…˙ŒÔªØ—߬€Œƒ

°°°°’™ “™£∫°°≥˝¡À◊˜Œ™“≈¥´–≈œ¢µƒ‘ÿÃÂ,DNAÀ˘’πœ÷≥ˆµƒÃÿ ‚≤ƒ¡œ–‘ƒÐ“˝∆¡Àπ„∑∫πÿ◊¢°£ª˘”⁄ºÓª˘ª•≤π≈‰∂‘‘≠‘Úµƒæ´»∑–‘∫Õø…±ý≥Ö‘ πµ√∫ÀÀ·ƒ…√◊Ω·ππµƒππΩ®÷≤Ω¥”“ªŒ¨µ•¡¥∑¢’πµΩ∂˛Œ¨∆Ω√Ê“‘º∞»˝Œ¨¡¢ÃÂΩ·ππ°£º∆À„ª˙∏®÷˙π§æþµƒΩ¯≤Ω“≤¥ŸΩ¯¡À∏˜÷÷¥Û–°∫Õ–Œ◊¥µƒDNAƒ…√◊Ω·ππµƒ◊‘∂ت؅˺∆,∂¯Ω¸ƒÍ¿¥ππΩ®µƒ“øÚºÐ∫ÀÀ·(FNAs)”Œ™…˙ŒÔ¥Û∑÷◊”ƒ…√◊≥þ∂»…œµƒæ´»∑≈≈¡–÷π©¡À–¬∑Ω∑®,∆‰πÔ–µƒ…˙ŒÔ—ßπ¶ƒÐ“‘º∞ø…∂®÷∆µƒÃÿ–‘ πµ√∆‰‘⁄ŒÔ¿Ì°¢ªØ—ß∫Õ…˙ŒÔµ»¡Ï”Úæþ”– Æ∑÷π„¿´µƒ”¶”√«∞æ∞°£±æŒƒ≤˚ ˆ¡Àæ´»∑◊‘◊È◊∞µƒFNAsµƒ∏≈ƒÓ,≤¢∏≈ ˆ¡ÀFNAs‘⁄µ∞∞◊æ´»∑◊È◊∞µ»¡Ï”Úµƒ◊Ó–¬Ω¯’π;÷ÿµ„¬€ ˆ¡ÀFNAsµƒ”≈ ∆À˘¥¯¿¥µƒ∂‘µ∞∞◊ø’º‰≈≈≤ºº∞∆‰–‘ƒÐµƒµ˜øÿƒÐ¡¶,Ã÷¬€¡À∏√¡Ï”ڥʑ⁄µƒÃÙ’Ω,≤¢∂‘∆‰∑¢’πª˙”ˆΩ¯––¡À’πÕ˚°£
°°°°πÿº¸¥ £∫°°øÚºÐ∫ÀÀ·; DNAƒ…√◊ºº ı; DNA’€÷Ω; µ∞∞◊æ´»∑◊È◊∞;
°°°°Abstract£∫°°In addition to being the carrier of genetic information, DNA has attracted wide interest as a new class of material in the synthetic realm. The exploitation of the precise and programmable Watson-Crick base pairing of DNA or RNA has led to the development of exquisite nucleic acid nanostructures from one to three dimensions. Advances in computer-aided tools have also facilitated the automated design of DNA nanostructures with various sizes and shapes. The “framework nucleic acids(FNAs)” constructed in recent years provide a new method for precise organization of biological macromolecules with nanometer precision. The intrinsic biological properties and tailorable functionalities of FNAs hold great promise for physical, chemical, and biological applications. This review elaborates the concept of precise self-assembled FNAs, and summarizes the recent advances of FNAs in the field of protein precise arrangement. The unique features of FNAs that benefit the arrangement of proteins and their performance are highlighted. The challenges and opportunities of this field are also discussed.
°°°°Keyword£∫°°Framework Nucleic Acids; DNA nanotechnology; DNA origami; Protein precise arrangement;
°°°°DNAÀ´¬ð–˝Ω·ππ «…˙ŒÔõƒ“≈¥´—ߪ˘¥°[1],≥˝¡À’‚÷÷◊Ó◊≈√˚µƒΩ·ππÕ‚,∫ÀÀ·‘⁄◊‘»ªΩÁ÷–“≤¥Ê‘⁄◊≈–Ì∂ý∆‰À˚µƒππ–Õ,¿˝»Áµ•À´¡¥∫Àëø™πÿ°¢∫À√∏[2]°¢ª∑◊¥RNA[3]“‘º∞Ω·ππ∏¸º”∏¥‘”µƒG-Àƒ¡¥ÃÂ[4]∫Õi-motif[5]µ»(Õº1(a))°£’‚–©ÃÏ»ªµƒÃÿ ‚Ω·ππ“—æ≠±ª÷§√˜‘⁄ªÓœ∏∞˚÷–∆◊≈÷ÿ“™◊˜”√,»Áœ∏∞˚◊™¬ºøÿ÷∆°¢–≈∫≈Õ®¬∑µ˜øÿµ»°£“‘◊‘»ªŒ™¡È∏–,—–æø»À‘±∑¬…˙ππΩ®¡À“ªœµ¡–∏¸º”∏¥‘”∂ý—˘µƒDNAΩ·ππ,∞¸¿®DNAÕþ∆¨[6,7]°¢DNAÀƒ√ÊÃÂ[8]°¢DNA’€÷Ω[9,10]“‘º∞DNA∫Ռު˙ªØ∫œŒÔ‘”Ωª–Œ≥…µƒ«Ú–Œ∫ÀÀ·[11,12]µ»(Õº1(b))°£’‚–©»À‘ϵƒDNAΩ·ππ‘⁄µ˜øÿªÓÃÂ∫Õœ∏∞˚ƒ⁄µƒ…˙ŒÔΩ¯≥Ã∑Ω√Êæþ”–º´¥Ûµƒ«±¡¶°£
°°°°20 ¿ºÕ80ƒÍ¥˙,SeemanÕ®π˝∂®¡øªÏ∫œÀƒ∏ˆæþ”–Ãÿ∂®–Ú¡–µƒ»Àπ§∫œ≥…DNAµ•¡¥¥Ú∆∆¡Àœ∏∞˚ƒ⁄∑«πÃ∂®µƒHollidayΩª≤ʵƒ–Ú¡–∂‘≥∆–‘œÞ÷∆, µœ÷¡ÀπÃ∂®µƒHollidayΩª≤ʵƒÃÂÕ‚◊‘◊È◊∞[6,7]°£Seeman»œŒ™,æþ”–N(N≥3)Ãı ÷±€¡¥µƒDNAΩª≤Ê∂ºƒÐπªÕ®π˝’‚÷÷∑Ω∑® µœ÷◊È◊∞,≤¢«“Õ®π˝…˺∆”√”⁄‘”Ωªµƒ’≥–‘ƒ©∂À,ø…“‘ πæþ”–∂ýÃı ÷±€¡¥µƒDNAΩª≤ÊΩ¯“ª≤Ω◊‘◊È◊∞≥…Œ™∏¥‘”µƒƒ…√◊Ω·ππ[6]°£’‚“ª≥…π˚ πDNA≥…Œ™¡À≤ªΩˆœÞ”⁄“ªŒ¨µƒƒ…√◊º∂æ€∫œŒÔ,ª˘”⁄’‚“ª…ËœÎ,ø∆—ߺ“…˺∆≤¢Ω®¡¢¡À‘Ω¿¥‘Ω∏¥‘”∂ý—˘°¢∆ı∫œ”¶”√µƒDNAƒ…√◊Ω·ππ°£

°°°°DNAƒ…√◊Ω·ππµƒø…±ý≥Ö‘∫Õƒ…√◊æ´◊º–‘“‘º∞Ω·ππ∑·∏ª–‘µ»–‘÷ π∆‰ƒÐπª◊˜Œ™ø…øÿ∏’–‘÷ߺБ⁄ƒ…√◊≥þ∂»æ´»∑≈≈¡–∂ý∏ˆ∑÷◊”,¥”∂¯ππΩ®æþ”–º∂¡™–ß”¶µƒ∑÷◊”Õ¯¬Á°£Õ¨ ±,DNAƒ…√◊Ω·ππ”…”⁄∆‰≤ªÕ¨Œªµ„µƒ–Ú¡–Ãÿ“Ï–‘,ø…“‘◊˜Œ™ƒ…√◊≥þ∂»µƒø’º‰ø…—∞÷∑–‘÷ߺÐ,‘⁄’‚÷÷«Èøˆœ¬,ƒÐπª µœ÷…˙ŒÔ∑÷◊”º‰æý¿Î∫Õœý∂‘±»¬ µƒæ´»∑µ˜øÿ[13,14]°£’‚∆‰÷–,æþ”–Ãÿ ‚ŒÔ¿ÌªØ—ßÃÿ–‘µƒ“øÚºÐ∫ÀÀ·(Framework nuclear acids,FNAs)”ƒ…√◊Ω·ππŒ™Ãƒ⁄∫ÕÃÂÕ‚∑÷◊”◊È◊∞÷π©¡À–¬µƒÀº¬∑[15,16,17,18,19]°£FNAsÃÿ ‚µƒ¿ÌªØ–‘÷ ÷˜“™ÃÂœ÷‘⁄“‘œ¬∑Ω√Ê:µ⁄“ª,FNAs±æ…Ì «∏þ∂»µ•∑÷…¢ƒ…√◊Ω·ππ,”Îæ´∂»Ωœ≤ÓµƒŒÞª˙∫Õ”–ª˙ƒ…√◊ø≈¡£œý±»,∆‰æþ”–Ω”Ω¸‘≠◊”º∂±µƒæ´»∑∂»;µ⁄∂˛,FNAsƒÐπª µœ÷–°∑÷◊”°¢¥Û∑÷◊”ªÚ «ƒ…√◊ø≈¡£‘⁄ƒ…√◊≥þ∂»…œµƒæ´»∑Œªµ„µ˜øÿ[20,21,22,23,24];µ⁄»˝,FNAs∫Õµ•À´¡¥DNA‘⁄œ∏∞˚ƒ⁄ªØ∫Õ∑÷≤º∑Ω√Ê”–◊≈œ‘◊≈µƒ≤Ó“Ï[25,26,27,28,29,30]°£“‘FNAsµƒª˙–µ–‘ƒÐŒ™¿˝,FNAsµƒ∏’–‘ «∏þ∂»ø…øÿ∫Õø…±ý≥õƒ°£”…”⁄æ€∫œŒÔµƒ—”…Ï≥§∂»”Î∞Îæ∂µƒ4¥Œ∑Ω≥…’˝±»,”…∞Îæ∂Œ™10°´100 nmµƒFNAsÀ˘ππΩ®µƒDNAƒ…√◊Ω·ππ‘⁄—”…Ï≥§∂»…œ≥¨π˝µ•À´¡¥DNA 4°´6∏ˆ ˝¡øº∂°£“Ú¥À,»·–‘µƒDNA ÷±€¡¥ΩȵºµƒŒÞ∑Ï‘”Ωª∂¯ªÒµ√µƒ∏’–‘FNA÷ߺЌ™ƒ…√◊≥þ∂»Ω·ππµƒππΩ®Ã·π©¡À√˜»∑µƒÀº¬∑°£
°°°°Õº1 ÷÷¿ý∑·∏ªµƒ∫ÀÀ·Ω·ππ
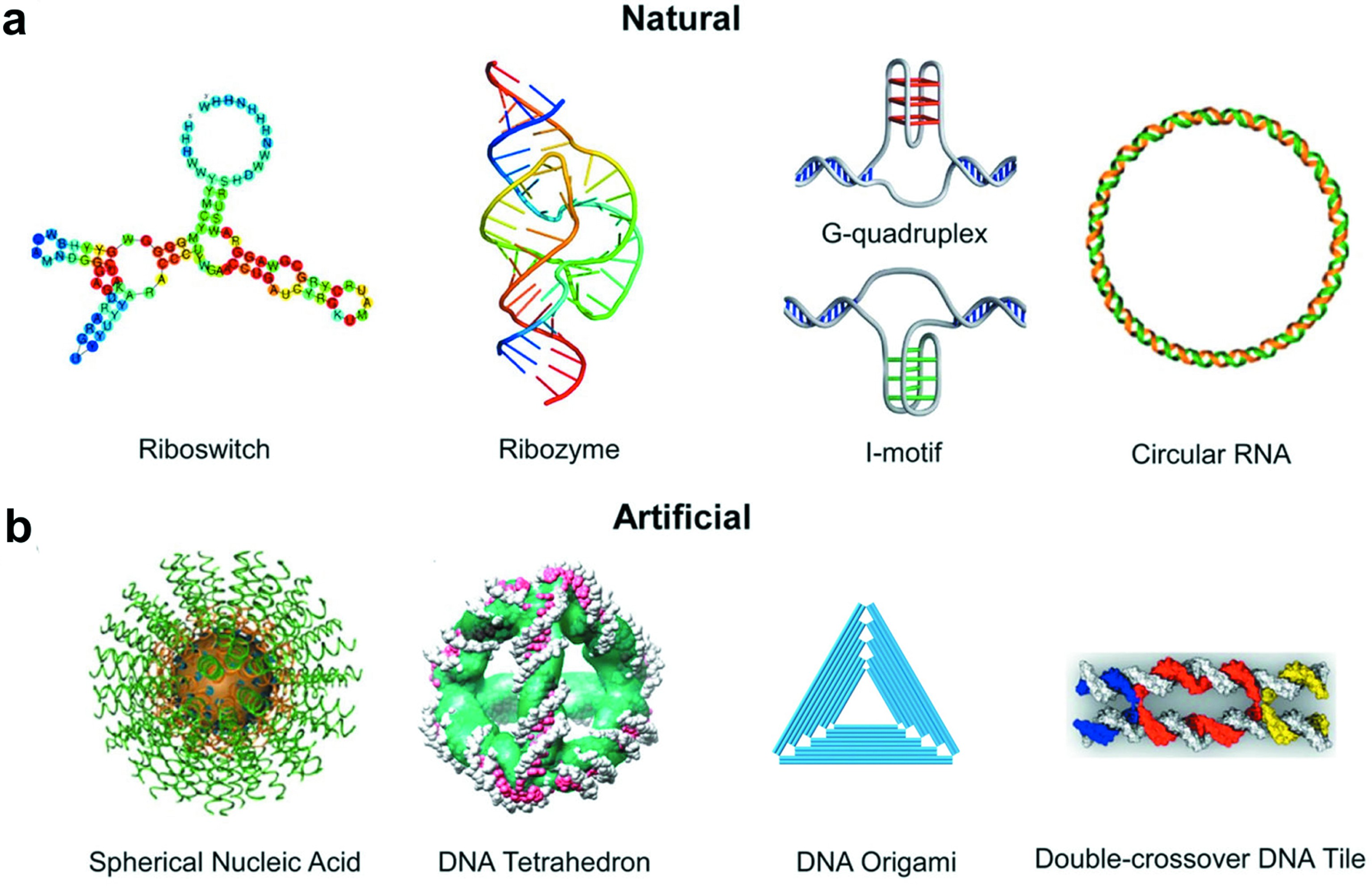
°°°°Fig.1 A wide variety of nucleic acid structures
°°°°(a)◊‘»ªΩÁ÷–¥Ê‘⁄µƒ∫ÀÀ·Ω·ππ[2°´5];(b)»Àπ§…˺∆µƒ∫ÀÀ·Ω·ππ[6,8,9,11]
°°°°µΩƒø«∞Œ™÷π,ø∆—ߺ““—æ≠—–æø¡ÀFNAsΩȵºµƒæý¿Î“¿¿µ–‘∑÷◊”œýª•◊˜”√°¢µ◊ŒÔÕ®µ¿ªØ∫Õ«¯ “ªØ–ß”¶µ»°£¥ÀÕ‚FNAs“—æ≠±ª”√”⁄ππΩ®¥”ÃÂÕ‚–°∑÷◊”…˙ŒÔ¥´∏–∆˜µΩ∏¥‘”µƒÃƒ⁄÷Œ¡∆–‘“©ŒÔµðÀÕœµÕ≥“‘º∞∂ý√∏Õ¯¬Áµ»∂ý÷÷ƒ…√◊∆˜º˛[13]°£±æŒƒ∑÷Œˆ¡À…˙ŒÔ∑÷◊”ø’º‰≈≈¡–∂‘∆‰π¶ƒÐµƒ”∞œÏ,≤¢Ω¯“ª≤ΩÃΩÃ÷¡À≤ªÕ¨Œ¨∂»µƒFNAs‘⁄∂ý΃∫Õµ∞∞◊µƒæ´»∑◊È◊∞∑Ω√ʵƒ◊Ó–¬—–æøΩ¯’π°£
°°°°1°¢“ªŒ¨œþ–‘øÚºÐ∫ÀÀ·
°°°°ÀÊ◊≈DNAªØ—ß∫œ≥…µƒ÷Ω•≥… Ï,∞¸¿®”´π‚»æ¡œ°¢¡Ú¥º°¢…˙ŒÔÀÿµ»‘⁄ƒ⁄µƒ∂ý÷÷ªØ—ߪ˘Õ≈º∏∫ıø…“‘”ÎFNAsµƒ◊È∑÷¡¥‘⁄»Œ∫ŒŒªµ„ µœ÷π≤º€¡¨Ω”[31],–°∑÷◊”πŸƒÐÕ≈µƒπ≤º€–Þ Œ“≤≥…Œ™¡Àµ∞∞◊”Î∫ÀÀ·Ω·∫œµƒª˘¥°,FNAsµƒæ´»∑◊‘◊È◊∞Ãÿ–‘‘Úø…“‘ µœ÷∂‘’‚–©ªÓ–‘∑÷◊”µƒæ´»∑ø’º‰µ˜øÿ°£“ªŒ¨œþ–‘∫ÀÀ·◊˜Œ™◊ÓºÚµ•µƒøÚºÐΩ·ππ”Î∏˜¿ýπ≤º€Ω·∫œ–Þ Œºº ıÕ˘Õ˘±ª”√”⁄µ˜øÿÀ´µ∞∞◊÷ƺ‰µƒø’º‰∑÷≤º¥”∂¯—–æøµ∞∞◊–≠Õ¨◊˜”√∂‘∆‰π¶ƒÐµƒ”∞œÏ°£Freemanµ»[32]µƒ—–æø±Ì√˜,DNAÀ´¡¥ƒÐπªµ˜Ω⁄∂ý΃÷ƺ‰µƒº‰æý, µœ÷∂‘∂ý΃–≠Õ¨◊˜”√µƒµ˜øÿ°£∂ý΃RGDS∫ÕPHSRN «œÀŒ¨¡¨Ω”µ∞∞◊µƒ◊È∑÷,À¸√«–≠Õ¨µ˜øÿœ∏∞˚µƒ§∏Ω––Œ™°£“‘DNAÀ´¡¥◊˜÷ߺÐ,À˚√«”≈ªØ¡ÀRGDS∫ÕPHSRN÷ƺ‰µƒæý¿Î,÷∏þ¡À∂ý΃–≠Õ¨◊˜”√µƒ–߬ ,”Î∂‘’’◊Èœý±»,œ∏∞˚§∏Ω–ß¬ ÷∏þ¡À46%(Õº2(a))°£
°°°°≥˝¡À∂‘≤ªÕ¨µ∞∞◊–≠Õ¨◊˜”√µƒø’º‰µ˜øÿ,“ªŒ¨FNAs“≤”√”⁄ππΩ®∑¬…˙µƒ»Àπ§µ∞∞◊°£“‘FNAs◊˜Œ™π«ºÐ, µœ÷¡Àµ∞∞◊π¶ƒÐ—«ª˘µƒ◊‘◊È◊∞°£Niemeyerµ»[33]¿˚”√FNAsæý¿Îµ˜øÿ µœ÷¡Àœ∏∞˚…´ÀÿP450 BM3—«ª˘µƒ◊È◊∞,BM3∫¨”–¡Ω∏ˆ—«ª˘,º¥ªπ‘≠√∏—«ª˘BMR∫Õþ≤þ¯—«ª˘BMP,¡Ω∏ˆ—«ª˘∑÷±”ÎHaloTagµ∞∞◊»⁄∫œ,HaloTagµ∞∞◊∑÷±”ά»ÕÈÃ˛–Þ ŒµƒDNA≈º¡™,µ±DNAÕ®π˝ºÓª˘ª•≤π≈‰∂‘¡¨Ω”‘⁄“ª∆ ±,‘⁄Ãÿ∂®µƒº‰æýœ¬,¡Ω∏ˆµ∞∞◊—«ª˘ƒÐπª µœ÷÷ÿ◊È,≤¢«“±£¡Ù¡ÀBM3µƒ√∏ªÓ–‘(Õº2(b))°£
°°°°Õº2 “ªŒ¨œþ–‘øÚºÐ∫ÀÀ·
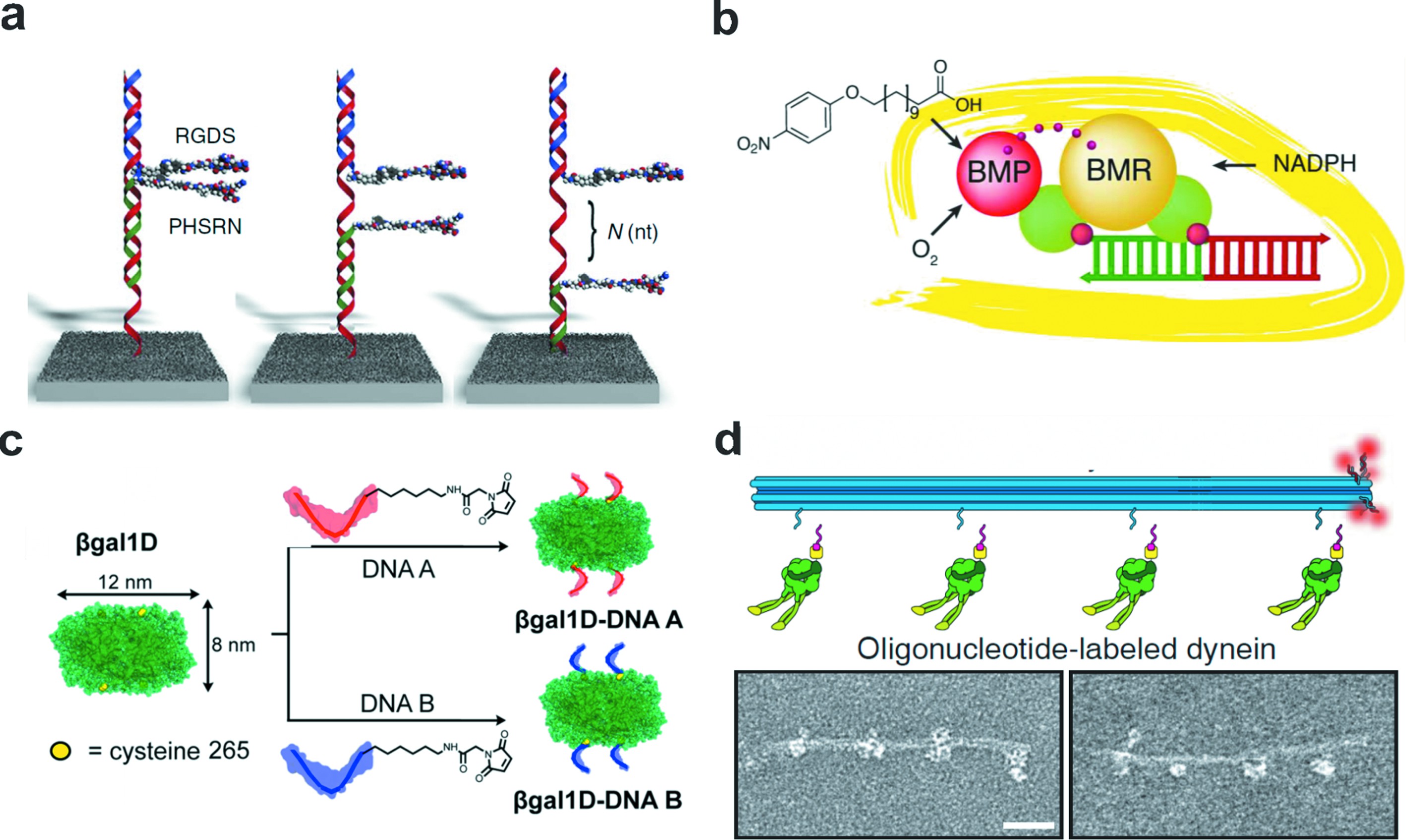
°°°°Fig.2 1D framework nucleic acids
°°°°(a)¿˚”√øÚºÐ∫ÀÀ·µ˜’˚∂ý΃RGDS∫ÕPHSRNµƒº‰æý∂¯µ˜øÿÀ¸√«÷ƺ‰µƒ–≠Õ¨◊˜”√[32];(b)‘⁄øÚºÐ∫ÀÀ·÷ߺЅœ◊È◊∞BMRªπ‘≠√∏Ω·ππ”Ú∫ÕBMPþ≤þ¯Ω·ππ”Ú£¨ÃΩæøœ∏∞˚…´ÀÿP450 BM3µƒæý¿Î“¿¿µ–‘—«ª˘◊‘◊È◊∞[33];(c)¿˚”√øÚºÐ∫ÀÀ· µœ÷β-∞λÈëВ√∏µƒ”––Ú◊‘◊È◊∞[34];(d)øÚºÐ∫ÀÀ·…œ◊È◊∞¬Ì¥Ôµ∞∞◊∑¬…˙ƒ£ƒ‚∆‰…˙ŒÔ—ßπ¶ƒÐ[35]
°°°°(a) FNAs were used to tune the distance between the peptides RGDS and PHSRN for study of their synergy[32];(b) Design of distance-dependence study of cytochrome P450 BM3 subunit self-assembly with organizing the BMR reductase domain and the BMP porphyrin domain on FNA[33];(c) Controllable self-assembly ofβ-galactosidase using FNA[34];(d) An ensemble of motor proteins on FNA for biomimicking of biological functions[35]
°°°°FNAsµƒ∏þ–ß◊‘◊È◊∞ πµ√∆‰‘⁄µ∞∞◊÷ π§≥áϔړ≤”–◊≈”¶”√º€÷µ°£“¿Õ–“ªŒ¨FNAsµƒœþ–ŒΩ·ππ,McMillanµ»[34] π”√∂‘≥∆Àƒæ€Ãµ∞∞◊÷ β-∞λÈëВ√∏◊˜Œ™ƒ£–Õµ∞∞◊,Ω´265∫≈ŒªµƒÀ’∞±À·Õª±‰≥…∞ÎÎ◊∞±À·,”√”⁄‘⁄µ∞∞◊∂•≤ø∫Õµ◊≤ø∑÷±Ωª¡™¬Ì¿¥ı£—«∞∑–Þ ŒµƒDNA,¿˚”√DNAΩȵºµƒ∑ΩœÚøÿ÷∆¥Ÿ πβ-∞λÈëВ√∏÷·œÚ≈≈¡–≥…“ªŒ¨DNA-µ∞∞◊÷ ƒ…√◊œþ,∂¯≤ª «∑«Ãÿ“Ï–‘怺Ø(Õº2(c))°£’‚÷÷FNAsΩȵºµƒµ∞∞◊”––Ú怺ؓ≤±ª”¶”√”⁄ππΩ®∑¬…˙µƒ…˙ŒÔ∑¥”¶ÃÂœµ,Ω¯∂¯—–æø…˙ŒÔÃ⁄µƒ∑¥”¶Ω¯≥Ã,¿˝»Á¬Ì¥Ôµ∞∞◊µƒ…˙ŒÔ—ßπ¶ƒÐ°£—–æø’þ‘⁄FNAs÷ߺЅœ◊È◊∞∂ý÷÷¬Ì¥Ôµ∞∞◊∑÷◊”¿¥—–æøÀ¸√«÷ƺ‰µƒœýª•◊˜”√,Derrµ»[35]¿˚”√DNA’€÷Ω—–æø¡À∂Ø¡¶µ∞∞◊∫Õ«˝∂ص∞∞◊‘⁄œ∏∞˚Œ¢πÐ…œ‘ÀÀÕªıŒÔµƒπϺ£,‘⁄ Æ∂˛¬ð–˝ ¯DNA’€÷Ω…œ◊È◊∞¡Àº´–‘œý∑¥µƒ∂Ø¡¶µ∞∞◊∫Õ«˝∂ص∞∞◊,∑¬…˙ƒ£ƒ‚¡À’‚¡Ω÷÷µ∞∞◊‘⁄œ∏∞˚Œ¢πÐ…œµƒ“∞Œ∫””––Œ™(Õº2(d))°£ªπ”–—–æø’þø™∑¢¡Àª˘”⁄¬Ì¥Ôµ∞∞◊∫ÕDNA∏¥∫œŒÔµƒ»Àπ§◊‘◊È◊∞‘À ‰œµÕ≥,‘⁄Ãÿ∂®∑÷◊”µƒ”’µºœ¬,∏√œµÕ≥ƒÐπª µœ÷◊È◊∞∫Õ≤–∂“‘º∞‘⁄∑÷◊”πϵ¿…œ◊∞–∂ªıŒÔ°£
°°°°2°¢ ∂˛Œ¨∆Ω√ÊøÚºÐ∫ÀÀ·
°°°°ÃÏ»ª¥Ê‘⁄µƒœ∏∞˚Õ®¬∑“‘º∞º∂¡™∑¥”¶¥Û∂ý–Ë“™∂ý∏ˆµ∞∞◊≤Œ”Î,“ªŒ¨œþ–‘µƒFNAs±æ…Ì–Þ ŒŒªµ„µƒÿ—∑¶œÞ÷∆¡À∂ýµ∞∞◊–≠Õ¨“‘º∞µ∞∞◊º∂¡™∑¥”¶µƒ—–æø°£ª˘”⁄DNAø…±ý≥Ö‘,ø∆—ߺ“…˺∆¡À∏¸Œ™∑·∏ª°¢Œ¨∂»∏¸∏þµƒDNAƒ…√◊Ω·ππ°£∂˛Œ¨FNAs”…”⁄∆‰Ãÿ ‚µƒ∆Ω√ÊΩ·ππ“‘º∞±Ì√Ê’≥–‘ƒ©∂Àµƒø……˺∆–‘, µœ÷¡À∆Ω√ʜޔÚ√∏º∂¡™ÃÂœµµƒæ´»∑◊È◊∞,ª˘”⁄¥À∂‘∂ýµ∞∞◊–≠Õ¨◊˜”√Ω¯––—–æø,≤¢Õ®π˝∑÷◊”ø™πÿ µœ÷¡Àµ∞∞◊º∂¡™∑¥”¶µƒ»Àπ§∏…‘§°£ª˘”⁄FNAsµƒæ´»∑◊‘◊È◊∞∫Õø…—∞÷∑–‘,Wilnerµ»[36]¬ œ»‘⁄∂˛Œ¨¡˘Ω«–Œ÷ߺЅœ µœ÷¡À∆œÃ—ë—ıªØ√∏(GOX)/¿±∏˘π˝—ıªØŒÔ√∏(HRP)µ∞∞◊ƒ…√◊æ´∂»µƒ◊‘◊È◊∞,“‘∂˛Œ¨¡˘Ω«–ŒDNA’Û¡–◊˜Œ™÷ߺÐ,Õ®π˝∏ƒ±‰ÃΩ’Î¡¥µƒŒª÷√¿¥ µœ÷µ∞∞◊ø’º‰∑÷≤ºµƒ»Àπ§µ˜øÿ°£”…”⁄¡˘±þ–ŒDNAøںеƒæý¿Îµ˜øÿ”αÌ√ʜޔÚ◊˜”√,∑¥”¶÷–º‰≤˙ŒÔπ˝—ıªØ«‚µƒ¿©…¢–߬ ∏¸∏þ,“Ú∂¯ µœ÷∏¸∏þµƒº∂¡™–߬ °£“Ú¥À,“‘¡Ω∏ˆ¡˘±þ–ŒDNAøÚºÐ◊˜Œ™÷ߺР±µƒ√∏ªÓ–‘±»Àƒ∏ˆ¡˘±þ–ŒDNAøÚºÐ∏þ≥ˆ20%(Õº3(a))°£”–»§µƒ «,’‚÷÷æý¿Îæˆ∂®µƒº∂¡™–߬ »¥≤ª «ÕÍ»´ÀÊæý¿Î‘ˆº”∂¯÷Ω•ΩµµÕµƒ°£ª˘”⁄DNAƒ…√◊≤ƒ¡œµƒ∏þƒ…√◊æ´∂»,ø∆—ߺ“¿˚”√DNA∆Ω√ÊΩ·ππΩ¯––¡À∏¸æ´»∑µƒæý¿Îøÿ÷∆,œµÕ≥µÿ—–æø¡À√∏º‰æý∂‘º∂¡™–߬ µƒ”∞œÏ°£ µ—ÈΩ·π˚±Ì√˜,‘⁄‘ˆº”√∏º‰æý ±,º∂¡™ªÓ–‘≤¢Œ¥÷Ω•ΩµµÕ,∂¯ «‘⁄√∏º‰æýŒ™10nm ±≥ˆœ÷¡À–߬ ÷∏þµƒœ÷œÛ,’‚ «”…”⁄µ∞∞◊œýª•◊˜”√ ±–Œ≥…µƒÀÆ∫Õ≤„¥ŸΩ¯∑¥”¶÷–º‰ÃÂπ˝—ıªØ«‚µƒ¿©…¢µº÷¬µƒ°£
°°°°Yanµ»[37]Õ®π˝‘⁄GOX∫ÕHRP÷ƺ‰ππΩ®≤ªÕ¨¥Û–°µƒµ∞∞◊÷ «≈—È÷§¡À’‚∏ˆª˙÷∆,«≈¡¨µ∞∞◊Ω´∑¥”¶÷–º‰Ãµƒ≤º¿ ‘À∂Ø◊™±‰≥…“¿¿µµ∞∞◊º‰–Œ≥…µƒÀÆ∫Õ≤„Ω¯––¿©…¢(Õº3(b))°£»ª∂¯,≥˝¡ÀGOX-HRP√∏º∂¡™ÃÂœµ“‘Õ‚µƒ∆‰À˚√∏º∂¡™∑¥”¶,…ʺ∞µΩπ˝—ıªØ«‚“‘Õ‚µƒ∆‰À˚∑¥”¶÷–º‰Ãªπ–Ë“™Ω¯––Ω¯“ª≤Ωµƒ—–æø,¥”∂¯≥‰∑÷¡ÀΩ‚√∏º∂¡™∑¥”¶µƒæý¿Î“¿¿µ–‘°£Ã·∏þµ◊ŒÔÕ®µ¿µƒ÷–º‰ÃÂ¥´µð–߬ “≤”–÷˙”⁄÷∏þ√∏º∂¡™–߬ ,¥”±æ÷ …œ¿¥Ω≤,µ◊ŒÔÕ®µ¿”√”⁄Ω´÷–º‰ÃÂ¥”“ª÷÷√∏◊™“∆µΩ¡Ì“ª÷÷√∏,“‘¥ŸΩ¯√∏º∂¡™∑¥”¶°£Yanµ»[38]¿˚”√»Àπ§∞⁄±€∑¬…˙ππΩ®¡À»Àπ§µ◊ŒÔÕ®µ¿,Ω®¡¢¡À“ª÷÷ª˘”⁄FNAsµƒ∂ý√∏∏¥∫œŒÔ,‘⁄¡Ω∏ˆÀ´¡¥∏’–‘πÃ∂®µƒÕ—«‚√∏∆œÃ—ë-6-¡◊À·Õ—«‚√∏(G6pDH)∫Õ∆ªπ˚À·Õ—«‚√∏(MDH)÷ƺ‰–Þ Œ¡À“ªÃı∫¨20Tµƒ»·–‘»Àπ§∞⁄±€‘⁄¡Ω÷÷√∏÷ƺ‰∞⁄∂Ø,¥”∂¯ µœ÷µ◊ŒÔ∑÷◊”‘⁄√∏º∂¡™ÃÂœµ÷–∂ý∏ˆªÓ–‘Œªµ„÷ƺ‰÷±Ω”◊™“∆(Õº3(c))°£¿˚”√FNAs÷ߺÐ∂‘∑¥”¶µ◊ŒÔ±Ì√ʜޔÚ, µœ÷¡À√∏º∂¡™ÃÂœµµ•∏ˆ◊È∑÷µƒæ´»∑Œªµ„øÿ÷∆°£ µ—ÈΩ·π˚±Ì√˜,”ΔŒ¿ÎÃÂœµœý±»,ª˘”⁄FNAsµƒ»Àπ§√∏º∂¡™ÃÂœµµƒ∑¥”¶ªÓ–‘÷∏þ¡À¥Û‘º90±∂,≤¢«“Õ®π˝∏ƒ±‰¡Ω÷÷√∏µƒœý∂‘±»¿˝ƒÐπªΩ¯“ª≤Ω÷∏þ∑¥”¶ªÓ–‘°£≥˝¡Àµ˜øÿ√∏º∂¡™∑¥”¶ªÓ–‘Õ‚,FNAsµƒø…±ý≥Ö‘∫Õ…˙ŒÔœý»ð–‘ πµ√∆‰‘⁄∑¬…˙¡Ï”Ú“≤”–◊≈”¶”√º€÷µ°£Nielsenµ»[39]¿˚”√–«–ŒFNAs”ÎøπÃÂÃÿ∂®Œªµ„π≤º€Ω·∫œ,‘⁄≤ª”∞œÏøπÃÂπ¶ƒÐµƒ«Èøˆœ¬∑¬…˙ππΩ®øπõ∞∞◊ŒÂæ€ÃÂ,ƒ£ƒ‚IgMµƒΩ·ππ“‘º∞∆‰…˙ŒÔ—ßπ¶ƒÐ(Õº3(d))°£
°°°°Õº3 ∂˛Œ¨∆Ω√ÊFNAs
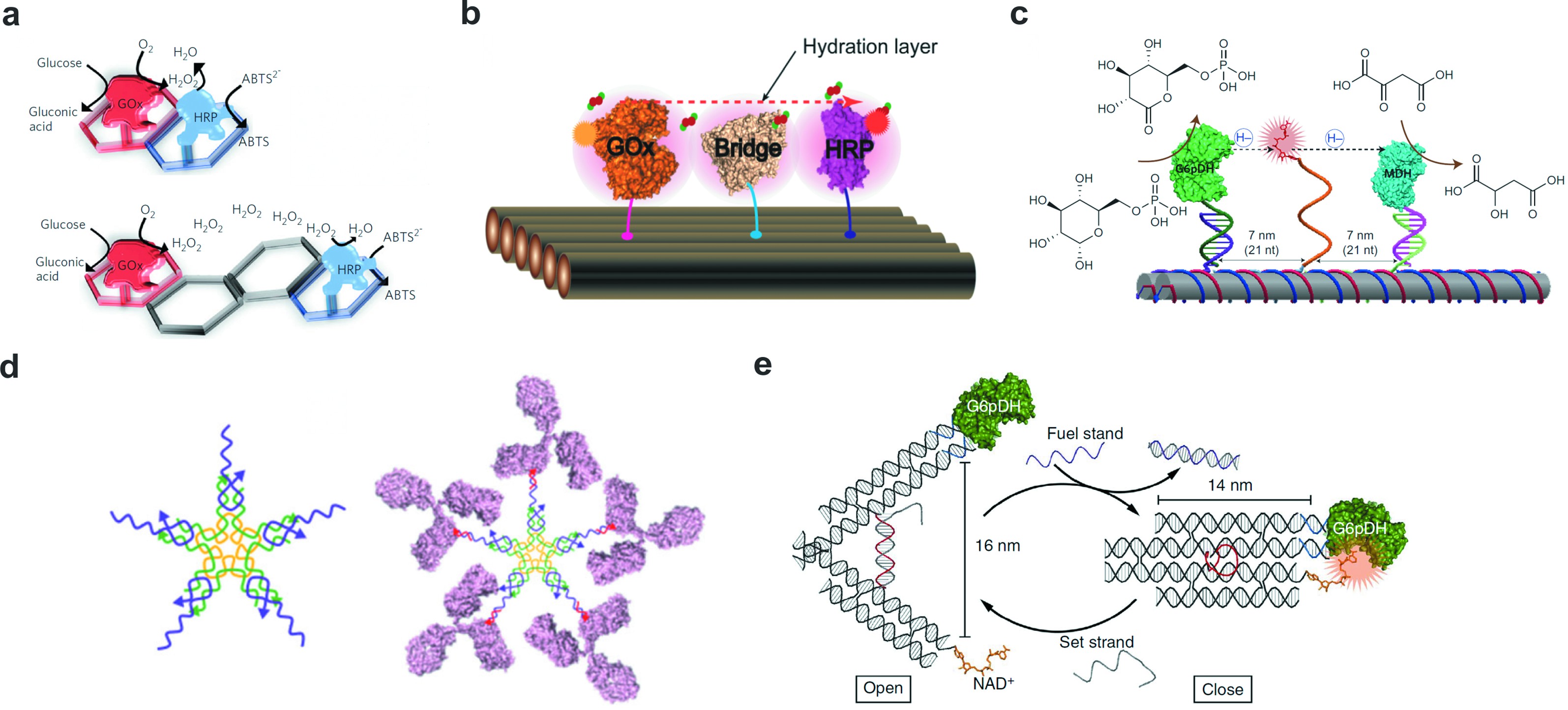
°°°°Fig.3 2D framework nucleic acids
°°°°(a)∆œÃ—ë—ıªØ√∏(GOx)∫Õ¿±∏˘π˝—ıªØŒÔ√∏(HRP)◊È◊∞‘⁄¡˘±þ–ŒFNAs…œ£¨¿˚”√¡˘±þ–Œµƒ∏ˆ ˝µ˜Ω⁄µ∞∞◊÷ƺ‰µƒº‰æý[36];(b) FNAs±Ì√ÊππΩ®GOx/HRP√∏º∂¡™ÃÂœµ£¨µ∞∞◊÷ «≈◊˜Œ™ÀÆ∫œ≤„¥ŸΩ¯π˝—ıªØ«‚¿©…¢[37];(c) FNAs±Ì√ÊππΩ®∆œÃ—ë¡◊À·Õ—«‚√∏(G6p DH)∫Õ∆ªπ˚À·Õ—«‚√∏(MDH)µƒ∂ý√∏∏¥∫œŒÔ£¨≤¢¿˚”√œŸý—þ ∂˛∫ÀÐ’À·(NAD+)–Þ Œµƒµ•¡¥DNA◊˜Œ™∞⁄∂ر€¥ŸΩ¯∑¥”¶÷–º‰≤˙ŒÔµƒ◊™“∆[38];(d) FNAs”√”⁄∑¬…˙ππΩ®√‚“þ«Úµ∞∞◊Ig M[39];(e) FNAsππΩ®√∏¥þªØ∑¥”¶∑÷◊”ø™πÿ[41]
°°°°(a) Scheme of glucose oxidase (GOx) and horseradish peroxidase (HRP) cascade organized with two different distances between GOxand HRP on two-hexagon and four-hexagon strips[36];(b) Scheme of the assembled GOx/HRP pair with a protein bridge working as a connected hydration layer to facilitate H2O2diffusion[37];(c) An artificial swinging-arm multienzyme complex consisting of glucose-6-phosphate dehydrogenase (G6p DH) and malic dehydrogenase(MDH) organized on FNA,with a nicotinamide adenine dinucleotide (NAD+)-modified ss DNA as the swinging arm,facilitating the transfer of hydrides[38];(d) Bio mimic construction of immunoglobulin Ig M using FNA[39];(e) FNA constructs enzyme catalytic reaction molecular switch[41]
°°°°ª˘”⁄µ∞∞◊ø’º‰∑÷≤ºœµÕ≥—–æø,ø∆—ߺ““≤Ω®¡¢¡À“ªœµ¡–µƒ∑÷◊”ø™πÿ,∂‘√∏º∂¡™∑¥”¶Ω¯––»Àπ§∏…‘§,∂¯œýΩœ”⁄“ªŒ¨œþ–ŒΩ·ππ,∂˛Œ¨∆Ω√ÊΩ·ππµƒœÞ”Ú◊˜”√ µœ÷¡Àº∂¡™∑¥”¶µƒø…ƒÊ∏…‘§°£◊˜Œ™“ª∏ˆæ≠µ‰µƒ∂˛Œ¨∑÷◊”ø™πÿ,DNAƒ˜◊”±ªπ„∑∫”√”⁄◊È∑÷÷ƺ‰µƒæý¿Îµ˜øÿ¥”∂¯ µœ÷µ∞∞◊º∂¡™∑¥”¶µƒº§ªÓ°£ƒ˜◊”Õ®π˝DNAÀ´¡¥¡¨Ω”,‘⁄º”»Î»°¥˙¡¥÷Æ∫Û–Œ≥…Œ»∂®µƒæ•ª∑Ω·ππ,ª˘”⁄¥À,—–æø’þ√«ÃΩæø¡À≤ªÕ¨◊¥Ã¨œ¬GOX/HRPº∂¡™œµÕ≥µƒªÓ–‘°£Ω·π˚±Ì√˜,DNAƒ˜◊”µƒ¡Ω∏ˆ±€‘⁄¥Úø™∫Õπÿ±’µƒ◊¥Ã¨œ¬√∏º∂¡™ªÓ–‘œý≤Ó‘º2±∂[40]°£∂¯”…”⁄DNA∑÷◊”ø™πÿµƒø…ƒÊ–‘,DNAƒ˜◊”ø…“‘‘⁄¥Úø™∫Õπÿ±’◊¥Ã¨÷ƺ‰«–ªª÷¡…Ÿ4∏ˆ÷Ð∆⁄,“Ú¥ÀDNAƒ˜◊”“≤±ª”¶”√”⁄øÿ÷∆√∏º∞∆‰∏®√∏µƒΩ·∫œ∫ÕΩ‚¿Î°£Yanµ»[41]Ω´G6pDH∫Õ—Ãı£∞∑œŸý—þ ∂˛∫ÀÐ’À·(NAD+)∑÷±¡¨Ω”µΩDNAƒ˜◊”µƒ¡Ω∏ˆ±€…œ,”…”⁄√∏Ωˆ‘⁄÷±Ω”“Ω”¥•”∏®√∏ ±≤≈æþ”–ªÓ–‘,“Ú¥À”Îø™∑≈◊¥Ã¨œý±»,±’∫œ◊¥Ã¨œ¬µƒ√∏ªÓ–‘÷∏þ¡À‘º5±∂,≤¢«“’‚∏ˆø™πÿ◊¥Ã¨ «ø…“‘Õ®π˝Ã̺”÷√ªª¡¥ µœ÷ø…ƒÊµ˜Ω⁄µƒ(Õº3(e))°£
°°°°3°¢ »˝Œ¨¡¢ÃÂøÚºÐ∫ÀÀ·
°°°°”…”⁄“ªŒ¨°¢∂˛Œ¨FNAsµ∞∞◊–Þ Œæ˘±©¬∂‘⁄Ω·ππ±Ì√Ê, πµ√∆‰ƒ—“‘ µœ÷µ∞∞◊¥ÿø’º‰œÞ”Ú—–æø°£»˝Œ¨FNAs”…”⁄∆‰¡¢ÃÂΩ·ππƒÐπªæ´»∑µÿ µœ÷∑÷◊”µƒø’º‰≈≈≤º,√÷≤π¡À“ªŒ¨ªÚ∂˛Œ¨FNAsµÕŒ¨∂»µƒ≤ª◊„°£»˝Œ¨ƒ…√◊Ω·ππΩȵºµƒ¡˝◊”◊¥FNAsƒÐ𪃣ƒ‚ÃÏ»ª«¯ “ππΩ®ƒ…√◊∑¥”¶∆˜,»ª∫ÛΩ´¥˙–ª√∏”––Úµƒ∑‚◊∞Ω¯»•,¥”∂¯‘ˆ«ø¥þªØªÓ–‘≤¢Ã·∏þµ∞∞◊√∏œ˚ªØÃıº˛œ¬µƒŒ»∂®–‘(Õº4(a))[42]°£
°°°°¿˚”√»˝Œ¨DNAæßÃÂππΩ®µ∞∞◊Õ¯¬Á“≤ «“ªº˛ Æ∑÷”–»§µƒ ¬«È,◊‘¥”µ⁄“ª∏ˆø…◊‘Œ“◊È◊∞µƒDNAæßÃÂΩ·ππ±ª…Ëº∆≥ˆ¿¥,—–æø»À‘±≤ª∂œ≥¢ ‘∂‘DNAæßÃÂΩ¯––π¶ƒÐªØ(Õº4(b))[43]°£ π”√DNAæß൜÷µ∞∞◊µƒø’º‰æ´»∑µ˜øÿ «∑«≥£”–“‚“µƒ,”ε•∑÷◊”œµÕ≥œý±»,—–æø»˝Œ¨Õ¯¬Á»Á∫Œ”∞œÏµ∞∞◊π¶ƒÐ”–¿˚”⁄∏¸º”…ӻΡÀΩ‚Ãƒ⁄œýπÿ…˙ŒÔ—ßπ¶ƒÐ°£
°°°°»˝Œ¨¡¢ÃÂFNAsµƒø’º‰Ω·ππ πµ√∆‰ƒÐπªΩ´∑÷◊”∑‚◊∞∆¿¥,±Ð√‚∑÷◊” еΩ÷Ќߪ∑æ≥µƒ”∞œÏ,“Ú¥Àª˘”⁄»˝Œ¨FNAsµƒƒ…√◊‘ÿ“©œµÕ≥‘⁄º≤≤°’Ô¡∆¡Ï”Ú“≤”–◊≈∆‰”¶”√º€÷µ°£¥´Õ≥µƒ“©ŒÔ‘ÿêÚ∂ýªÚ…Ÿµÿ¥Ê‘⁄“ª–©πÔ–µƒæ÷œÞ–‘,¿˝»Á,÷¨÷ Ô…”⁄∆‰≥þ¥Á≤ªæ˘“ª∂¯ø…ƒÐ”’µº√‚“þ∂æ–‘°¢ŒÞª˙ƒ…√◊ø≈¡£∂æ–‘‘™ÀÿªÚ≤ªø…ΩµΩ‚µƒ≤–¡ÙŒÔ“≤¥Ê‘⁄∞≤»´“˛ªº°£∂¯FNAs «”…∫ÀÀ·◊È≥…µƒ,“Ú¥Àæþ”–ÃÏ»ªµƒ…˙ŒÔœý»ð–‘∫Õ…˙ŒÔø…ΩµΩ‚–‘µƒ”≈ ∆,¥ÀÕ‚,FNAsƒÐ𪓑ÕÍ»´Õ≥“ªµƒ≥þ¥ÁΩ¯––◊È◊∞≤¢«“æþ”–¡º∫√µƒ“©ŒÔ◊∞‘ÿƒÐ¡¶,À¸√«µƒ»˝Œ¨º∏∫Œ–Œ◊¥°¢±Ì√ʪؗß∫Õ∂Øèπ¶ƒÐ∂º «æ´◊ºø…øÿµƒ°£¡ÌÕ‚”–“»Àπ§øπÔ÷Æ≥∆µƒDNA ≈‰Ãµƒ∑¢’π∏≥”Ë¡ÀDNA ∂±π¶ƒÐ,÷∏þ¡À“©ŒÔµðÀÕµƒ∞–œÚ–‘,’‚–©”≈ ∆ πµ√FNAs≥…Œ™“©ŒÔµðÀÕµƒ∂¿Ãÿ‘ÿð£Douglasµ»[44]2012ƒÍ ◊œ»ππΩ®¡ÀƒÐπª∞–œÚµðÀÕ“‘º∞øÿ÷∆“©ŒÔ Õ∑≈µƒDNAƒ…√◊Ω·ππ,Õ®π˝‘⁄DNAƒ…√◊∫–±Ì√Ê–Þ Œ“ª∏ˆΩªª• Ω“À¯”,∂¯Ω‚ø™’‚∏ˆ“À¯”µƒ“‘ø≥◊”∑÷◊”’˝ «Œª”⁄ƒø±Íœ∏∞˚µƒ±Ì√Ê, µœ÷¡À∑÷◊”∞–œÚ“‘º∞“©ŒÔ Õ∑≈µƒæ´»∑µ˜øÿ(Õº4(c))°£Liµ»[45]‘⁄¥Àª˘¥°…œ∫œ◊˜ø™∑¢¡À“ª÷÷ƒ…√◊ª˙∆˜»À”√”⁄÷Œ¡∆÷◊¡ˆ(Õº4(d))°£’‚—˘“ª∏ˆø™¥¥–‘µƒπ§◊˜’˝ «ª˘”⁄FNAsÕÍ≥…µƒ, ◊œ»‘⁄90nm×60nmµƒ’€÷Ω±Ì√Ê–Þ Œ4∏ˆƒ˝—™√∏,»ª∫ÛΩ´∂˛Œ¨’€÷ΩæÌ≥…÷–ø’πÐ(÷±æ∂‘º19nm),’‚—˘ƒ˝—™√∏æÕ∏Ω◊≈‘⁄÷–ø’πЃ⁄≤ø,‘Ÿ”√“ªœµ¡–µƒ“ΩÙπú˛”Ω´∆‰πÃ∂®,Õ¨ ±,“ΩÙπú˛”◊˜Œ™∫À» Àÿ ≈‰Ãªπæþ”–∑÷◊”¥´∏–∆˜µƒπ¶ƒÐ,À¸ƒÐÃÿ“Ï–‘Ω·∫œ÷◊¡ˆ—™πбͺ«ŒÔ∫À» Àÿµ∞∞◊°£æ≤¬ˆ◊¢…‰÷Æ∫Û,‘⁄æ≠π˝’˝≥£◊È÷Ø—™πÐ ±,DNAƒ…√◊Ω·π𱣪§ƒ˝—™√∏ π∆‰≤ª З™–°∞Â∫Õ—™Ω¨œÀŒ¨‘≠µƒ”∞œÏ,‘⁄µΩ¥Ô÷◊¡ˆ—™πÐ◊È÷Ø ±,∫À» µ∞∞◊”ΓΩÙπú˛”Ω·∫œ π∆‰Ω‚¿Î¥”∂¯µº÷¬πÐ◊¥Ω·ππ∆∆¡—,FNAsµƒππœÛ∏ƒ±‰ πƒ˝—™√∏±©¬∂‘⁄—™“∫÷–,”’µº÷◊¡ˆ≤øŒªµƒ—™“∫ƒ˝πÃ,“÷÷∆÷◊¡ˆ…˙≥§≤¢µº÷¬÷◊¡ˆªµÀ¿°£”…”⁄À˘”–µƒ µÃ¡ˆ◊È÷Ø—™πÐ∂º «ª˘±æ“ª÷¬µƒ,À˘“‘‘≠‘Ú…œ’‚÷÷∑Ω∑®ø…“‘”––ßµÿ∂‘øπ∂ý÷÷¿ý–Õµƒ÷◊¡ˆ,æþ”–Ωœ∏þµƒ∆’ –‘°£æ°πÐ’˚∏ˆπ˝≥Ö˓™∂ý∏ˆ◊Ⱥ˛µƒ≤Œ”Î,µ´∆‰ µ’˚∏ˆÃÂœµ «“◊”⁄∫œ≥…µƒ,“ÚŒ™’‚–©◊Ⱥ˛(ΩÙπú˛,“©ŒÔ∫Õ≥…œÒ±Í÷æŒÔ)ø…“‘∫лð“◊µÿºØ≥…µΩæ´–ƒ…˺∆µƒFNAs…œ,≤¢«“æ˘“ª∂»º∞≤˙¬ ∫Ð∏þ°£Ω´∂ý∏ˆπ¶ƒÐª˘Õ≈◊È∫œ‘⁄“ª∆”––ßµÿ÷∏þ¡Àƒ…√◊ª˙∆˜»Àµƒ»Àπ§÷«ƒÐ,À¸ø…“‘∂¡»°ª∑æ≥–≈œ¢≤¢æˆ∂®∫Œ ±°¢∫Œµÿ“‘º∞ «∑Ò≤…»°––∂Ø°£‘⁄÷◊¡ˆ—–æø÷–,ƒ˝—™√∏”…”⁄∆‰∑«Ãÿ“Ï–‘µƒƒ˝Ω·◊˜”√∫Õ∂Ñ𵃗™“∫—≠ª∑∞ÎÀ•∆⁄∂¯œÞ÷∆¡À‘⁄÷◊¡ˆ÷Œ¡∆…œµƒ”¶”√,µ´ «Õ®π˝…˺∆Ãÿ∂®Ω·ππ∫Õπ¶ƒÐµƒFNAsƒÐπªÕÍ√¿µƒπʱВ‚–©œÞ÷∆°£Ω´ƒ˝—™√∏’‚÷÷≤ªŒ»∂®∑÷◊”æ´»∑◊È◊∞‘⁄±£ª§ø«øںЃ⁄,Õ¨ ±Ω´∂ý∏ˆ∑÷◊”¥´∏–∆˜“‘ƒ…√◊º∂∑÷±Ê¬ ºØ≥…µΩøںеƒ µ±Œª÷√,ΩË÷˙Ãÿ“Ï–‘µƒ÷◊¡ˆ∞–œÚ≈‰ÃÂ,ª˘”⁄»˝Œ¨FNAsµƒƒ…√◊ª˙∆˜»Àø…“‘‘⁄÷◊¡ˆ≤øŒªÃÿ“Ï–‘怺Ø≤¢“‘◊‘÷˜∑Ω Ω Õ∑≈–Ø¥¯µƒƒ˝—™√∏,¥”∂¯◊Ó¥Û≥Ã∂»µÿ∑¢ª”“©–ß,≤¢◊Ó¥Û≥Ã∂»µÿΩµµÕ∆‰∂æ–‘°£’‚∏ˆÃÂœµ“≤Õ¨—˘”¶”√”⁄”…”⁄¥Ê‘⁄Œ»∂®–‘Œ ªÚ∂æ∏±◊˜”√∂¯±ª≈≈≥˝‘⁄¡Ÿ¥≤÷ÆÕ‚µƒ∆‰À˚±∏—°“©ŒÔ°£¥ÀÕ‚,∏√ÃÂœµ‘⁄÷Œ¡∆æþ”–ªØ—߃Փ©–‘µƒ÷◊¡ˆ∑Ω√Êæþ”–∏¸∏þµƒº€÷µ,“ÚŒ™À¸µƒπ¶ƒÐª˙÷∆∫Õ∞©œ∏∞˚µƒ∑÷◊”±≥æ∞ŒÞπÿ,≤¢«“’Î∂‘“ª÷÷¡∆∑®≤˙…˙µƒƒÕ“©–‘“≤≤ªÃ´ø…ƒÐ”Ρ̓ª÷÷¡∆∑®÷ÿµ˛°£Xuµ»[46]“≤ª˘”⁄’‚∏ˆÀº¬∑Ω´µ∞∞◊∫ÕFNAs◊˜Œ™ƒ£øÈπ≤Õ¨ππΩ®¡ÀÀƒ√ÊÃÂøÚºÐΩ·ππ,“‘µ∞∞◊÷ ◊˜Œ™∂•µ„,ƒÐπª‘⁄‘ÿ“©µƒÕ¨ ± µœ÷Œªµ„Ãÿ“Ï–‘∞–œÚ ∂±(Õº4(e))°£
°°°°Õº4 »˝Œ¨¡¢ÃÂøÚºÐ∫ÀÀ·
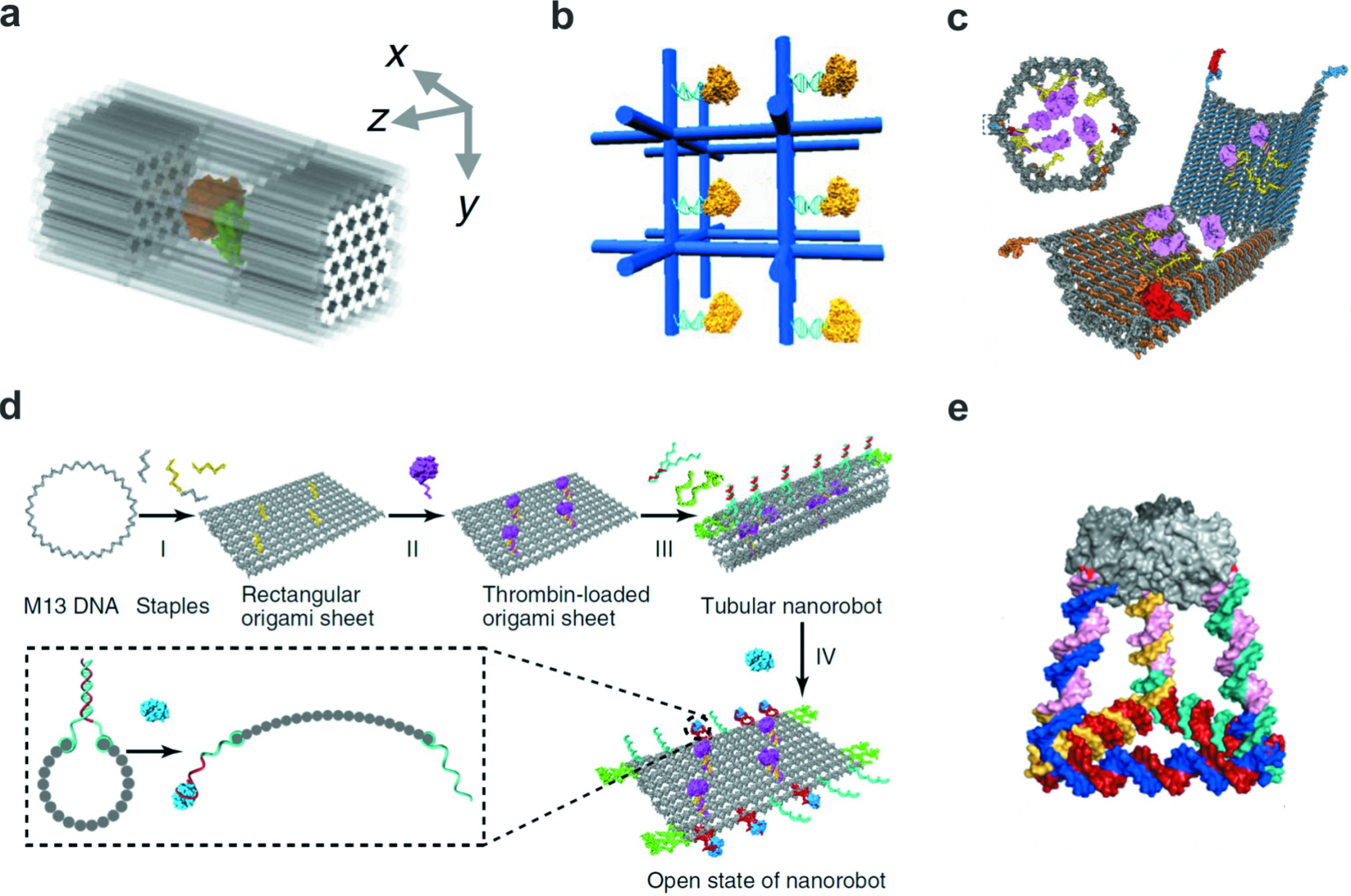
°°°°Fig.4 3D framework nucleic acids
°°°°(a)‘⁄DNAƒ…√◊¡˝◊”÷–æ´»∑◊È◊∞“ª∂‘µ∞∞◊√∏[42];(b)»˝Œ¨DNAæßÃÂ÷–æ´»∑◊È◊∞µ∞∞◊[43];(c) ≈‰ÃÂ◊˜Œ™∑÷◊”ø™πÿµƒDNAƒ…√◊ª˙∆˜»À”√”⁄‘ÿ‘Àµ∞∞◊[44];(d)øÚºÐ∫ÀÀ·ππΩ®µƒƒ…√◊ª˙∆˜»À”√”⁄÷◊¡ˆ÷Œ¡∆[45];(e)µ∞∞◊∫ÕDNAπ≤Õ¨ππΩ®µƒÀƒ√ÊÃ…√◊¡˝◊””√”⁄“©ŒÔ∞–œÚµðÀÕ[46]
°°°°(a) A pair of protein enzymes precisely organized in a DNA cage[42];(c) Scheme of aptamer-gated DNA nanorobot loaded with a protein payload.Two DNA-aptamer locks are used to control opening and closing of the device[44];(d) A FNA-based nanorobot for cancer therapy[45];(e) Tetrahedral nanocages constructed by proteins and DNA are used for targeted drug delivery[46]
°°°°4 °¢◊ÐΩ·”Î’πÕ˚
°°°°±æŒƒ∏≈¿®¡ÀDNAøÚºÐ∫ÀÀ·µƒ∏≈ƒÓ,≤¢ºÚ“™◊ÐΩ·¡ÀøÚºÐ∫ÀÀ·‘⁄µ∞∞◊µ˜øÿ∑Ω√ʵƒ◊Ó–¬Ω¯’π°£◊‘»ªΩÁ÷–µƒ–Ì∂ý–≈œ¢¥¶¿Ì∫Õπ¶ƒÐµ˜Ω⁄∂º «”…∫ÀÀ·∫Õµ∞∞◊÷ π≤Õ¨ µœ÷µƒ,»Àπ§…˺∆µƒDNA-µ∞∞◊÷ ƒ…√◊Ω·ππµƒø™∑¢‘⁄–Ì∂ý¡Ï”Ú∂º”–◊≈÷ÿ“™“‚“Â,∞¸¿®∫œ≥……˙ŒÔ—ß°¢ƒ…√◊ºº ı∫Õ…˙ŒÔ“Ω—ßµ»°£Ω¸ƒÍ¿¥,DNAƒ…√◊ºº ıµƒøÏÀŸ∑¢’π“˝∆¡Àµ∞∞◊÷ µƒæ´»∑µ˜øÿµƒ—–æø»»≥±,øÚºÐ∫ÀÀ·µƒø…±ý≥Ö‘∫Õ…˙ŒÔœý»ð–‘∂‘”⁄ππΩ®∫Õƒ£ƒ‚µ∞∞◊÷ ÃÂœµ∑Ω√Ê”–◊≈π„¿´µƒ«∞æ∞°£¿˝»Á,¿˚”√øÚºÐ∫ÀÀ·µ˜øÿµ∞∞◊º‰æý¥”∂¯ µœ÷µ∞∞◊π¶ƒÐ“‘º∞”––Ú◊È◊∞µƒµ˜øÿ,“‘øÚºÐ∫ÀÀ·◊˜Œ™÷ߺÐππΩ®√∏º∂¡™ÃÂœµ≤¢ÃΩÀ˜∆‰∑¥”¶ªÓ–‘µƒ”∞œÏ“ÚÀÿ,ππΩ®ª˘”⁄øÚºÐ∫ÀÀ·µƒƒ…√◊ª˙∆˜»À”√”⁄“©ŒÔ∞–œÚ∫Õ Õ∑≈µ»°£
°°°°…˺∆∫ÕππΩ®∏þ∂»∏¥‘”µƒDNAƒ…√◊Ω·ππµƒ∏þÀŸ∑¢’πŒ™µ˜øÿ∏˜÷÷…˙ŒÔ∑÷◊”µƒ◊È◊∞÷π©¡À“ª∏ˆæÞ¥Ûµƒª˙ª·°£»ª∂¯,DNA”Î’‚–©…˙ŒÔ∑÷◊”÷ƺ‰µƒ’˚∫œ»‘”–¥˝Ω¯“ª≤ΩÃΩÀ˜,—∞’“∏¸Œ™∆’ µƒ‘⁄≤ª”∞œÏµ∞∞◊π¶ƒÐ”Úµƒ«Èøˆœ¬∂‘DNAΩ¯––µ∞∞◊–Þ Œµƒ∑Ω∑® «ø∆—ߺ“ÿΩ¥˝Ω‚戵ƒŒ °£æ°πп˚”√∏˜÷÷DNA÷ߺÐππΩ®µ∞∞◊÷ Õ¯¬Á“—æ≠≤ø∑÷ µœ÷¡ÀÃÏ»ªÃÂœµµƒπ¶ƒÐ,µ´ªπ”–÷Ó»Á∑ΩœÚ–‘µ»∂ý÷÷œýπÿ≤Œ ˝”–¥˝Ω¯“ª≤ΩΩ¯––¿Ì¬€Ω®ƒ£∫ÕœµÕ≥ µ—È°£ ¬ µ…œ,DNAƒ…√◊ºº ı“—æ≠÷π©¡À“ª∏ˆ∏þ∂»ø…±ý≥õƒ∆Ωî,—–æø’þ√«–Ë“™Àºøº»Á∫Œª˘”⁄∫ÀÀ·øÚºÐΩ¯––∏¸…ӻεÿÃΩÀ˜”Î∑¢œ÷°£
°°°°≤ŒøºŒƒœ◊
°°°°[1] Serganov A,Nudler E.Cell,2013,152:17°´24.
°°°°[2] Forster A C,Symons R H.Cell,1987,49:211°´220.
°°°°[3] Sen D,Gilbert W.Nature,1988,334:364°´366.
°°°°[4] ’≈Àÿ∏Ò,ÀÔ∫Ïœº,Ã∆—«¡÷.ªØ—ßÕ®±®,2016,79(5):387°´394.
°°°°[5] Zeraati M,Langley D B,Schofield P,et al.Nat.Chem.,2018,10:631°´637.
°°°°[6] Seeman N C.J.Theor.Biol.,1982,99:237°´247.
°°°°[7] Kallenbach N R,Ma R I,Seeman N C.Nature,1983,305:829°´831.
°°°°[8] Kato T,Goodman R P,Erben C M,et al.Nano Ltt.,2009,9:2747°´2750.
°°°°[9] Liu X,Zhang F,Jing X,et al.Nature,2018,559:593°´598.
°°°°[10] Rothemund P W K.Nature,2006,440:297°´302.
°°°°[11] Cutler J I,Auyeung E,Mirkin C A.J.Am.Chem.Soc.,2012,134:1376°´1391.
°°°°[12] ’≈œ£Œ∞,µ•÷æ,Õıœ‘œÈ.ªØ—ßÕ®±®,2014,77(9):848°´852.
°°°°[13] Yang Y R,Liu Y,Yan H.Bioconjug.Chem.,2015,26:1381°´1395.
°°°°[14] Deng M,Li M,Li F,et al.ACS Mater.Lett.,2019,1:671°´676.
°°°°[15] Ge Z,Gu H,Li Q,et al.J.Am.Chem.Soc.,2018,140:17808°´17819.
°°°°[16] Lu N,Pei H,Ge Z,et al.J.Am.Chem.Soc.,2012,134:13148°´13151.
°°°°[17] Ye D,Zuo X,Fan C.Ann.Rev.Anal.Chem.,2018,11:171°´195.
°°°°[18] Yang F,Zuo X,Fan C,et al.Natl.Sci.Rev.,2018,5:740°´755.
°°°°[19] –ªÐ‘,π»≈Ê¡ÿ,¿Ó”Ó,µ».∑¯…‰—–æø”Î∑¯…‰π§“’—ß±®,2019,37:050101(10).
°°°°[20] Zhang H,Chao J,Pan D,et al.Nat.Commun.,2017,8:14738.
°°°°[21] Chao J,Wang J,Wang F,et al.Nat.Mater.,2019,18:273°´279.
°°°°[22] Qi X,Zhang F,Su Z,et al.Nat.Commun.,2018,9:4579.
°°°°[23] Zhang F,Jiang S,Wu S,et al.Nat.Nanotechnol.,2015,10:779°´784.
°°°°[24] Liu W,Halverson J,Tian Y,et al.Nat.Chem.,2016,8:867°´873.
°°°°[25] Li J,Pei H,Zhu B,et al.ACS Nano,2011,5:8783°´8789.
°°°°[26] Walsh A S,Yin H,Erben C M,et al.ACS Nano,2011,5:5427°´5432.
°°°°[27] Jiang D,Ge Z,Im H J,et al.Nat.Biomed.Eng.,2018,2:865°´877.
°°°°[28] He L,Lu D,Liang H,et al.J.Am.Chem.Soc.,2018,140:258°´263.
°°°°[29] Zhu G,Zheng J,Song E,et al.PNAS,2013,110:7998°´8003.
°°°°[30] Hao Y Y,Liu L,Zhang L H,et al.Nucl.Sci.Tech.,2018,29:83°´89.
°°°°[31] Chen Q,Liu H,Lee W,et al.Lab Chip,2013,13:3351°´3354.
°°°°[32] Freeman R,Stephanopoulos N,Alvarez Z,et al.Nat.Commun.,2017,8:15982.
°°°°[33] Erkelenz M,Kuo C H,Niemeyer C M.J.Am.Chem.Soc.,2011,133:16111°´16118.
°°°°[34] McMillan J R,Mirkin C A.J.Am.Chem.Soc.,2018,140:6776°´6779.
°°°°[35] Derr N D,Goodman B S,Jungmann R,et al.Science,2012,338:662°´665.
°°°°[36] Wilner O I,Weizmann Y,Gill R,et al.Nat.Nanotechnol.,2009,4:249°´254.
°°°°[37] Fu J,Liu M,Liu Y,et al.J.Am.Chem.Soc.,2012,134:5516°´5519.
°°°°[38] Fu J,Yang Y R,Johnson-Buck A,et al.Nat.Nanotechnol.,2014,9:531°´536.
°°°°[39] Nielsen T B,Thomsen R P,Mortensen M R,et al.Angew.Chem.Int.Ed.,2019,58:9068°´9072.
°°°°[40] Xin L,Zhou C,Yang Z,et al.Small,2013,9:3088°´3091.
°°°°[41] Liu M,Fu J,Hejesen C,et al.Nat.Commun.,2013,4:2127.
°°°°[42] Zhao Z,Fu J,Dhakal S,et al.Nat.Commun.,2016,7:10619.
°°°°[43] Rusling D A,Chandrasekaran A R,Ohayon Y P,et al.Angew.Chem.Int.Ed.,2014,53:3979°´3982.
°°°°[44] Douglas S M,Bachelet I,Church G M.Science,2012,335:831°´834.
°°°°[45] Li S,Jiang Q,Liu S,et al.Nat.Biotechnol.,2018,36:258°´264.
°°°°[46] Xu Y,Jiang S,Simmons C R,et al.ACS Nano,2019,13:3545°´3554.