��Ϊ8��, �ֱ���V1~V4��˫���FFA��˫��STS.����ȡ����ά�ֽ� (��10~12��) ÿ�������ļ���ģʽͼ, ���û���ѧϰ�ķ�����֤����ģʽ�Ƿ�������ּ�������������Ƿ���.�ڵ��� (ά��һ��ͼƬ��Ϣ) �������, ���������Ľ�����ȷ�ʾ������Ÿ���50%�����ˮƽ.���ڸ߸��� (ά������ͼƬ��Ϣ) �������, �ܶԷ�����Ϣ�������Ϣ�������ֵ���������:V4 (T (11) =5.17, P=0.002, Bonferroni���رȽϽ���) , ��������Ϊ57.40%;���FFA (T (11) =5.91, P=0.0008, Bonferroni���رȽϽ���) , ������Ϊ59.90%;���STS (T (11) =3.99, P=0.02, Bonferroni���رȽϽ���) , ������Ϊ56.46%;�Ҳ�STS (T (11) =5.05, P=0.003, Bonferroni���رȽϽ���) , ������Ϊ60.94%;�Ҳ�FFA (T (11) =2.65, P=0.02, δ����) , ������Ϊ59.80%.����ģʽ�����ʾ:�����ڸ߸�����������Ȥ���������Ϣ�ͷ�����Ϣ�Ľ�����ȷ�����Ÿ���ƽ��ˮƽ, �����ͼ4.
����3.4�� ȫ�Է������
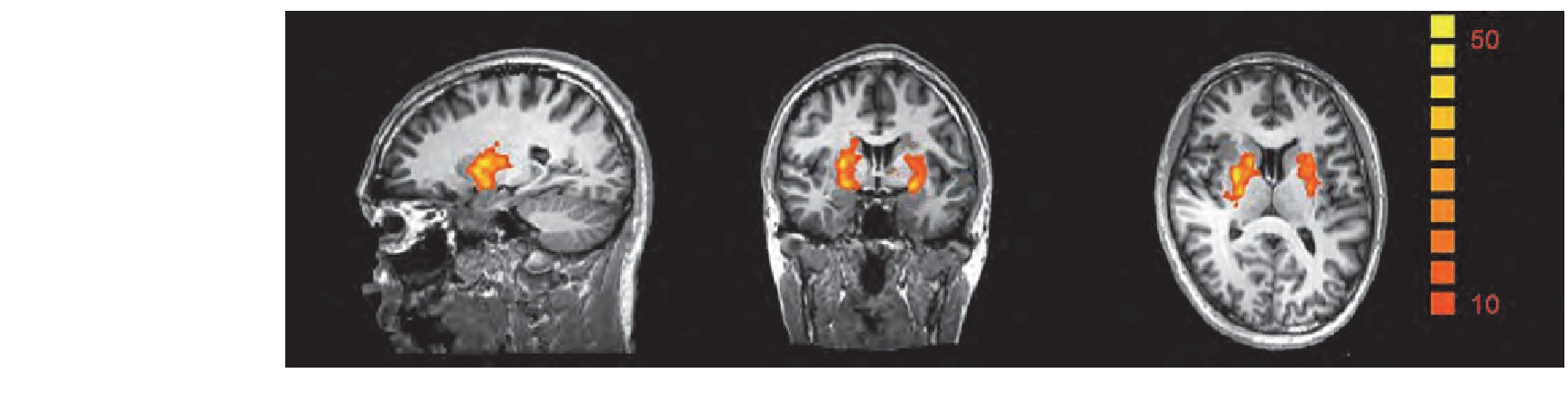
������ȫ�Է�����, ���о����ö�ÿ��voxel�ļ������2×2�������, �����ʾ, �ڼ���ά�ֽ� (��5, 6��TR) ��, �̼������������ЧӦ����Ȥ�������һ��:������Ϣά�ֽ������ں��������Ͷ��ڹ��ļ�����.����������ЧӦ������˫��Ǻ� (putamen) �ļ���ǿ���� (FDR���ؼ���P<0.05, ����>200) .�̼����Ϻ�����֮�䲻���ڽ���ЧӦ, ����, ��ʵ���еĸ���ЧӦ�����ڿǺ˵ļ���ǿ���ж������ܵ��̼����������Ӱ��, ���Ǻ˵���ǿTalairach����Ϊ (20, 4, 2) , �Ҳ�Ǻ˵���ǿTalairach����Ϊ (-24, 4, 8) .��������ЧӦȫ�Է������ͼ��ͼ5.
����ͼ3 (������ɫ) ����ǿ�Ƚ��ͼ.V1��V2��V3��V4��˫��FFA��˫��STS��˫��PPA��˫��IPS��Ȥ���ļ���ǿ�Ƚ��.˫��PPA��IPS���Ӿ���Ϣ���ֽ� (��10~12��) �ļ���ǿ�ȿ������ּ���ά������, ��Է�����Ϣά���еļ���ǿ�����Ÿ��ڶ������Ϣά���еļ���ǿ��.Ȼ��, ������Ȥ���ļ���ǿ���ڶ������Ϣ�ͷ�����Ϣ��ά�ֻ�в��������Ų���Figure 3 (Color online) Activaty intensity result.Activation intensity results for regions of interest for V1, V2, V3, V4, bilateral FFA, bilateral STS, bilateral PPA, and bilateral IPS.The activation intensity of the bilateral PPA and IPS in the visual information retention phase (10-12 s) can distinguish memory-maintaining content, which is significantly stronger in activation of house information maintenance than in activation of face information.However, there is no significant difference in the activation intensity of other areas of interest in the maintenance activities of face information and house information
����ͼ4 ����ά�ֽ��и߸��������¶�����ģʽ���� (MVPA) ���.V4��˫��FFA��˫��STS������ά�ֽα����Ӿ�����������Ϣ, �������ȷ�����Ÿ���50%.�ý������Bonferroni���رȽϽ��� (*:P0.05;***:P0.005;****:P0.001;·:P=0.02, δ����) Figure 4 Multivariate pattern analysis (MVPA) results under high load.V4, bilateral FFA, and bilateral STS were able to characterize visual working memory during the maintenance phase, and their decoding accuracy was significantly higher than 50%.This result was corrected by Bonferroni multiple comparison (*;P0.05;***:P0.005;****:P0.001;·:P=0.02, uncorrected)
����4�� ����
����4.1�� ������������Ϣ�ĸ���ЧӦ
������Ϊ�������, �ڶԶ�����������Ϣ���Ӿ��������������д������Ÿ���ЧӦ, �����ż�����Ŀ������, ����ȷ�ʻ������½�, �������ͺͽ���ЧӦ��������.�ڱ��о���, �̼����Ϸ�Ϊ���ͼƬ�ͷ���ͼƬ, ���и�������ЧӦ����, �̼�����������ЧӦ�ͽ���ЧӦ��������, ��ǰ���о��������.һ���ֹ����Ӿ�����������о���, ���ò�ͬ�Ĵ̼����Ͼ��������Ӿ��������为��ЧӦ[2,27].���ǵ�ʵ����, ���Ӿ���������������ŵĸ���ЧӦ, �Ӿ����������������ֳ�������.֪������ЧӦ������Ϊ�����ڶ���֪����Ϣ���мӹ�ʱ����֪��Դ������[28].������֪��Դ��Ҫ���䵽�Ը��ӿ�����Ϣ�ļ���ά����֪�ӹ�������, ÿ��������Ϣ����Ҫһ��������֪��Դ, ���Ե���ÿ��������ά�ֽ϶���Ϣʱ��֪��Դ��������[29].�ڵ���������, ������֪��Դ���Գ���ط����ڶ�ÿ��������Ӿ���Ϣ�ӹ�������.�ڸ߸���������, ������֪��Դ���ܳ���ط��䵽��ÿһ������ľ�ȷ����ά�ֹ�����, ���Ե����Ӿ�����������ȷ�������½�.֪������������ע����Դ�Ƿ�õ���Ч�ӹ��������Ӿ��������������еĸ���ЧӦ[28].����һ�������Ӿ��������为��ЧӦ������ƫ�ྺ��ģ��[30].��ģ��ǿ���������Ӿ��������������л���ڸ�����������ص����϶��µ�ע�����.�ڵ���������, ע����Դ�Ἧ���ڵ�ǰ����������;�ڸ߸��������»���Ϊע����Դ�ķ��併�ͼ��侫��.����, ���ڹ�������ľ������ۻ���Baddeley�Ĺ���������ϵͳ����, ��������Ϊ�ڹ��������д���3��ϵͳ:����ִ��ϵͳ���Ӿ��ռ���±���������·.����, ����ִ��ϵͳ����ע����ƽ��е�ǰ�ļ���ά������[1].��Щ���ۿ��Խ������Ӿ�����������Ϊʵ���д������ŵĸ���ЧӦ������������.
����������Ϊʵ��������ʾ������ЧӦ�Ĵ���, ǰ��ʵ���һ����֤�����о����ȶ��ԺͿ��ظ���.����������ЧӦ������, �ҽ������ò�����, ���Ա��о��е�ʵ������δ�ܵ������Ƿ�ƫ�ü��䷿����Ϣ�������Ϣ��Ӱ��.���, ���о��е���Ϊʵ�����, �ڸ��ӿ�����Ϣ���Ӿ���������������Ҳ�������Ӿ������������������Ժ���ЧӦ, ���Ҳ����ܵ��̼����������Ӱ��.
����ͼ5 (������ɫ) ȫ�Է���������ЧӦ���ŵ�����.FDR����P0.05, voxel>200Figure 5 (Color online) Whole brain analysis results.FDR correction P0.05, voxel>200
����4.2�� ��������ά�ֽ��Ӿ�ϵͳ�������Ϣ�ı���
��������̬���ܺ˴Ź������ݽ�����Ȥ��������ʾ, �ɼ���̼����շ���BOLD�ź�ǿ���ڷ�ֵʱ��������״̼��ͷ��ݴ̼�.��һ���˵����ֵ�ź���Ҫ��ӳ���Ǵ̼������������շ��ķ�Ӧ, ��������Ĺ�ϵ��С.���ڲ�ȡ���º������ķ�ʽ��ʾ���������, ��˱����˼�����������������ϵ�һ��.����, ����ڼ�������ά�ֽ�, ����źų�������ͷ���֮��IJ���, ��ɹ���ڼ������ݵIJ�ͬ, ���Dz����������ԵIJ�ͬ.����������Ԥ�ڵ�, ��ά�ֽ�, ���Ƿ���˫��PPA��˫��IPS�Է���ͼƬ�ļ���ǿ�����Ÿ������ͼƬ.������ά��1�ŷ���ͼƬ��Ϣ��2�ŷ���ͼƬ��Ϣ֮��ļ���ǿ�Ȳ��첻����.�÷������˵��, �ڱ�ʵ����, ͨ��PPA��IPS���Ӿ���������ά�ֽεļ���ǿ�ȴ�С�Ϳ�������ά�ֽεļ��������Ƿ�����Ϣ���������Ϣ.ǰ�˹��ں����������о�����, �伤��ǿ���뷿�������Ϣ��������[31,32].�ڶԷ�����Ϣ�������Ϣ���ƶ�ע�ӵ���Ϣ��ѡ����ע����, ������������״�غ��˶�������������Ϣ�ļ���ǿ�ȷֱ�ϸ߲��������Ų���.����, ��3��������������ִ̼�Ҳ����һ��ǿ�ȵļ���[31].��֪�����̵���, ���������ļ�����Ԥ��Է�����Ϣ�ļӹ�����[33].��������������̼� (����������Ϣ) �ļ�����롢����ά���Լ����������ж����������Ŵ���Ѫ���źż���.ǰ�˹��ڼ�����о�����, ���ֺ��������������ڼ����ά�ֽλ������Ͻζ�������ʮ����Ҫ������[34,35].�ڹ������������������, ���������ڱ���κ�ά�ֽζ������ƫ�õĴ̼� (����) �������ŵļ���[35].�ڱ��о���, ��ֱ�ӵ�֤�ݱ���, ���������ڹ�������ά�ֽεĻ�ͼ���������и߶ȵ����.
������ʵ���п���ͨ�����ڹ����Ӿ���������ά�ֽεļ���ǿ�����������ά������Ϊ����ͼƬ��Ϣ.��Ȼ�ڶ����ͼƬ��Ϣ�ͷ���ͼƬ��Ϣ�ļ���ά���ж�������˫�ඥ�ڹ��ļ���.����, ��ά�ֹ�����, ���ڹ��Է�����Ϣ�ļ���ǿ�����Ÿ��������Ϣ, �ò��������Դ��ά�����ݵIJ�����.����, ���ڹ��ڿռ�ע��ӹ�����Ĺ������ܵ��㷺�о�[36].�������������ṩ�˸���Ŀռ�������Ϣ, ��˶��ڹ��Է��ݵķ�Ӧ��ǿ.���ǵ��о�����, ���ڹ������ڴ̼����ֽζԷ��ݵȿռ���Ϣ���мӹ�, �ڴ̼���ʧ֮����Ȼ������ӹ��ռ�������Ϣ.
�����������ϵ���, ���ѡ�������� (FFA��STS) �ڹ�������ά�ֽεĻǿ����������״̼��ͷ��ݴ̼�.ǰ�˵��о�����FFA�ͻ��������ά�ֵ����ݺ����й�[37~39].���ǵ��о���ǰ���о��IJ�������, ���Dz�������������ķ�ʽ, ��������Ҫ��ͬʱ��ס��ͷ��ݵ���Ϣ, Ȼ�����������ѡ������һ����Ϣ��ǿ���̼���.��Ȼ���ַ�ʽ�����������ԵĿ��ƱȽ��ϸ�, ���ǻᵼ���ڱ���ζ���ͷ��ݴ̼���������һ���̶ȵļ���.�����ѡ�����п��ܶ����ֶ��ݵļ���Ҳʮ������, ������BOLDˮƽ�ڱ��Բ�δ����������ʱ��Ҳ��ά��һ���Ƚϸߵ�ˮƽ.��һ������ѡ�����ͷ���ѡ������Ѫ������ѧ���߿��Կ�������.�ڼ�������Ϊ���ʱ, PPA�Ļ�ڴﵽ��ֵ֮�������һ���dz������½�, ˵�������Ϣ�ܵ��˽�ǿ������.��FFA�Ļ�ڼ��䷿��ʱ, ��Ȼ���������½�ˮƽ, ˵�������Ϣ��δ�ܵ���Ч������.�Ժ���о������ڸ���Ϊ��ʽ�м��������������ʾ��̽�ּ��䷿����Ϣʱ�����Ϣ�Ƿ�Ҳ��Ȼ�õ��˼ӹ�.����, �������������ֽ�, �����ñ��Լ��䷿�ݵ�������ʾ, �����ڲ��Խ�, ȴ���ַ��ݴ̼�.��ʱ, ���Բ�δ������״̼�, ������˵, ����Ϊ�ɼ�Ӧ�ýϲ�.������������Ȼ���ֳ�һ���̶ȵĶ���ļ�������, ����������Ϣ��δ�ܵ���ȫ������.
������Ȼ����ǿ�ȷ�����δ�������ѡ�����Լ������ݵ�������, �������ǵļ���ģʽ������ʾ, �ڸ߸��������, ���ǿ���ͨ��V4������״���Լ���Ϲ��ļ���ģʽ�������ڼ���ά�ֽεļ�������.ǰ��ͨ������ģʽ��������, ������Ƥ�� (V1~V4) ���Ӿ���������ά�ֽ���ͨ����ͬ�ļ���ģʽ�Բ�ͬ����Ĺ�դ���м���ά��[17].ͬʱҲ���о�����, ���Ӿ�����������Ϣά�ֽ�, �����Ӿ�Ƥ��������Ϣ�ͷ�����Ϣ�ܽ�������[22].�ڶԵͼ��Ӿ�Ƥ��ķ�������, ���о������������Ȥ����V1~V4, ���Ƕ�����ģʽ���������ʾֻ��V4�ܹ��ڸ߸��������¶Է�����Ϣ�������Ϣ������Ч�ؽ���.ǰ�˹��ڳ����Ӿ�Ƥ����о�����, �ͼ��Ӿ�Ƥ���ڶ���Ϣ�ӹ������п�������Ϊ����ͨ��, ��һ�����Ӿ���Ϣͨ��V1, V2���ݵ�V5�ٵ�����СҶ����;�ڶ������Ӿ���Ϣͨ��V1, V2���ݵ�V4�ٵ��Ҷ����[40~42].����, �ڱ��о���V4���ڸ߸��������¾��нϸߵĽ�����, �����Ӿ���Ϣ�������ȴ��뵽V1����ͨ��V4��, ������V4������Բ�ͬ�Ĵ̼����ֳ���ͬ�ļ���ģʽ.��״���ھ��������о�֮����Ϊ���ʶ����[43~45].��Ϲ������ڶ���������Ϣ���мӹ�ʱҲ����ֳ����ż���[46], ����Ϲ��ڴ�����ͬ�Ĵ̼�ʱ���ܱ����ż���.�ڱ��о���, ��״�غ���Ϲ���������ͬ���������¶������Ϣ�ͷ�����Ϣ�ļ���ǿ��û�����Ų���, Ҳ��ǰ���о����һ��[45,47,48].�ڱ��о������Dz�������״�غ���Ϲ��ļ���ǿ�����ж��Ӿ���������ά�ֽεļ�������, ��ͨ������ģʽ�������ǿ��Զ����ѡ�������������и�����̵���ʶ:���ѡ���� (��״�غ���Ϲ�) �������Ϣ��ƫ�ò������������ڶ������Ϣ�ӹ�ʱ�������ŵļ���, �ڸ߸���������, ������ͨ�����ѡ�����ļ���ģʽ���ֱ���ά�ֵ���Ϣ�Ƿ��ӻ������.����, ���ѡ���� (FFA��STS) ��V4���������ŵĽϸ߽�����ȷ��, ��������Ϊ�ڸ߸���������������Խ���, ������Ҫ���������Ŭ�����ܶ��Ӿ���Ϣ������Чά��.����Ϊ�ɼ����Կ���, �����ڵ��������µijɼ����Դﵽ90%����, ˵������Ȼ�н϶�ļ�����Դ���Լ������������Ϣ, ������Ը��Ŵ̼�������Ч������, �Ӷ����½�������̼�������Ƿ��.���ڸ߸���������, ���Եijɼ���Ϊ70%����, �����Ը��Ŵ̼���������, ����������һ����, ��˻Ὣ���м�����Դ�����ڶԼ���Ŀ��̼���ά��, �Ӷ�ǿ���Ըô̼��ı���, ���½�����ȷ�����.
����4.3�� ��������ĸ���ЧӦ
������ȫ�Է�����, ����ЧӦ����ЧӦ������˫��Ǻ˵ļ���ǿ����.��ǰ���о���, �Ǻ���ѧϰ�ͼ����з�������Ҫ����[49~51].�Ҳ����ʺ�����������ͿǺ�֮���ǿ�Ĺ������������˶Լ������ݸ��ߵ�������[52].����, �Ǻ�����ļ����ڹ������������и���������Ϣ��֪�ӹ�������شӶ����¸��幤�������������ֳ�����[49].�ڱ��о���, �Ǻ����������ŵĸ���������ЧӦ, ����������Ϊ�Ǻ�����ļ������Ӿ��������������и�ά�ֿ�����Ϣ�����й�, ��������Ϣ������.�ڵ��������º߸�����������Ҫά�ֵĿ�����Ϣ������ͬ�����˿Ǻ�����ļ���ǿ�Ȳ�������.���, �ڶ������Ϣ�ͷ�����Ϣ���Ӿ���������ά����, �Ժ�����ļ���ǿ���������Ӿ��������为��ЧӦ.
����5�� ����
�������о��Ӷ��������ӿ��� (�����Ϣ�ͷ�����Ϣ) ���Ӿ����������������̽�����ڼ���ά�ֽεĴ�����֪����.�о��������, �ڶԸ��ӿ�����Ӿ��������������б��ֳ��Ӿ��������������������Ժ���ЧӦ.ͬʱ, ���ǿ���ͨ��˫�ຣ��������˫�ඥ�ڹ����ļ���ǿ��Ԥ��ά�ֽεļ�������.Ȼ��, ����ֻ��ͨ��˫����״�ء�˫����Ϲ��Լ�V4���ļ���ģʽ�����Ӿ���������ά�ֽεľ�������.����, �Ӿ���������ά�ֹ����еĸ���ЧӦ�����ڿǺ����ļ���ǿ����.
�������о��Ĵ��µ����������¼�������.����, ���о��Ը��ӿ�����Ϣ���Ӿ���������ά�ֹ������˸����������ʶ, Ϊ�������������Բ�ͬ��Ϣ�ļӹ�ģʽ�ṩ�˻���.����, ���о�̽�����Ӿ�Ƥ���ڼ���ά�ֽ�����ζԲ�ͬ�Ŀ�����Ϣ���б����Լ��Ӿ��������为��ЧӦ����֪����, �Ӿ�Ƥ��ͿǺ������ڸ������е���֪�ӹ����ƽ�Ϊ����һ�������Ӿ���������ά�ֹ����ṩ������.���, ���о�Ϊ�Ӿ����������о������ṩ�˿ɹ��ο�����֪�����֤��.
�������о��IJ���֮������Ľ���������: (1) ��Ҫ����ά�ֽεĻ���, ��δ�о��������ȡ���Ƿ�Ҳ�����ƵĻ���; (2) ������̬�ĽǶ�̽���Ӿ���������ά�ֽε���֪����, ������ά�ֽεIJ���������.���, �ڽ������о������ǿ��Խ��о��ӽ��ؿ������Ӿ������������нε���֪�����Լ����������о���.
�����ο����ף�
����[1] Baddeley A D, Hitch G.Working memory.Psychol Learn Motiv, 1974, 8:47-89
����[2] Luck S J, Vogel E K.The capacity of visual working memory for features and conjunctions.Nature, 1997, 390:279-281
����[3] Vogel E K, Machizawa M G.Neural activity predicts individual differences in visual working memory capacity.Nature, 2004, 428:748-799
����[4] Unsworth N, Fukuda K, Awh E, et al.Working memory and fluid intelligence:Capacity, attention control, and secondary memory retrieval.Cogn Psychol, 2014, 71:1-26
����[5] Tian X.Guidance of attention from the content of working memory in high perceptual load condition (in Chinese) .Doctoral dissertation.Chongqing:Southwest University, 2012[����.���������д洢����Ϣ�ڸ�֪����������������ע��ѡ��.��ʿѧλ����.����:���ϴ�ѧ, 2012]
����[6] Yang H B.Research on the development of visual working memory’s effect on top-down attention control (in Chinese) .Doctoral dissertation.Tianjin:Tianjin Normal University, 2008[���.�Ӿ�������������϶���ע�����Ӱ��ķ�չ�о�.��ʿѧλ����.���:���ʦ����ѧ, 2005]
����[7] Zhou S J, Zhang L Y, Yang J.Development of working memory aattery and its validity in primary school student (in Chinese) .Chin JClin Psychol, 2005, 3:261-264[������, ������, ���.����������ײ���ı��Ƽ���Сѧ���еij���Ч�ȷ���.�й��ٴ�����ѧ��־, 2005, 13:261-264]
����[8] de Fockert J W, Rees G, Frith C D, et al.The role of working memory in visual selective attention.Science, 2001, 291:1803-1806
����[9] Tek C, Gold J, Blaxton T, et al.Visual perceptual and working memory impairments in schizophrenia.Arch Gen Psychiat, 2002, 59:146-161
����[10] Woodman G F, Vecera S P, Luck S J.Perceptual organization influences visual working memory.Psychon B Rev, 2003, 10:80-88
����[11] Bai X J, Zang C L, Wang L H.Reasoning and working memory (in Chinese) .Adv Psychol Sci, 2007, 15:16-21[��ѧ��, 갴���, ������.�����빤������.������ѧ��չ, 2007, 15:16-21]
����[12] Liu H J, Guo D J, Li H L, et al.Relationship among achievement goal orientation, test anxiety and working memory (in Chinese) .Acta Psychol Sin, 2006, 38:254-261[���ݾ�, ���¿�, �����, ��.�ɾ�Ŀ�궨���齹���빤������Ĺ�ϵ.����ѧ��, 2006, 38:254-261]
����[13] Ester E F, Anderson D E, Serences J T, et al.A neural measure of precision in visual working memory.J Cogn Neurosci, 2013, 25:754-761
����[14] Miller G A.The magical number seven plus or minus two:Some limits on our capacity for processing information.Psychol Rev, 1994, 101:343-352
����[15] Lee S H, Baker C I.Multi-voxel decoding and the topography of maintained information during visual working memory.Front Neurosci-SWITZ, 2016, 10:2
����[16] Shen M W, Yi Y J, Zhang F.A study of visual working memory load in n-back paradigm (in Chinese) .Stud Psychol Behav, 2003, (3) :166-170[��ģ��, ������, �ŷ�.n-back�������Ӿ��������为���о�.��������Ϊ�о�, 2003, (3) :166-170]
����[17] Harrison S A, Tong F.Decoding reveals the contents of visual working memory in early visual areas.Nature, 2009, 458:632-635
����[18] Xing Y, Ledgeway T, Mcgraw P V, et al.Decoding working memory of stimulus contrast in early visual cortex.J Neurosci, 2013, 33:10301
����[19] Tong F, Nakayama K, Moscovitch M, et al.Response properties of the human fusiform face area.Cogn Neuropsychol, 2000, 17:257-280
����[20] Kanwisher N, Mc Dermott J, Chun M M.The fusiform face area:A module in human extrastriate cortex specialized for face perception.JNeurosci, 1997, 17:4302-4311
����[21] Grill-Spector K.The neural basis of object perception.Curr Opin Neurobiol, 2003, 13:159-166
����[22] Tong F, Rademaker R, Cohen E.Working memory for complex objects revealed by f MRI decoding of human visual cortical activity.JVison, 2012, 12:1271-1271
����[23] Serences J T, Yantis S.Selective visual attention and perceptual coherence.Trends Cogn Sci, 2006, 10:38-45
����[24] Wang Y, Luo Y J.Standardization and assessment of college students’facial expression of emotion (in Chinese) .Chin J Clin Psychol, 2005, 13:181-192[����, ��Ծ��.��ѧ����ױ�����ϵı�����������.�й��ٴ�����ѧ��־, 2005, 13:181-192]
����[25] Engel S A, Glover G H, Wandell B A.Retinotopic organization in human visual cortex and the spatial precision of functional MRI.Cereb Cortex, 1997, 7:181-192
����[26] Chang C C, Lin C J.LIBSVM:A library for support vector machines.ACM T Intel Syst Tec, 2011, 2:1-27
����[27] Eng H Y, Chen D, Jiang Y.Visual working memory for simple and complex visual stimuli.Psychon B Rev, 2005, 12:1127-1133
����[28] Downing P E.Interactions between visual working memory and selective attention.Psychol Sci, 2000, 11:467-473
����[29] Alvarez G A, Cavanagh P.The capacity of visual short-term memory is set both by visual information load and by number of objects.Psychol Sci, 2004, 15:106-111
����[30] Desimone R, Duncan J.Neural mechanisms of selective visual attention.Annu Rev Neurosci, 1995, 18:193-222
����[31] O’Craven K M, Downing P E, Kanwisher N.f MRI evidence for objects as the units of attentional selection.Nature, 1999, 401:584-587
����[32] Spiridon M, Kanwisher N.How distributed is visual category information in human Occipito-Temporal cortex?Neuron, 2002, 35:1157-1165
����[33] Summerfield C, Egner T, Mangels J, et al.Mistaking a house for a face:Neural correlates of misperception in healthy humans.Cereb Cortex, 2006, 16:500-508
����[34] Ranganath C.Working memory for visual objects:Complementary roles of inferior temporal, medial temporal, and prefrontal cortex.Neuroscience, 2006, 139:277-289
����[35] Ranganath C, Cohen M X, Dam C, et al.Inferior temporal, prefrontal, and hippocampal contributions to visual working memory maintenance and associative memory retrieval.J Neurosci, 2004, 24:3917-3925
����[36] Corbetta M, Kincade M J, Lewis C, et al.Neural basis and recovery of spatial attention deficits in spatial neglect.Nat Neurosci, 2005, 8:1603-1610
����[37] Ranganath C, De Gutis J, D’Esposito M.Category-specific modulation of inferior temporal activity during working memory encoding and maintenance.Cognitive Brain Res, 2004, 20:37-45
����[38] Druzgal T J, D’Esposito M.Activity in fusiform face area modulated as a function of working memory load.Cognitive Brain Res, 2001, 10:355-364
����[39] Druzgal T J, D’esposito M.Dissecting contributions of prefrontal cortex and fusiform face area to face working memory.J Cognitive Neurosci, 2003, 15:771-784
����[40] Leeuwen T V.Color specificity in the human V4 complex-An f MRI repetition suppression study//Tribal livelihood and globalisation:The potential of non-timber forest produce development in central India.Nehru Memorial Museum Library, 2014
����[41] Wang B, Yan T, Ohno S, et al.Retinotopy and attention to the face and house images in the human visual cortex.Expl Brain Res, 2016, 234:1623-1635
����[42] Williams R J, Reutens D C, Hocking J.Functional localization of the human color center by decreased water displacement using diffusion-Weighted f MRI.Brain Behav, 2015, 5:e00408
����[43] Grillspector K, Knouf N, Kanwisher N.The fusiform face area subserves face perception, not generic within-category identification.Nat Neurosci, 2004, 7:555-562
����[44] Sergent J, Ohta S, Macdonald B.Functional neuroanatomy of face and object processing.A positron emission tomography study.Brain, 1992, 115:15-36
����[45] Kanwisher N, Tong F, Nakayama K.The effect of face inversion on the human fusiform face area.Cognition, 1998, 68:B1
����[46] Hoffman E A, Haxby J V.Distinct representations of eye gaze and identity in the distributed human neural system for face perception.Nat Neurosci, 2000, 3:80-84
����[47] Gauthier I, Tarr M J, Moylan J, et al.The fusiform“face area”is part of a network that processes faces at the individual level.J Cognitive Neurosci, 2000, 12:495-504
����[48] Haxby J V, Gobbini M I, Furey M L, et al.Distributed and overlapping representations of faces and objects in ventral temporal cortex.Science, 2001, 293:2425-2430
����[49] Mcnab F, Klingberg T.Prefrontal cortex and basal ganglia control access to working memory.Nat Neurosci, 2008, 11:103-110
����[50] Schendan H E, Searl M M, Melrose R J, et al.An f MRI study of the role of the medial temporal lobe in implicit and explicit sequence learning.Neuron, 2003, 37:1013-1038
����[51] Voytek B, Knight R T.Prefrontal cortex and basal ganglia contributions to visual working memory.Proc Natl Acad Sci USA, 2010, 107:18167-18172
����[52] Hamilton J P, Gotlib I H.Neural substrates of increased memory sensitivity for negative stimuli in major depression.Biol Psychiat, 2008, 63:1155-1162