生物工程论文

摘 要: 自2006年Takahashi和Yamanaka报道生成诱导多能干细胞(induced pluripotent stem cells, iPSCs)以来,多能干细胞领域进入了前所未有的发展状态,在疾病建模、药物发现以及细胞疗法等各方面都发挥重要作用,促进了细胞生物学和再生医学等学科的发展。目前,iPSCs技术已成为研究病理机制的重要工具,利用iPSCs技术筛选的新药物正在研发中,使用iPSCs衍生细胞的临床试验数量也在逐渐增长。iPSCs与基因编辑技术以及3D类器官相结合的最新研究进展促进了iPSCs在疾病研究中的进一步应用。本文介绍了近年来重编程方法的革新,分析了整合病毒载体系统、整合非病毒载体系统、非整合病毒载体系统以及非整合非病毒载体系统四种重编程方法的利弊;同时综述了iPSCs在疾病建模以及临床治疗等方面的最新研究进展,为促进iPSCs各领域的深入研究提供参考。
关键词: iPSCs; 重编程; 疾病建模; 细胞疗法;
Abstract: Since Takahashi and Yamanaka reported the generation of induced pluripotent stem cells(iPSCs) in 2006, the field of pluripotent stem cells has entered an unprecedented state of development. It plays an important role in disease modeling, drug discovery and cell therapy, and promotes the development of cell biology and regenerative medicine. At present, iPSC technology has become an important tool for studying of pathological mechanisms. New drugs screened by iPSC technology are being developed, and the number of clinical trials using i PSC-derived cells is gradually increasing. The latest research progress of iPSCs, combined with gene editing technology and 3D organoid methodology, promotes the further applications of iPSCs in disease research. In this review, we introduce the innovation of reprogramming methods in recent years, analyze the advantages and disadvantages of four reprogramming methods: integrated virus vector system,integrated non-viral vector system, non-integrated virus vector system and non-integrated non virus vector system. At the same time, we summarize the latest research progress on iPSCs in disease modeling and clinical treatment strategies, so as to provide a reference for further in-depth research in various fields of iPSCs.
Keyword: iPSCs; reprogramming; disease modeling; cell therapy;
最初人们认为,动物的成熟体细胞的基因组被永久锁定在体细胞状态,无法恢复到具有完全的多能性状态[1]。然而,1958年Gurdon等[2]将非洲爪蟾(Xenopus laevis)蝌蚪的体细胞核注射到同一物种的去核卵母细胞中,由此产生了一只功能完善的蝌蚪。39年后,Wilmut等[3]利用核移植技术培育了第一个哺乳动物克隆羊——“多莉”。这些克隆研究表明,分化细胞仍然保留着对机体发育至关重要的“遗传”记忆。1981年,Evans等[4]发现胚胎干细胞(embryonic stem cells,ESCs)可以从小鼠囊胚的内细胞团中获得,其后Thomson等[5]从人囊胚内细胞团成功获得了人胚胎干细胞(human embryonic stem cells,h ESCs)。ESCs是多能的,它们具有无限制的自我更新能力,同时拥有向所有细胞类型分化的潜能。1987年,Weintraub等[6]发现单个基因Myo D的表达可以将小鼠成纤维细胞转化为骨骼肌细胞。这一发现证明每种细胞都有自身的主调节基因,这些主调节基因起到维持细胞特性的作用。2006年,Takahashi和Yamanaka[7]从24个不同的基因入手,通过逆转录病毒载体向体细胞引入转录因子,结果发现引入4种转录因子Oct4、Sox2、Klf4和c-Myc (OSKM)可以将小鼠成纤维细胞转化为多能状态,从而建立了诱导多能干细胞(induced pluripotent stem cells,i PSCs)技术。

2007年,日本京都大学Takahashi研究小组和美国威斯康星大学Yu研究小组分别报道了从人成纤维细胞中产生人类诱导多能干细胞(hi PSCs)的案例[8,9]。自此,迅速发展起来的hi PSCs技术为疾病建模、药物发现和再生医学领域开启了一个令人兴奋的新时代。鉴于hi PSCs在疾病建模方面相对于传统细胞筛选的优势,将hi PSCs模型用于药效和潜在毒性的药物筛选变得越来越流行。hi PSCs的优点包括它们起源于人类、易获得、可扩展和能够产生几乎所有所需的细胞类型,避免与h ESCs相关的伦理问题,以及具有可以利用患者特定的i PSCs开发个性化药物,进行个性化治疗的潜力。随着3D培养技术以及基因编辑技术,特别是CRISPR/Cas9技术的发展,hi PSCs在人类疾病建模和基于干细胞的临床治疗方面展示出巨大的前景。本文主要介绍了重编程方法的革新以及i PSCs在人类遗传疾病建模和细胞治疗方面的最新发展,总结了目前i PSCs技术面临的挑战并探讨了解决这些问题的方法。
1、 重编程方法
1.1 、OSKM转录因子的作用
Oct4是一种与多能干细胞(PSCs)的多能性维持有关的同源域转录因子。Sox2在控制Oct4的表达中起着至关重要的作用[10],与Nanog和Oct4一起构成了多能性的关键转录网络。c-Myc是一种与多种癌症病因相关的原癌基因,负责招募染色质修饰蛋白,诱发广泛的转录激活。由于c-Myc有一定的致瘤性,后续有研究用无转化活性的L-Myc替代以解决此问题[11]。Klf4作为肿瘤蛋白或肿瘤抑制因子,是白血病抑制因子的下游靶点,能激活Sox2的表达[12]。重组转录因子与体细胞多能性相关序列结合的能力主要受DNA甲基化、组蛋白修饰和ATP依赖性染色质重塑产生的染色质结构变化的调节[13]。据Stadtfeld和Hochedlinger[14]的报道,细胞的多能性被诱导时会激发两个转录波。在第一个转录波中,c-Myc通过甲基化的H3K4me2和H3K4me3与体细胞基因组相结合,标志着染色质的重塑。随后是体细胞相关基因表达的沉默,包括Thy1、Snai1、Snai2、Zeb1和Zeb2等间充质基因[13,15]。第二个转录波更局限于染色质重塑的细胞。OSKM到达早期多能相关基因(pluripotency-associated genes,PAG)的增强子和启动子,并触发其转录和表达。在此转录波中,体细胞被迫改变其形态,加强增殖,并经历间充质样向上皮样的转化(mesenchymal-to-epithelial transition,MET)。这导致了Cdh1、Epcam和Ocln等上皮基因的上调和上皮特征基本状态的建立[16],形成了更大的ES样细胞团。但MET是一个随机和低效的过程,因为多能诱导基因上存在甲基化组蛋白,而甲基化组蛋白负责封闭染色质构象[13]。
Klf4在这两个阶段都扮演着重要的角色。首先,在第一阶段Klf4结合并激活包括E-cadherin在内的上皮基因,抑制分化基因[17]。其次,在第二阶段加速内源性Oct4和Sox2的表达,从而建立维持多能状态的自我调节环。Klf4通过控制细胞的发育、增殖、分化和凋亡等过程,在多能干细胞中发挥重要的作用。Klf4与Oct4和Sox2之间超强的相互作用激活一组如Nanog、Esrrb、Klf2、Sall4和ZFP42等转录因子,以及如Smad1和Stat3等信号通路调节因子[18]。
1.2、 重编程载体
在Yamanaka的开创性研究之后,陆续有几个研究室使用不同的方法来生成i PSCs,以满足有效治疗应用的安全性和质量标准。这些重编程策略大致分为两组:(1)通过基因整合或非整合转移系统;(2)利用病毒或非病毒方法。两组重编程策略将现有的重编程方法主要分为四种:整合病毒载体转移系统、整合非病毒载体转移系统、非整合病毒载体转移系统以及非整合非病毒载体转移系统,通过4种重编程方法将各类细胞重编程成i PSCs的过程如图1所示。每种方法都有其优缺点,目前并未找到一个无限制或无潜在不良后果的单一的重编程转移系统。
1.2.1、 整合病毒载体转移系统的重编程
i PSCs最初是通过逆转录病毒载体引入重编程因子产生的,逆转录病毒载体已被广泛用作体外和体内研究的基因转移载体[19]。在重编程过程的后期,由于表观遗传修饰[20],转入基因的表达被沉默[21],逆转录病毒载体只提供外源DNA序列的瞬时表达。若未能完全激活与多能性相关的内源基因的表达,通过逆转录病毒载体生成的i PSCs的质量将受到部分损害[22]。此外,一些报道指出,病毒转基因的重新激活及其在产生i PSCs中的残留活性会改变细胞的发育过程,并可能导致嵌合动物肿瘤的形成[23,24]。
慢病毒载体(lentivirus,LV)因其广泛的亲和性而被认为比逆转录病毒载体更加有效[24],因此LV被用来重编程许多体细胞类型。LV基因传递方法如今仍是最有效的重编程策略之一,重编程效率为0.1%~1%[14,25],但科学家们仍在努力改善这一途径的安全性[26]。在设计有效的重编程LV方面取得的进展之一是研制了一种多顺反子LV,它在一个表达系统中携带了由源自口蹄疫病毒的2A“自裂”肽序列连接的一个启动子驱动的四个重编程因子[23,27]。该系统减少了病毒在转基因细胞中整合的拷贝数,降低了转基因沉默的风险,简化了转化过程,并建立了一致的重编程因子的化学计量法[28]。此外,为了消除低效率沉默和转基因再激活的影响,通过引入可切除载体(Cre/lox P系统)[29]和可诱导系统(四环素/强力霉素诱导系统)[26,30]重组多顺反子病毒载体,整合后的转基因可以通过瞬时表达Cre从宿主细胞基因组中移除。但这种策略的转移效率很低[31],并且可能导致i PSCs突变,因为Cre/lox P系统在重新编程后可能会留下lox P的痕迹[32]。
图1 产生i PSCs的各种细胞来源和重编程方法
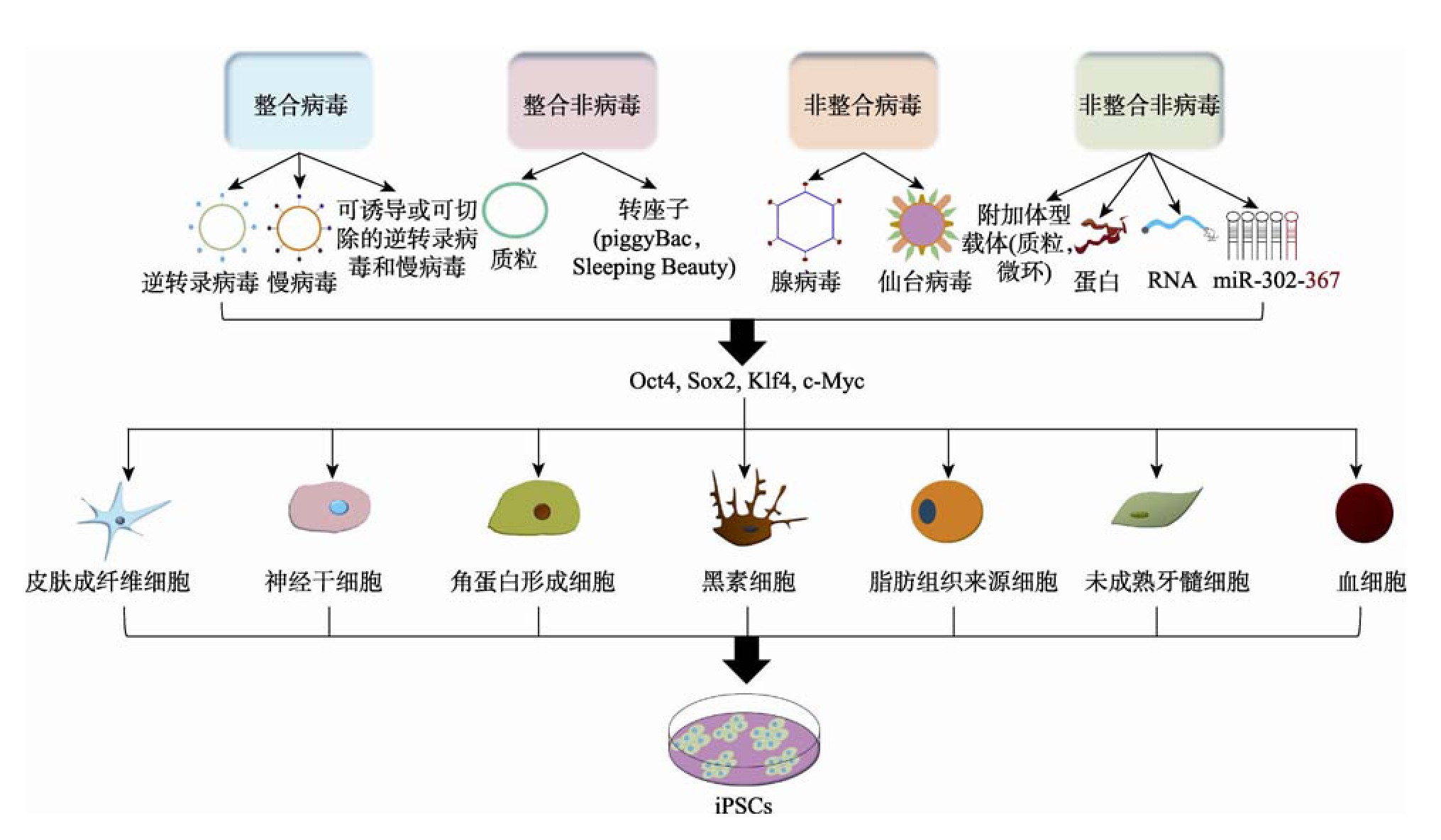
Fig.1 Various cell sources and reprogramming methods for the generation of i PSCs
1.2.2 、整合非病毒载体转移系统的重编程
由于目前整合病毒转移系统的局限性,科学家们一直在积极研究其他重编程方法,如非病毒转移系统,这些方法对于治疗应用更安全。第一个成功的非病毒性i PSCs是由成熟的胚胎成纤维细胞经两个质粒载体转染产生的:第一个质粒编码c-Myc,而第二个质粒多顺反子载体编码OSKM重编程因子[23]。这些发现表明OSKM的瞬时过表达足以诱导体细胞的多能性,但是整合的风险和重编程的效率低下又成了主要问题[33]。科学家们又设计了一个整合的依赖性基因转移载体,将转录因子整合到重组结构的lox P位点中以解决整合风险的问题[28,29]。然而,短的载体片段可以存在于切除后的基因组细胞中,这可能会影响细胞功能[33]。人们又发现利用如piggy Bac(PB)转座子等可移动的基因元件来传递外源多能性基因非常有效,且PB转座系统具有转座效率高、删除精确等优点[34]。此方法通过短暂的转位表达,可以从重新编程的细胞中完全去除该元件的残余[35]。不幸的是,人类基因组中有内源性的类PB转座子元件,在转基因切除后会引起非特异性的基因组改变[36]。为克服PB转座子的局限性,人们又引入了Sleeping Beauty(SB)系统,使其整合频率低于PB转座子,并且人类基因组中没有类SB元件[37]。然而这又导致了新的问题:此方法的重编程效率很低,使用可切除的元件还可能会导致重新整合的风险[33]。
1.2.3、 非整合病毒载体转移系统的重编程
Stadtfeld等[38]利用腺病毒等非整合病毒载体成功地建立了人和小鼠i PSCs,从这些研究中获得的i PSCs显示在宿主基因组中没有外源DNA的插入。然而,目前的非整合病毒载体传递方法的重编程效率仅限于0.001%,因此有人认为OSKM的瞬时表达不足以产生完全的表观遗传重塑[14,36]。尽管如此,腺病毒方法在转化医学中的应用仍有很大的前景[24]。另一种方法是使用负单链RNA仙台病毒(Sendaivirus,Se-V),因为它在许多类型的细胞和组织中导入外源基因非常有效,但也遇到了低重编程效率的障碍[39]。尽管如此,人们仍在努力开发一种改良的Se-V[40],因为Se-V在囊性纤维化基因治疗[41]和艾滋病疫苗[42]方面具有巨大的潜力,希望可用于人i PSCs的细胞替代治疗[43]。
1.2.4 、非整合非病毒载体转移系统的重编程
为了产生不含载体整合到染色体的i PSCs,可以使用细胞质RNA、附加体型载体(一种自我复制和选择的载体)[44]或多顺反子微环DNA非病毒载体系统[45],将多潜能标记基因直接和瞬时地传递到体细胞中。这些方法相对容易使用,但重编程效率比LV低5~10倍[24]。因此,使用细胞质RNA和微环DNA载体需要广泛的优化以备将来的应用[24]。
小鼠和人成纤维细胞已成功地通过直接转移纯化的重编程蛋白[46]或从ESCs[47]、转基因HEK293细胞中分离的总蛋白[48]提取物进行重编程。然而这种方法存在一定的弊端,因为大量合成这样的蛋白质具有很大的挑战性,且转化效率特别低,细胞重编程过程需要8周。有实验室指出通过化学重编程产生i PSCs也许是可行的,但这一过程可能导致突变,因为细胞基因组易受DNA和组蛋白修饰[14,33]。引入合成RNA或编码重编程因子的信使RNA(m RNA)也许是建立无整合多能干细胞的有力平台,尽管这些方法可能需要多轮转染,但在生成具有更安全的i PSCs方面相对有优势[49]。
1.3、 提高重编程效率的方法
为了改进重编程过程,人们采用了诸如micro RNA(mi RNA)等新方法来提高重编程效率。例如,mi R-291-3p、mi R-294和mi R-295被用来代替c-Myc以产生均匀的hi PSCs集落[50]。此外,化学化合物如丙戊酸、丁酸钠和组蛋白脱乙酰酶抑制剂等已被证明能促进i PSCs的生成[51,52,53]。培养环境的改变,如缺氧培养也能提高重编程效率[54]。抑制p53途径[55]或抑制Nu RD(Mbd3/核小体重塑和去乙酰化抑制因子)复合物的组成成分Mbd3,都可以进一步促进i PSCs的生成[56]。卵母细胞中特异表达的其他因子,如Glis1和H1foo,也能提高重编程效率[57,58]。
除了可以提高重编程效率,有些小分子在重编程过程中还能发挥部分OSKM转录因子的作用,甚至能完全代替OSKM转录因子将体细胞重编程为i PSCs。例如,使用组蛋白脱乙酰酶抑制剂和转化生长因子β抑制剂的培养基可以在没有c-Myc或Klf4的情况下提高i PSCs的生成,并且可以替代Oct4来维持细胞的多能性[59]。此外,用Aza(DNA甲基化抑制剂)和TSA(组蛋白脱乙酰酶抑制剂)诱导多能基因Oct4、Nanog、Sox2的表达,可以将小鼠骨髓前体细胞(BPCs)重编程为i PSCs[60]。另一项研究报告表明,尽管重编程效率较低,但转入mi RNA-302/367家族可以在没有转入OSKM转录因子的情况下成功地将小鼠和人类体细胞重编程为i PSCs[61]。Hou等[62]使用7种小分子化合物成功将小鼠体细胞重编程成i PSCs,且重编程效率高达0.2%。虽然单用小分子对体细胞进行重编程在临床上有很大的应用价值,但由于重编程不完全,诱导后的多能干细胞容易恢复体细胞特征,这将限制小分子重编程在临床研究中的发展。
2 、基于i PSCs的疾病建模
2.1 、基因编辑技术
基于i PSCs的疾病模型的建立和使用通常包括以下步骤:收集来源患者的体细胞,通过重编程技术将其重编程成病人特异性i PSCs;利用CRISPR/Cas9等基因编辑技术建立等基因对照(通常通过编辑已知疾病相关基因获得修复后的健康i PSCs);将细胞分化为各种与疾病相关的特定细胞,并通过比较患病和健康的特定细胞或类器官来定义疾病特异性表型;在分子水平上研究这些表型可以识别新的病理机制,为药物发现和个性化治疗提供新的机会。i PSCs衍生的特定细胞和类器官在疾病建模、药物发现和细胞治疗中的应用如图2所示。
快速发展的基因组编辑技术现在能够在特定位置将基因改变引入i PSCs,包括纠正患者来源的i PSCs中的致病基因突变以及将特定突变引入非疾病野生型i PSCs。这些方法提供以引入的突变作为唯一的变量来产生基因匹配的、等基因的i PS细胞系,确保了对真实病理的可靠鉴定,同时避免了由于可能的细胞系间变异导致的遗传背景差异或偶发现象的混淆。等基因i PSCs控制在模拟散发性或多基因疾病时尤其重要,因为这些疾病的表型差异很小[63]。
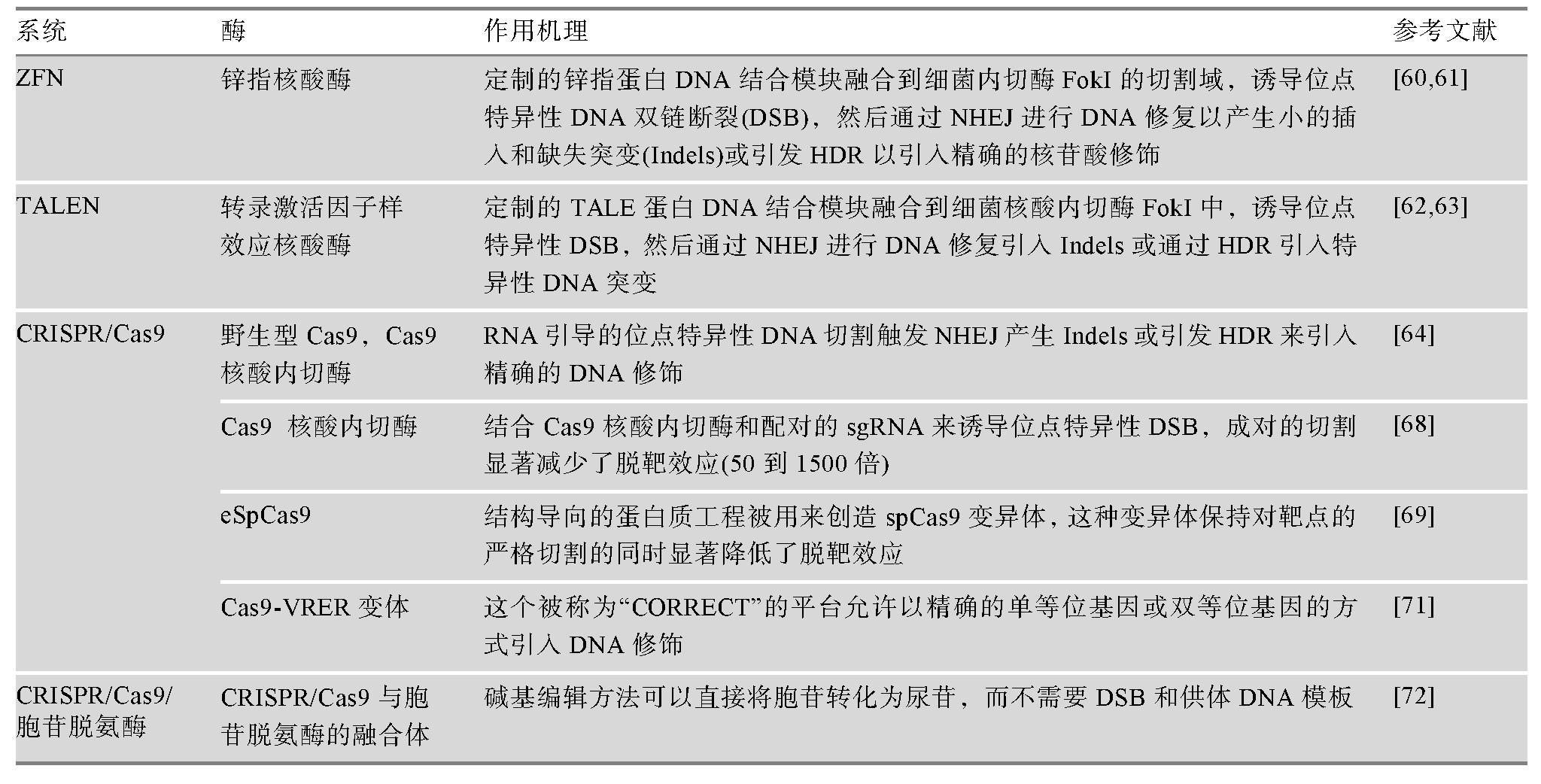
可编程位点特异性核酸酶的开发,包括锌指核酸酶(zinc-finger nuclease,ZFN)[64,65]、转录激活物样效应器核酸酶(transcription activator-like effector nucleases,TALENs)[66,67]和CRISPR/Cas9系统[68],通过诱导基因修饰位点的DNA双链断裂,显着提高了人ESCs和i PSCs的基因编辑效率。其中CRISPR/Cas9技术由于设计简单、使用方便等优点,在人类胚胎干细胞和i PSCs的基因编辑中得到了广泛的应用。这种基因编辑技术允许研究人员将致病突变引入非患者i PSCs或消除患者i PSCs中的致病突变,从而创建基于i PSCs的疾病建模等基因控制平台。尽管使用CRISPR/Cas9技术可能会产生偏离目标的效果,如CRISPR/Cas9在肿瘤细胞系[69]中发现了相对较高水平的非靶基因修饰,但来自多个实验室的全基因组测序(whole genome sequencing,WGS)研究表明,非靶基因修饰在正常人类细胞(包括人i PSCs和ESCs)中很少见[70,71]。基因编辑工具也在不断改进和完善,这可能有助于解决脱靶效应的问题。最初CRISPR/Cas9通过诱导DNA双链断裂来编辑一个基因组位点;后来由成对向导RNA或特异性增强的工程化Cas9核酸酶变体(engineered Cas9 nuclease variants with enhanced specificity,e Sp Cas9)指导的Cas9核酸内切酶变体(D10A突变体)越来越多地用于基因组编辑[72],因为这两种方法都能在保持对靶点的严格切割的同时显着降低了脱靶效应[73]。此外,催化死亡的Cas9 (dead Cas9,d Cas9)通过与转录激活剂或抑制因子融合,用于调节内源基因的转录或通过与荧光蛋白融合来成像基因组位点[74]。对CRISPR/Cas9系统的改造也能以精确的单等位或双等位基因的方式高效地引入DNA序列的变化[75]。最近在碱基编辑方面的一个新进展是利用CRISPR/Cas9与胞苷脱氨酶融合,使胞苷直接转化为尿苷而不需要双链DNA断裂[76]。这种新方法提高了基因编辑效率,并将进一步促进人类i PSCs的基因编辑发展。用于人类i PSCs的基因编辑技术以及作用机理如表1所示。
图2 来源于i PSCs的特定细胞和类器官在疾病建模、药物发现和细胞治疗中的应用
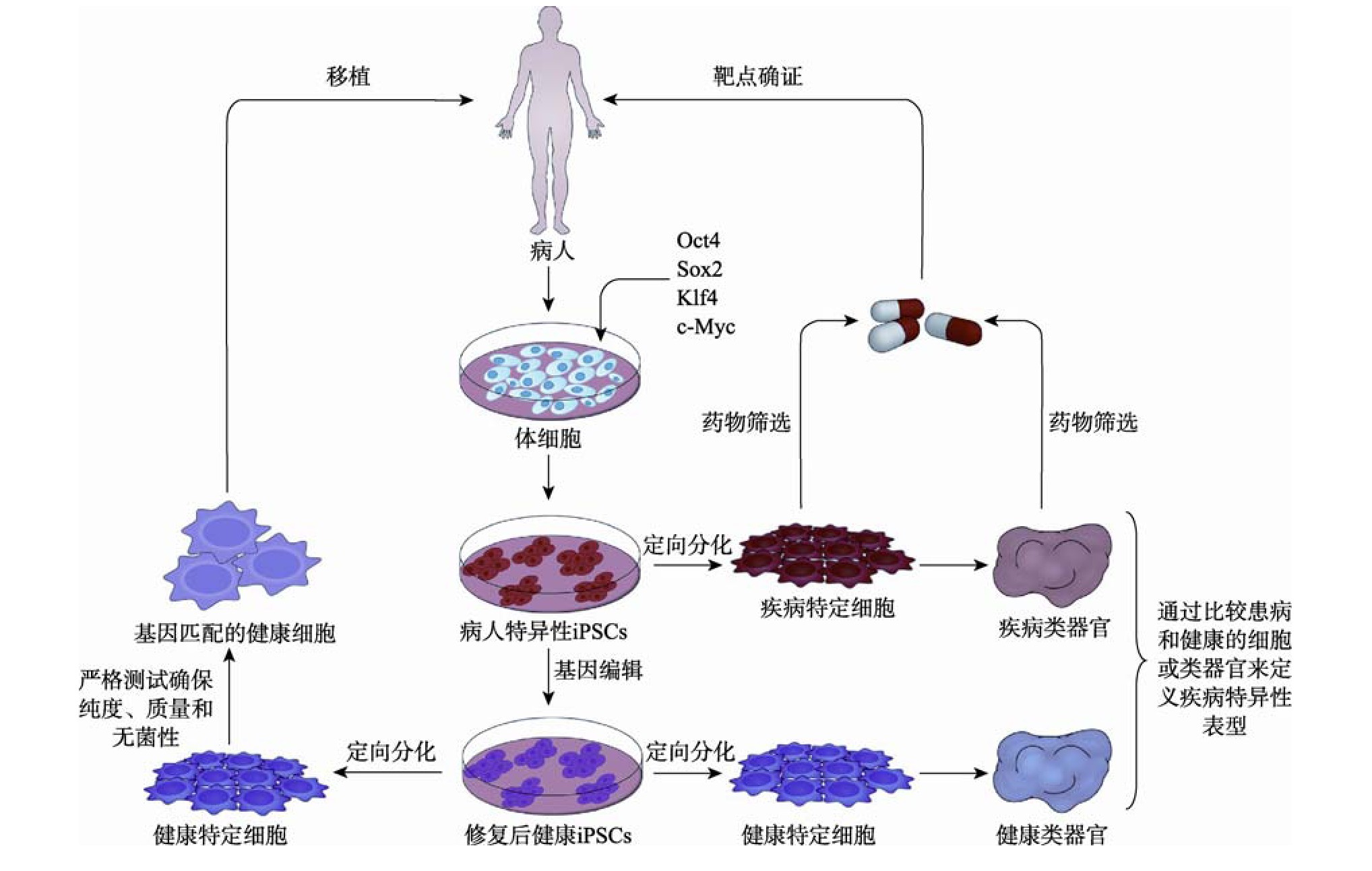
Fig.2 Application of specific cells and organoids derived from i PSCs in disease modeling,drug discovery and cell therapy
2.2 、传统二维细胞建模
识别人类疾病的病理机制对于发现新的治疗策略具有关键作用。虽然利用原代患者来源细胞建立疾病模型有助于研究人类疾病的病因和制定治疗策略,但一个关键的限制因素是无法获得可扩展的患者原代细胞,尤其是脑细胞和心脏细胞。人类i PSCs由于自我更新的内在特性和几乎可以分化为体内任何细胞类型的潜能而成为一个有吸引力的替代品,因为人类疾病(特别是那些有明确遗传原因的疾病)原则上可以使用容易获得的细胞类型(如皮肤成纤维细胞和血细胞)的i PSCs建模。患者特异性i PSCs可以提供大量与疾病相关的细胞和各种以前无法获得的细胞类型,如神经元和心肌细胞。此外,由于i PSCs可以从相关患者自身获得,因此它们可以使个性化疾病建模成为精准医学的核心部分。
基于i PSCs的疾病模型被广泛用于研究由单基因突变引起的疾病(单基因疾病)[77],这种方法非常适合于此类疾病,因为i PSCs可以很容易地从这些疾病患者身上获得并分化成如神经元等与疾病相关的细胞。例如,从患者i PSCs中分化出来的神经元被用来模拟脊髓性肌萎缩症(spinal muscular atrophy,SMA),这是一种由SMN1基因突变引起的早发性疾病,SMN1基因突变导致运动神经元变性和随后的肌肉萎缩[77]。Ⅰ型SMA患者通常在出生后6个月出现症状,疾病进展迅速,在两岁时死亡[78]。在最初的基于i PSCs的疾病模型研究中[77],i PSCs来源于Ⅰ型SMA患者的成纤维细胞,并分化为与疾病相关的细胞类型——运动神经元。与未受影响的对照组相比,患者i PSCs分化的运动神经元存活率降低。此外,来源于SMA患者的i PSCs能够对丙戊酸和妥布霉素(已知两种化合物能增加SMN蛋白的表达量)产生反应,且这两种化合物可以提高SMA患者i PSCs的SMN蛋白水平[77]。这项研究提供了一个原则性的证据,病人来源的i PSCs可以用来模拟早发性遗传疾病,并可作为潜在的药物筛选平台。
表1 用于人类i PSCs的基因编辑技术
对发病较晚的疾病进行建模更具挑战性,因为从人类i PSCs分化出来的细胞通常表现出不成熟的表型。例如,理想情况下来源于i PSCs的心肌细胞在用于再生医学或药物发现时应该是成熟的,其成熟度将类似于成年心肌中心肌细胞的成熟度,从而使衍生细胞显示出相似的收缩性、电生理性能和对药物刺激的反应。然而,在现实中,i PSCs来源的心肌细胞是未成熟的,其形态特征等方面更符合胚胎状态下的心肌细胞[79]。未成熟心肌细胞较少显示有组织的肌节结构和钙处理机制,这些特征反映在成熟相关肌节基因(如MYL2、MYH7、TCAP和MYOM2)和离子转运相关基因(如KCNJ2和RYR2)的低表达上[80]。研究人员已在诱导心肌细胞成熟方面做了很多努力:添加甲状腺激素[81],长期培养[82],用三维心肌组织中的机械调节结合电刺激[83]等方法都对i PSCs衍生心肌细胞的成熟有作用。除了要解决i PSCs衍生细胞不成熟表型的问题外,建模晚发性疾病更大的困难在于如何诱导细胞衰老。对人i PSCs衍生的细胞进行诱导衰老的一种方法是用细胞应激源处理细胞,包括如吡咯菌素和MG-132等的靶向线粒体功能或蛋白质降解的化合物[84];另一种诱导细胞衰老的方法是异位表达如早衰蛋白等诱导早衰的基因产物[85]。然而,细胞应激源或早衰蛋白的表达是否能通过一种类似于正常衰老的机制诱导细胞衰老仍有待确定。
i PSCs还为研究散发性疾病提供了一种新的方法(其病因尚未在患者的家族史或基因突变中确定),这一点十分重要,因为许多疾病的大多数患者都属于散发性疾病。例如,在老年痴呆症中,95%的病人属于散发性疾病。研究人员通过对散发性阿尔茨海默病患者i PSCs衍生神经细胞的分析发现,一些散发性病例表现出与特定基因突变的家族性阿尔茨海默病相同的表型[86],这表明使用i PSCs也许可以对散发性疾病进行重新鉴定。然而,使用i PSCs对散发性疾病进行建模通常比单基因疾病更困难,因为这类疾病的表型变化是由多个小效应遗传风险变异体和环境因素共同诱导的。虽然来源于此类疾病患者的i PSCs可能包含与疾病相关的风险变异体,但使用i PSCs来模拟此类疾病由于遗传和表观遗传背景的逐行变异而变得复杂,而且散发性疾病i PSCs的衍生细胞的表型预计比单基因疾病i PSCs的衍生细胞更为精细。因此,基于i PSCs的散发性疾病模型的一个关键问题是如何产生仅在相关风险变异上存在差异的成对等基因细胞系[63]。利用CRISPR/Cas9技术产生基因编辑的等基因i PS细胞系的功能可以创造一个良好的控制系统,确保与疾病相关的基因风险变异体成为唯一的变量[87]。这种方法被用于产生帕金森病不同相关风险变异体的等基因i PS细胞系,结合等位基因特异性分析能够对该遗传风险变异体进行强有力的分辨和基因定位[87]。这一实验策略可用于研究与其他疾病相关的遗传危险因素。
此外,基于i PSCs的疾病模型也成为研究肿瘤疾病的发病机制和筛选新的治疗药物的有用工具。在不改变细胞基因组序列的情况下,来源于肿瘤疾病患者的任何细胞都能重编程成i PSCs,再分化为与对应肿瘤疾病相关的细胞类型,以此研究i PSCs向特异性肿瘤细胞转化的过程。例如,青少年骨髓单核细胞白血病(juvenile myelomonocytic leukemia,JMML)是一种侵袭性骨髓增生性肿瘤,其发生的原因是突变导致细胞因子受体信号转导异常。将两名JMML患者的骨髓细胞和外周血细胞重编程成i PSCs,此i PSCs具有与患者体细胞相同的PTPN11基因的p.E76K错义突变,该突变体编码SHP-2 (一种非受体酪氨酸磷酸酶)[88]。与正常i PSCs体外分化产生的髓系细胞相比,JMML-i PSCs衍生的髓系细胞增殖能力增强、粒细胞–巨噬细胞集落刺激因子(GM-CSF)被激活且STAT5/ERK磷酸化增强,这些特征与JMML患者的原代髓系细胞相似,且MEK激酶的药理抑制作用能降低JMML-i PSCs衍生的髓系细胞GM-CSF的活性[88]。这项研究表明i PSCs衍生细胞在人类原发性恶性肿瘤体外建模中的效用,为该疾病潜在的靶向治疗提供理论依据。基于i PSCs的疾病建模在遗传性肿瘤疾病的研究中也展示出巨大的前景。Li-Fraumeni综合征(Li-Fraumeni syndrome,LFS)是一种常见的由p53基因突变引起的恶性肿瘤综合征[89]。来源于LFS患者的i PSCs衍生的成骨细胞显示出分化缺陷性和致瘤性,且H19基因的表达受阻,这些特征与骨肉瘤(osteosarcoma,OS)细胞一致。恢复LFS-i PSCs中H19的表达促进了成骨细胞的分化并抑制了其致瘤性[90]。该研究结果证明了用i PSCs研究遗传性人类肿瘤疾病的可行性。
2.3 、类器官:培养皿里的复杂组织
i PSCs衍生产品的临床应用在很大程度上依赖于定向分化、细胞状态转换和组织工程的最新技术。早期利用i PSCs定向分化的研究经常利用单基因疾病中的细胞水平表型,但转化为组织水平和器官水平的疾病需要开发更复杂的3D多细胞系统。随着i PSCs衍生类器官的发展,i PSCs在疾病建模方面取得了重要进展[91]。类器官是由干细胞分化而来的三维多细胞聚集体,能够自我组织,可以表现成熟组织的结构特征和细胞间的相互作用[92]。用于指导类器官与i PSCs分化的可溶性生物物理学方法已逐渐完善,以产生越来越复杂的“人造组织”[91]。患者来源的i PSCs和基因纠正的i PSCs平行分化成类器官,可以将类器官水平的疾病表型归因于特定的分子病变。一旦建立了清晰的类器官水平研究平台,病变的类器官就可以用于药物筛选和验证研究。
早期的ESCs和i PSCs研究利用神经分化方法来模拟神经系统疾病[93]。从人i PSCs衍生的3D大脑类器官的创建就是建立在这些基本的神经分化方法的基础上,但随后要在悬浮生物反应器中生长长达70天,另外这些3D大脑类器官还给研究人员带来形态建成的启发线索[94]。这些器官包含脑皮层组织区域以及特定前脑和后脑区域的功能性神经元,甚至包含未成熟视网膜和脉络丛的分化结构[94]。Quadrato和Sloan等[95,96]对3D大脑类器官培养方法的改进还能诱导3D类器官形成大脑中的海马体和小脑等特定区域结构,以及诱导形成皮层折叠结构。神经类器官因为显着的复杂性而被用来模拟各种单基因和多基因神经系统疾病,这些疾病的研究为神经疾病的病理生物学提供了深厚的基础。例如,CDK5RAP2编码一种中心体蛋白,该蛋白在有丝分裂过程中定位于纺锤体极,而CDK5RAP2基因的复合杂合子突变将引起小头畸形。CDK5RAP2基因突变患者i PSCs衍生的神经类器官最初被用来模拟小头畸形,由此发现患者特有的神经类器官经历了神经上皮早期分化,并显示出异常的径向胶质取向和较小的分化神经组织区域[94]。而另一种编码中心体蛋白的CENPJ基因在微管组装和成核的调节中起重要作用,CENPJ的突变将引起小头综合征,该病患者i PSCs衍生的神经类器官中也观察到类似的表型[97]。近年来,来源于人i PSCs的神经类器官还被用作研究脑肿瘤的平台,研究人员使用含有神经干细胞和前体细胞的大脑类器官,通过载体转化以引入常见于脑肿瘤的突变,然后研究突变后细胞的过度生长问题以模拟脑瘤的发生和发展[98]。因此,与2D系统相比,神经类器官的研究让人们增强了对神经系统疾病病理生物学的了解。
在从i PSCs生成肝细胞治疗各种肝脏疾病方面的二维分化方案已被证明是有效的[99]。最近,在三维类器官系统中,从患有肝胆疾病的i PSCs中已经衍生出胆管细胞。目前的分化方案是首先定向分化为内皮细胞和肝母细胞,然后在三维培养中分化为胆管细胞样细胞[100,101]。这些胆管细胞类器官可以流出胆汁酸并具有功能性分泌作用,从而可以模拟Alagille综合征,这是一种由于Notch信号中断而导致胆管形成受损的疾病[101]。在与多囊性肝病患者分化的胆管细胞类器官中,合成生长抑素类似物奥曲肽降低了类器官的大小,与该药物在治疗患者多囊性肝肿大中的作用一致[101]。携带F508del突变的多囊性肝病和囊性纤维化(CF)患者的i PSCs衍生的胆管细胞类器官显示出氯化物转运受损和CFTR蛋白表达量降低,而CF校正药物VX809能稳定该类器官中CFTR蛋白的表达,与该药物在囊性纤维化患者中的作用一致[100,101]。在每一个病例中,复杂疾病表型的建模都是通过三维类器官的发展而实现的。
近年来,心脏类器官的研究取得了极大的进展。i PSCs衍生的心脏类器官具有胎儿样分化的特点,虽已被用于心肌细胞损伤后再生的模型研究,但是这一发现强化了一种观点,即来源于i PSCs的心肌细胞和其他组织经常表现出胎儿样分化,这可能会妨碍成人疾病的建模[102,103]。最近有研究者产生了具有中心空腔的复杂心肌细胞类器官,这让分化方案逐步完善[104]。此外,i PSCs已经被用于构建三维心肌组织,以评估机械力、代谢和细胞外基质对心肌细胞成熟的影响,这些研究包括使用i PSCs的组织工程的各个方面[105]。研究人员还通过3D器官芯片技术将i PSCs用于模拟心肌病,这一策略被应用在许多其他疾病系统中,为模拟血管灌注提供了额外的理论支持[106,107]。
最近的两项研究强调了人i PSCs衍生的肺类器官在疾病建模中的作用[108,109],方法是将人i PSCs定向分化为NKX2-1+的气道祖细胞,该祖细胞能够随后形成近端或远端气道细胞[108]。在低Wnt激活的三维培养中,NKX2-1+祖细胞可重复形成包含各种近端气道细胞的类器官,包括分泌细胞、杯状细胞和基底细胞[108]。用forskolin同时处理来自囊性纤维化F508del-CFTR突变纯合子患者和来自对照的健康人类的i PSCs衍生的近端气道类器官,相比之下,患者衍生的类器官显示forskolin诱导的肿胀受损[108]。研究人员将这一表型归因于CFTR功能障碍,而后i PSCs中F508del的基因纠正改善了forskolin诱导的肿胀[108],进而揭示了基因编辑在i PSCs疾病模型中确认的基因型-表型关系。人i PSCs来源的NKX2-1+肺祖细胞在3D培养中也能分化为远侧肺泡类器官,这些肺泡类器官含有功能性的2型肺泡上皮细胞,这些细胞含有板层小体并分泌表面活性蛋白。而来自缺乏表面活性蛋白B (SFTPB)患者的i PSCs,其衍生的肺泡类器官虽然含有2型肺泡上皮细胞,但这些细胞缺少板层小体或不能合成SFTPB[109]。以上研究结果表明通过基因编辑来纠正SFTPB突变也许能改变该疾病患者i PSCs衍生细胞的表型。
2.4、 人–动物嵌合体
尽管基于类器官的疾病模型在复杂性、成熟度和细胞多样性方面有所拓展,但还是局限于组织培养,限制了与体内循环、神经和免疫系统、激素以及其他代谢介质相互作用的探索[110]。此外,类器官还缺少体内存在的许多形态发生信号[92]。例如,在模拟血液疾病时,体外分化系统很少将微环境因子纳入造血分化和造血干细胞(HSCs)的研究中[111,112]。近年来,细胞和类器官异种移植与人i PSCs的研究进展已经克服了这些局限性。鉴于最近许多细胞治疗的临床研究案例呈爆炸性增长之势,而基于i PSCs的异种移植嵌合体疾病模型已经能够更加忠实地模拟人类疾病并成为推动这些研究进展的可靠工具。
人类神经组织的异种移植已经用于研究人类疾病几十年了,人胚胎干细胞的分离培养、i PSCs技术的发展、人胚胎干细胞的神经分化倾向以及移植模型生物,都为细胞治疗提供了一种新型的治疗技术[113]。当人类i PSCs分化的神经元异种移植到发育中的小鼠脑中时可以整合、形成功能性突触并整合到神经回路中[114]。i PSCs分化的内源性神经干细胞(NSCs)在大鼠脑内植入后可分化为神经元和星形胶质细胞,并可促进缺血性脑卒模型的恢复[115]。从人类或非人类灵长类动物i PSCs中分化出来的多巴胺能神经元可以植入大鼠的大脑,改善帕金森病模型的功能;非人类灵长类i PSCs的多巴胺能神经元也可以自体移植,并在体内长期植入[116]。这种令人信服的功能恢复的临床前证据为人类帕金森病的临床试验提供了强有力的证明;而采用细胞疗法修复海马体内的神经连接,目前正在作为研究精神分裂症的治疗方法[117]。最近,宿主血管系统灌注和神经活动的电生理学实验描述了将人i PSCs衍生的神经类器官成功植入到成年小鼠脑中,这为神经系统疾病的全方位建模提供了一个新平台[118]。总之,这些实验证明了嵌合模型在用i PSCs模拟人类神经系统疾病中的价值以及基于i PSCs的细胞治疗在这些疾病中的可行性。
研究人员已经将人i PSCs衍生的类器官移植到小鼠体内,以整合血管系统来研究与正常宿主生理的相互作用[119]。来源于人i PSCs的肝内胚层细胞可以与内皮干细胞和间充质干细胞联合培养以形成原始的肝芽,这些肝芽异种移植后与小鼠血管系统形成功能联系,然后将人白蛋白释放到小鼠血清中并产生人类特有的药物代谢产物[119]。研究结果还表明移植这些肝芽可以提高小鼠肝衰竭模型的存活率[119]。从家族性高胆固醇血症患者的i PSCs中分化出的肝细胞已经被异种移植来模拟这种疾病[120]。
将肿瘤异种移植到免疫缺陷小鼠模型中是近年来肿瘤研究的一项关键技术。在人类i PSCs恶性血液肿瘤模型中,已经开发出形成造血嵌合体的案例[121]。急性髓系白血病(acute myeloid leukaemia,AML)细胞在MLL(也称为KMT2A)基因座上发生易位,可以通过阻断异常的MLL驱动的DNA甲基化并有效地将其重新编程为多能性细胞,并能再分化为非造血系细胞[121]。然而,在定向分化为造血祖细胞后,这些细胞在培养和移植过程中表现出异常的自我更新能力,因此该系统可用于模拟人AML的克隆结构,并可用于区分这些i PSCs衍生亚克隆的药物敏感的差异性[121]。基于人类i PSCs的嵌合体也被用来模拟实体肿瘤。人胶质母细胞瘤细胞重编程成i PSCs可消除抑瘤基因启动子的高甲基化功能,当i PSCs定向分化为神经祖细胞时,这些细胞保留了恶性潜能,在异种移植到小鼠纹状体时形成侵袭性肿瘤[122]。从患有Li–Fraumeni综合征的患者中获得的i PSCs衍生的成骨细胞携带有种系TP53突变,这些患者易患多种实体恶性肿瘤和血液系统恶性肿瘤,在免疫缺陷小鼠皮下注射后形成骨肉瘤样肿瘤[123]。今后利用来源于i PSCs的类器官和异种移植到小鼠体内的研究可以为实体瘤的发生和发展提供新的视角。
4 、基于i PSCs的临床研究
除了疾病建模之外,i PSCs最大的潜力之一是对许多遗传和退行性疾病进行细胞和基因替代治疗[124]。在这种方法中,病人的体细胞被分离和培养,再通过病毒或非病毒介导的基因转移被重新编程成i PSCs,接着使用基因编辑技术或病毒转导方法对患者来源的i PSCs进行基因纠正,基因纠正筛选出的i PSCs分化为受影响的细胞亚型。在进行细胞鉴定以及纯度、活性和安全性的质量控制试验后,将基因匹配的健康细胞移植到患者体内进行细胞治疗。这种自体移植的方法可以防止严重的并发症,如通常发生在同种异体移植后的移植物抗宿主病。i PSCs在疾病治疗中的应用如图3所示。
然而,在常规的临床应用开始之前,基于iPSCs的治疗有几个障碍需要解决,其中一个是iPSCs的致瘤风险[125]。由于iPSCs在培养基中培养了很长时间,它们会积累核型异常、拷贝数变异和杂合性丧失[126]。因此,在临床使用之前,iPSCs衍生产品需要仔细筛选以确定是否存在潜在风险的基因改变[126],并进行严格测试以确保其纯度、质量和无菌性。另一个问题是生成可用于临床的iPSCs的成本很高且耗时较长,解决这个问题需要建立能够储存大量人类白细胞抗原(HLA)纯合超供体iPSCs的细胞库[127]。其中一个例子是2013年日本成立了京都大学iPSCs研究与应用中心(CiRA)。CiRA是一家异基因iPSCs库,收集了50份来自健康志愿者的外周血T淋巴细胞和脐血HLA纯合标本,在HLA的三个主要位点(HLA-A、HLA-B和DRB1)进行匹配[128]。2015年,Cellular Dynamic International,Inc.还成立了一家细胞治疗库,可以满足19%的美国人口需求。人们相信,以可承受的成本为数百万患者提供大量合适细胞的HLA单体型纯合iPS细胞库可能是一种有效的细胞治疗的策略。第三个障碍是iPSCs衍生细胞的异质性和不成熟性。通常,iPSCs衍生的细胞是异质的。例如,在iPSCs衍生神经元细胞的各种方案中,衍生细胞常常包含不同类型的神经元和其他细胞(如胶质细胞)[129]。在临床使用前,必须严格控制这种异质性,以确保植入细胞的一致性。因此,可以通过荧光活化细胞分选法(fluorescence-activated cell sorting,FACS)或磁激活细胞分选法(magneticactivated cell sorting,MACS)对iPSCs衍生细胞进行分类[130],仅选择临床研究需要的相关衍生细胞。从人类iPSCs分化出的各类细胞都显示出类似胚胎或新生细胞的表型,这表明iPSCs衍生的细胞是不成熟的。添加甲状腺激素[81]、长期培养[82]等方法都能促进iPSCs衍生细胞成熟,将iPSCs衍生的组织作为嵌合体植入动物体内将进一步促进iPSCs衍生的组织成熟[131],但iPSCs衍生细胞仍然与成熟细胞的状态有一定差距。异种移植和类器官技术的进一步发展将有助于克服这个障碍,进而促进iPSCs临床研究的发展。
图3 iPSCs在疾病治疗中的应用
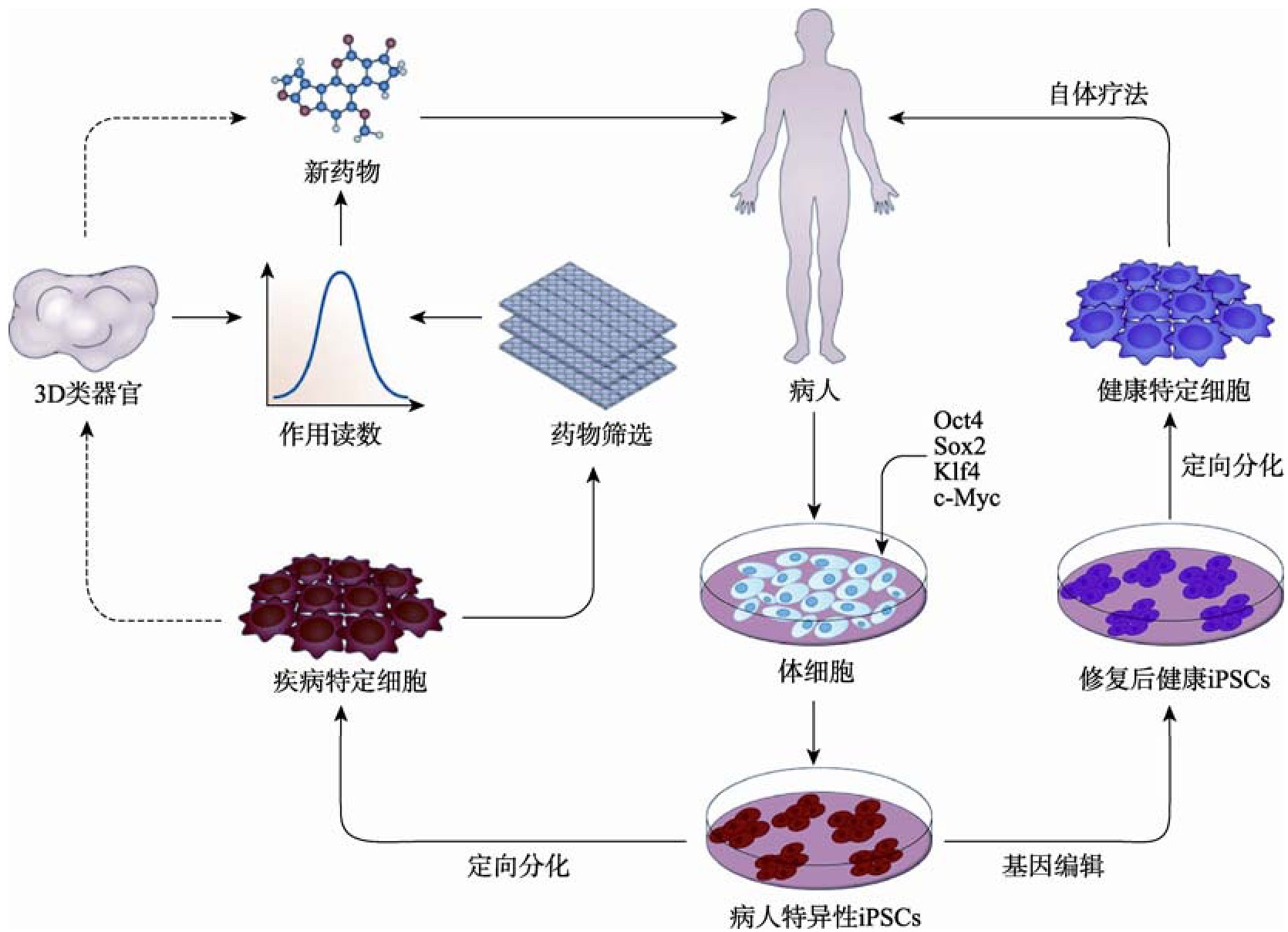
Fig.3 Application of iPSCs in disease treatment
第一次i PSCs临床试验(第一阶段)于2014年9月在日本进行。来自RIKEN发展生物学中心的高桥雅代证明了基于i PSCs的视网膜色素变性(RP)治疗的安全性,RP的主要特征是眼睛内额外的血管形成导致视力完全丧失。患者在接受自体i PSCs衍生的视网膜色素上皮细胞移植治疗后,视力有所改善[132]。与此相反的是,Kuriyan等[133]报告了使用自体脂肪干细胞治疗老年性黄斑变性(AMD)的灾难性结果,即患者视力迅速丧失,需要紧急护理。因为不需要大量的制备细胞,使用自体i PSCs产品进行个性化药物治疗似乎是单基因疾病的理想选择,但对于更常见的疾病,特别是如心肌梗死等急性常见疾病,自体i PSCs疗法可能不适用于大部分患者,因为仔细检验每个细胞系需要高昂的成本和大量的时间。也因为这些原因,日本i PSC-RPE实验的第二阶段将使用异基因产品[134]。这些事件表明了基于i PSCs的细胞治疗还有很长的一段路要走。
嵌合抗原受体(chimeric antigen receptor,CAR)T细胞治疗策略已被证明能成功地介导血液恶性肿瘤的消退。但大量制备自体T细胞可能成为负担,特别是由于免疫抑制或细胞毒性治疗、体外低淋巴细胞计数和健康T淋巴细胞的无效扩增[135]。解决方案是对患者衍生的i PSCs进行改造,从而生成和HLA无关的可定制抗原识别的抗原特异性的CART细胞。经基因组编辑的完全相同的人类i PSCs (不论其HLA单倍型如何)可以进一步分化为全功能的组织相容性肿瘤靶向T细胞[136]。迄今为止,动物模型的临床前研究表明,i PSCs衍生的工程CART细胞(i CART)细胞具有抗肿瘤活性[137]。2019年,Ci RA和Takeda合作宣布,第一个i CART细胞治疗计划将于2021年开始临床试验第一阶段。i CART治疗可以根据需要为患者量身定制现成的免疫疗法[138]。i CART杀死癌细胞的潜在益处,凸显了临床进化和治疗应用的一个重要里程碑。
与T细胞相比,自然杀伤细胞(NK细胞)是一种淋巴细胞,在天然免疫系统介导抗肿瘤和抗病毒活性的能力中发挥着关键作用而无需MHC限制。应用NK细胞免疫疗法治疗急性髓系白血病的临床试验已显示出显着的疗效,而对其他恶性肿瘤的疗效则较低[139]。但NK细胞与T细胞的健康细胞群都很难获得,通常由单核细胞和其他血细胞的异质混合物组成。i PSCs的出现提供了一种独特的解决方案:可以生产同质和明确的NK细胞群,这些NK细胞很容易通过基因改造来提高抗肿瘤活性,用于临床大规模生产“现成”细胞[140]。修饰后的NK细胞可以表达多种受体,如高亲和力DC16-Fc受体或CAR受体,也可以与其他疗法联合应用来提高其抗实体瘤的能力[141]。到目前为止,第一项研究显示i PSCs作为一个平台生产带有靶向肿瘤的CAR受体的NK细胞的可行性和有效性,在卵巢癌异种移植模型中显示出令人印象深刻的抗肿瘤活性,且该细胞在体内的存活扩展毒性较小[142]。2018年11月,人类临床试验(NCT0384110)获得美国食品和药物管理局(FDA)批准,首次使用克隆i PSCs产生的现成NK细胞(命名为FT500)与T细胞协同作用,以更好地治疗实体恶性肿瘤。向三名患者给药(每次给药含1×108细胞,并结合检查点抑制剂和单克隆抗体),初步安全性评估显示在最初28天的观察期内没有严重不良事件。2019年2月,FT500治疗方案在64名不同恶性肿瘤患者中进行了进一步的安全性测试,临床试验目前正在进行中(NCT0384110)。日本京都大学已授权武田制药(Takeda Pharmaceuticals)开发基于i PSCs的现成CART细胞平台并将其商业化,第一次临床试验的目标日期为2021年。将这些临床前研究转化为临床试验的潜力是令人期待的。
5 、结语与展望
自i PSCs技术建立以来,尽管人们在研究疾病机制和潜在治疗方面通过将人类i PSCs与其他新技术相结合而取得了很大进展,但仍有几个重要问题有待解决。除i PSCs衍生的如心肌细胞、肾细胞等各类细胞都显示出成熟度的不足外,i PSCs在重编程过程中与细胞特性、年龄、代谢相关的体细胞记忆的存在也格外引人注目。考虑到i PSCs可能保留与肿瘤细胞部分相关的体细胞记忆(癌基因表达、Warburg代谢谱)[143,144],筛选和消除这些记忆以减少移植时肿瘤形成的风险,此类研究更为急迫且意义重大,但是目前还缺乏有效的方法来避免或消除这些体细胞记忆。
综上所述,i PSCs技术的发展为定义和治疗疾病提供了一种强有力的新方法,因为i PSCs代表了一种范式的转变:允许人们直接观察和治疗相关的病人细胞,揭示了基因表达的新关系,拓宽和加深了人们对各种疾病发展的理解。以CRISPR/Cas9、3D类器官、动物嵌合体和micro RNA分子开关等为代表的新技术的出现和不断革新,将进一步推动基于i PSCs的疾病建模和治疗发展。虽然i PSCs的应用尚处于起步阶段,应用风险较大,但相信在下一个15年,i PSCs的应用会在个体化再生医学和肿瘤免疫治疗方面展现出广阔的前景。
参考文献
[1] Waddington CH.The strategy of the genes.Allen and Unwin:London,1957,Reprinted 2014.
[2] Gurdon JB,Elsdale TR,Fischberg M.Sexually mature inpiduals of Xenopus laevis from the transplantation of single somatic nuclei.Nature,1958,182(4627):64-65.
[3] Wilmut I,Schnieke AE,Mc Whir J,Kind AJ,Campbell KH.Viable offspring derived from fetal and adult mammalian cells.Nature,1997,385(6619):810-813.
[4] Evans MJ,Kaufman MH.Establishment in culture of pluripotential cells from mouse embryos.Nature,1981,292(5819):154-156.
[5] Thomson JA,Itskovitz-Eldor J,Shapiro SS,Waknitz MA,Swiergiel JJ,Marshall VS,Jones JM.Embryonic stem cell lines derived from human blastocysts.Science,1998,282(5391):1145-1147.
[6] Davis RL,Weintraub H,Lassar AB.Expression of a single transfected c DNA converts fibroblasts to myoblasts.Cell,1987,51(6):987-1000.
[7] Takahashi K,Yamanaka S.Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors.Cell,2006,126(4):663-676.
[8] Takahashi K,Tanabe K,Ohnuki M,Narita M,Ichisaka T,Tomoda K,Yamanaka S.Induction of pluripotent stem cells from adult human fibroblasts by defined factors.Cell,2007,131(5):861-872.
[9] Yu JY,Vodyanik MA,Smuga-Otto K,AntosiewiczBourget J,Frane JL,Tian SL,Nie J,Jonsdottir GA,Ruotti V,Stewart R,Slukvin II,Thomson JA.Induced pluripotent stem cell lines derived from human somatic cells.Science,2007,318(5858):1917-1920.
[10] Masui S,Nakatake Y,Toyooka Y,Shimosato D,Yagi R,Takahashi K,Okochi H,Okuda A,Matoba R,Sharov AA,Ko MS,Niwa H.Pluripotency governed by Sox2via regulation of Oct3/4 expression in mouse embryonic stem cells.Nat Cell Biol,2007,9(6):625-635.
[11] Nakagawa M,Takizawa N,Narita M,Ichisaka T,Yamanaka S.Promotion of direct reprogramming by transformation-deficient Myc.Proc Natl Acad Sci USA,2010,107(32):14152-14157.
[12] Niwa H,Ogawa K,Shimosato D,Adachi K.A parallel circuit of LIF signalling pathways maintains pluripotency of mouse ES cells.Nature,2009,460(7251):118-122.
[13] Takahashi K,Yamanaka S.A decade of transcription factor-mediated reprogramming to pluripotency.Nat Rev Mol Cell Biol,2016,17(3):183-193.
[14] Stadtfeld M,Hochedlinger K.Induced pluripotency:history,mechanisms,and application.Genes Dev,2010,24(20):2239-2263.
[15] Soufi A,Donahue G,Zaret KS.Facilitators and impediments of the pluripotency reprogramming factors’initial engagement with the genome.Cell,2012,151(5):994-1004.
[16] Papp B,Plath K.Epigenetics of reprogramming to induced pluripotency.Cell,2013,152(6):1324-1343.
[17] Plath K,Lowry WE.Progress in understanding reprogramming to the induced pluripotent state.Nat Rev Genet,2011,12(4):253-265.
[18] Nakatake Y,Fukui N,Iwamatsu Y,Masui S,Takahashi K,Yagi R,Yagi K,Miyazaki JI,Matoba R,Ko MSH,Niwa H.Klf4 cooperates with Oct3/4 and Sox2 to activate the Lefty1 core promoter in embryonic stem cells.Mol Cell Biol,2006,26(20):7772-7782.
[19] J?hner D,Stuhlmann H,Stewart CL,Harbers K,L?hler J,Simon I,Jaenisch R.De novo methylation and expression of retroviral genomes during mouse embryogenesis.Nature,1982,298(5875):623-628.
[20] Matsui T,Leung D,Miyashita H,Maksakova IA,Miyachi H,Kimura H,Tachibana M,Lorincz MC,Shinkai Y.Proviral silencing in embryonic stem cells requires the histone methyltransferase ESET.Nature,2010,464(7290):927-931.
[21] Stadtfeld M,Maherali N,Breault DT,Hochedlinger K.Defining molecular cornerstones during fibroblast to i PS cell reprogramming in mouse.Cell Stem Cell,2008,2(3):230-240.
[22] Sridharan R,Tchieu J,Mason MJ,Yachechko R,Kuoy E,Horvath S,Zhou Q,Plath K.Role of the murine reprogramming factors in the induction of pluripotency.Cell,2009,136(2):364-377.
[23] Okita K,Nakagawa M,Hyenjong H,Ichisaka T,Yamanaka S.Generation of mouse induced pluripotent stem cells without viral vectors.Science,2008,322(5903):949-953.
[24] Lee CS,Bishop ES,Zhang RY,Yu XY,Farina EM,Yan SJ,Zhao C,Zheng ZY,Shu Y,Wu XY,Lei JY,Li YS,Zhang WW,Yang C,Wu K,Wu Y,Ho S,Athiviraham A,Lee MJ,Wolf JM,Reid RR,He TC.Adenovirusmediated gene delivery:potential applications for gene and cell-based therapies in the new era of personalized medicine.Genes Dis,2017,4(2):43-63.
[25] Somers A,Jean JC,Sommer CA,Omari A,Ford CC,Mills JA,Ying L,Sommer AG,Jean JM,Smith BW,Lafyatis R,Demierre MF,Weiss DJ,French DL,Gadue P,Murphy GJ,Mostoslavsky G,Kotton DN.Generation of transgene-free lung disease-specific human induced pluripotent stem cells using a single excisable lentiviral stem cell cassette.Stem Cells,2010,28(10):1728-1740.
[26] Maherali N,Ahfeldt T,Rigamonti A,Utikal J,Cowan C,Hochedlinger K.A high-efficiency system for the generation and study of human induced pluripotent stem cells.Cell Stem Cell,2008,3(3):340-345.
[27] Abbar AA,Nordin N,Ngai SC,Abdullah S.Production of lentiviral vector with polycistronic transcripts for reprogramming of mouse fibroblast cells.Pertanika JSci Technol,2018,26(2):627-640.
[28] Kaji K,Norrby K,Paca A,Mileikovsky M,Mohseni P,Woltjen K.Virus-free induction of pluripotency and subsequent excision of reprogramming factors.Nature,2009,458(7239):771-775.
[29] Soldner F,Hockemeyer D,Beard C,Gao Q,Bell GW,Cook EG,Hargus G,Blak A,Cooper O,Mitalipova M,Isacson O,Jaenisch R.Parkinson's disease patientderived induced pluripotent stem cells free of viral reprogramming factors.Cell,2009,136(5):964-977.
[30] Wernig M,Lengner CJ,Hanna J,Lodato MA,Steine E,Foreman R,Staerk J,Markoulaki S,Jaenisch R.A druginducible transgenic system for direct reprogramming of multiple somatic cell types.Nat Biotechnol,2008,26(8):916-924.
[31] Zhang X,De Los Angeles A,Zhang J.The art of human induced pluripotent stem cells:the past,the present and the future.Open Stem Cell J,2010,2:2-7.
[32] Li J,Song W,Pan GJ,Zhou J.Advances in understanding the cell types and approaches used for generating induced pluripotent stem cells.J Hematol Oncol,2014,7:50.
[33] Yamanaka S.A fresh look at i PS cells.Cell,2009,137(1):13-17.
[34] Qian QJ,Che JQ,Ye LP,Zhong BX.The improvement and application of piggy Bac transposon system in mammals.Hereditas(Beijing),2014,36(10):965-973.钱秋杰,车家倩,叶露鹏,钟伯雄.piggy Bac转座系统的功能改进及在哺乳动物中的应用.遗传,2014,36(10):965-973.
[35] Woltjen K,Michael IP,Mohseni P,Desai R,Mileikovsky M,H?m?l?inen R,Cowling R,Wang W,Liu PT,Gertsenstein M,Kaji K,Sung HK,Nagy A.Piggy Bac transposition reprograms fibroblasts to induced pluripotent stem cells.Nature,2009,458(7239):766-770.
[36] Medvedev SP,Shevchenko AI,Zakian SM.Induced pluripotent stem cells:problems and advantages when applying them in regenerative medicine.Acta Nature,2010,2(2):18-27.
[37] Davis RP,Nemes C,Varga E,Freund C,Kosmidis G,Gkatzis K,de Jong D,Szuhai K,Dinnyés A,Mummery CL.Generation of induced pluripotent stem cells from human foetal fibroblasts using the Sleeping Beauty transposon gene delivery system.Differentiation,2013,86(1-2):30-37.
[38] Stadtfeld M,Nagaya M,Utikal J,Weir G,Hochedlinger K.Induced pluripotent stem cells generated without viral integration.Science,2008,322(5903):945-949.
[39] Macarthur CC,Fontes A,Ravinder N,Kuninger D,Kaur J,Bailey M,Taliana A,Vemuri MC,Lieu PT.Generation of human-induced pluripotent stem cells by a nonintegrating RNA Sendai virus vector in feeder-free or xeno-free conditions.Stem Cells Int,2012,2012:1-9.
[40] Kawagoe S,Higuchi T,Otaka M,Shimada Y,Kobayashi H,Ida H,Ohashi T,Okano HJ,Nakanishi M,Eto Y.Morphological features of i PS cells generated from Fabry disease skin fibroblasts using Sendai virus vector(Se Vdp).Mol Genet Metab,2013,109(4):386-389.
[41] Yonemitsu Y,Kitson C,Ferrari S,Farley R,Griesenbach U,Judd D,Steel R,Scheid P,Zhu J,Jeffery PK,Kato A,Hasan MK,Nagai Y,Masaki I,Fukumura M,Hasegawa M,Geddes DM,Alton EW.Efficient gene transfer to airway epithelium using recombinant Sendai virus.Nat Biotechnol,2000,18(9):970-973.
[42] Takeda A,Igarashi H,Kawada M,Tsukamoto T,Yamamoto H,Inoue M,Iida A,Shu T,Hasegawa M,Matano T.Evaluation of the immunogenicity of replication-competent V-knocked-out and replicationdefective F-deleted Sendai virus vector-based vaccines in macaques.Vaccine,2008,26(52):6839-6843.
[43] Nakanishi M,Otsu M.Development of Sendai virus vectors and their potential applications in gene therapy and regenerative medicine.Curr Gene Ther,2012,12(5):410-416.
[44] Yu JY,Hu KJ,Smuga-Otto K,Tian SL,Stewart R,Slukvin II,Thomson JA.Human induced pluripotent stem cells free of vector and transgene sequences.Science,2009,324(5928):797-801.
[45] Jia FJ,Wilson KD,Sun N,Gupta DM,Huang M,Li ZJ,Panetta NJ,Chen ZY,Robbins RC,Kay MA,Longaker MT,Wu JC.A nonviral minicircle vector for deriving human i PS cells.Nat Methods,2010,7(3):197-199.
[46] Zhou WB,Freed CR.Adenoviral gene delivery can reprogram human fibroblasts to induced pluripotent stem cells.Stem Cells,2009,27(11):2667-2674.
[47] Cho HJ,Lee CS,Kwon YW,Paek JS,Lee SH,Hur J,Lee EJ,Roh TY,Chu IS,Leem SH,Kim Y,Kang HJ,Park YB,Kim HS.Induction of pluripotent stem cells from adult somatic cells by protein-based reprogramming without genetic manipulation.Blood,2010,116(3):386-395.
[48] Kim D,Kim CH,Moon JI,Chung YG,Chang MY,Han BS,Ko S,Yang E,Cha KY,Lanza R,Kim KS.Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins.Cell Stem Cell,2009,4(6):472-476.
[49] Mandal PK,Rossi DJ.Reprogramming human fibroblasts to pluripotency using modified m RNA.Nat Protoc,2013,8(3):568-582.
[50] Ong SG,Lee WH,Kodo K,Wu JC.Micro RNA-mediated regulation of differentiation and transdifferentiation in stem cells.Adv Drug Deliv Rev,2015,88:3-15.
[51] Huangfu D,Osafune K,Maehr R,Guo W,Eijkelenboom A,Chen SB,Muhlestein W,Melton DA.Induction of pluripotent stem cells from primary human fibroblasts with only Oct4 and Sox2.Nat Biotechnol,2008,26(11):1269-1275.
[52] Huangfu D,Maehr R,Guo WJ,Eijkelenboom A,Snitow M,Chen AE,Melton DA.Induction of pluripotent stem cells by defined factors is greatly improved by smallmolecule compounds.Nat Biotechnol,2008,26(7):795-797.
[53] Mali P,Chou BK,Yen J,Ye ZH,Zou JZ,Dowey S,Brodsky RA,Ohm JE,Yu W,Baylin SB,Yusa K,Bradley A,Meyers DJ,Mukherjee C,Cole PA,Cheng LZ.Butyrate greatly enhances derivation of human induced pluripotent stem cells by promoting epigenetic remodeling and the expression of pluripotency-associated genes.Stem Cells,2010,28(4):713-720.
[54] Yoshida Y,Takahashi K,Okita K,Ichisaka T,Yamanaka S.Hypoxia enhances the generation of induced pluripotent stem cells.Cell Stem Cell,2009,5(3):237-241.
[55] Hong H,Takahashi K,Ichisaka T,Aoi T,Kanagawa O,Nakagawa M,Okita K,Yamanaka S.Suppression of induced pluripotent stem cell generation by the p53-p21pathway.Nature,2009,460(7259):1132-1135.
[56] Rais Y,Zviran A,Geula S,Gafni O,Chomsky E,Viukov S,Mansour AA,Caspi I,Krupalnik V,Zerbib M,Maza I,Mor N,Baran D,Weinberger L,Jaitin DA,Lara-Astiaso D,Blecher-Gonen R,Shipony Z,Mukamel Z,Hagai T,Gilad S,Amann-Zalcenstein D,Tanay A,Amit I,Novershtern N,Hanna JH.Deterministic direct reprogramming of somatic cells to pluripotency.Nature,2013,502(7469):65-70
[57] Maekawa M,Yamaguchi K,Nakamura T,Shibukawa R,Kodanaka I,Ichisaka T,Kawamura Y,Mochizuki H,Goshima N,Yamanaka S.Direct reprogramming of somatic cells is promoted by maternal transcription factor Glis1.Nature,2011,474(7350):225-229.
[58] Kunitomi A,Yuasa S,Sugiyama F,Saito Y,Seki T,Kusumoto D,Kashimura S,Takei M,Tohyama S,Hashimoto H,Egashira T,Tanimoto Y,Mizuno S,Tanaka S,Okuno H,Yamazawa K,Watanabe H,Oda M,Kaneda R,Matsuzaki Y,Nagai T,Okano H,Yagami KI,Tanaka M,Fukuda K.H1foo has a pivotal role in qualifying induced pluripotent stem cells.Stem Cell Reports,2016,6(6):825-833
[59] Maherali N,Hochedlinger K.Tgfbeta signal inhibition cooperates in the induction of i PSCs and replaces Sox2and c Myc.Curr Biol,2009,19(20):1718-1723.
[60] Rajasingh J,Thangavel J,Siddiqui MR,Gomes I,Gao XP,Kishore R,Malik AB.Improvement of cardiac function in mouse myocardial infarction after transplantation of epigenetically-modified bone marrow progenitor cells.PLo S One,2011,6(7):e22550.
[61] Anokye-Danso F,Trivedi CM,Juhr D,Gupta M,Cui Z,Tian Y,Zhang YZ,Yang WL,Gruber PJ,Epstein JA,Morrisey EE.Highly efficient mi RNA-mediated reprogramming of mouse and human somatic cells to pluripotency.Cell Stem Cell,2011,8(4):376-388.
[62] Hou PP,Li YQ,Zhang X,Liu C,Guan JY,Li HG,Zhao T,Ye JQ,Yang WF,Liu K,Ge J,Xu J,Zhang Q,Zhao Y,Deng HK.Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds.Science,2013,341(6146):651-654.
[63] Hockemeyer D,Jaenisch R.Induced pluripotent stem cells meet genome editing.Cell Stem Cell,2016,18(5):573-586.
[64] Hockemeyer D,Soldner F,Beard C,Gao Q,Mitalipova M,De Kelver RC,Katibah GE,Amora R,Boydston EA,Zeitler B,Meng XD,Miller JC,Zhang L,Rebar EJ,Gregory PD,Urnov FD,Jaenisch R.Efficient targeting of expressed and silent genes in human ESCs and i PSCs using zinc-finger nucleases.Nat Biotechnol,2009,27(9):851-857.
[65] Zou JZ,Maeder ML,Mali P,Pruett-Miller SM,Thibodeau-Beganny S,Chou BK,Chen GB,Ye ZH,Park IH,Daley GQ,Porteus MH,Joung JK,Cheng LZ.Gene targeting of a disease-related gene in human induced pluripotent stem and embryonic stem cells.Cell Stem Cell,2009,5(1):97-110.
[66] Christian M,Cermak T,Doyle EL,Schmidt C,Zhang F,Hummel A,Bogdanove AJ,Voytas DF.Targeting DNAdouble-strand breaks with TAL effector nucleases.Genetics,2010,186(2):757-761.
[67] Hockemeyer D,Wang HY,Kiani S,Lai CS,Gao Q,Cassady JP,Cost GJ,Zhang L,Santiago Y,Miller JC,Zeitler B,Cherone JM,Meng XD,Hinkley SJ,Rebar EJ,Gregory PD,Urnov FD,Jaenisch R.Genetic engineering of human pluripotent cells using TALEnucleases.Nat Biotechnol,2011,29(8):731-734.
[68] Cong L,Ran FA,Cox D,Lin SL,Barretto R,Habib N,Hsu PD,Wu XB,Jiang WY,Marraffini LA,Zhang F.Multiplex genome engineering using CRISPR/Cas systems.Science,2013,339(6121):819-823.
[69] Fu YF,Foden JA,Khayter C,Maeder ML,Reyon D,Joung JK,Sander JD.High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells.Nat Biotechnol,2013,31(9):822-826.
[70] Smith C,Gore A,Yan W,Abalde-Atristain L,Li Z,He CX,Wang Y,Brodsky RA,Zhang K,Cheng LZ,Ye ZH.Whole-genome sequencing analysis reveals high specificity of CRISPR/Cas9 and TALEN-based genome editing in human i PSCs.Cell Stem Cell,2014,15(1):12-13.
[71] Veres A,Gosis BS,Ding QR,Collins R,Ragavendran A,Brand H,Erdin S,Cowan CA,Talkowski ME,Musunuru K.Low incidence of off-target mutations in inpidual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing.Cell Stem Cell,2014,15(1):27-30.
[72] Ran FA,Hsu PD,Lin CY,Gootenberg JS,Konermann S,Trevino AE,Scott DA,Inoue A,Matoba S,Zhang Y,Zhang F.Double nicking by RNA-guided CRISPR Cas9for enhanced genome editing specificity.Cell,2013,154(6):1380-1389.
[73] Slaymaker IM,Gao LY,Zetsche B,Scott DA,Yan WX,Zhang F.Rationally engineered Cas9 nucleases with improved specificity.Science,2016,351(6268):84-88.
[74] Chen BH,Gilbert LA,Cimini BA,chnitzbauer J,Zhang W,Li GW,Park J,Blackburn EH,Weissman JS,Qi LS,Huang B.Dynamic imaging of genomic loci in living human cells by an optimized CRISPR/Cas system.Cell,2013,155(7):1479-1491.
[75] Paquet D,Kwart D,Chen A,Sproul A,Jacob S,Teo S,Olsen KM,Gregg A,Noggle S,Tessier-Lavigne M.Efficient introduction of specific homozygous and heterozygous mutations using CRISPR/Cas9.Nature,2016,533(7601):125-129.
[76] Komor AC,Kim YB,Packer MS,Zuris JA,Liu DR.Programmable editing of a target base in genomic DNAwithout double-stranded DNA cleavage.Nature,2016,533(7603):420-424.
[77] Ebert AD,Yu JY,Rose FF Jr,Mattis VB,Lorson CL,Thomson JA,Svendsen CN.Induced pluripotent stem cells from a spinal muscular atrophy patient.Nature,2009,457(7227):277-280.
[78] Munsat TL,Davies KE.International SMA consortium meeting.Neuromuscul Disord,1992,2(5-6):423-428.
[79] Yang XL,Pabon L,Murry CE.Engineering adolescence:maturation of human pluripotent stem cell-derived cardiomyocytes.Circ Res,2014,114(3):511-523.
[80] Funakoshi S,Miki K,Takaki T,Okubo C,Hatani T,Chonabayashi K,Nishikawa M,Takei I,Oishi A,Narita M,Hoshijima M,Kimura T,Yamanaka S,Yoshida Y.Enhanced engraftment,proliferation,and therapeutic potential in heart using optimized human i PSC-derived cardiomyocytes.Sci Rep,2016,6:19111.
[81] Yang XL,Rodriguez M,Pabon L,Fischer KA,Reinecke H,Regnier M,Sniadecki NJ,Ruohola-Baker H,Murry CE.Tri-iodo-l-thyronine promotes the maturation of human cardiomyocytes-derived from induced pluripotent stem cells.J Mol Cell Cardiol,2014,72:296-304.
[82] Kamakura T,Makiyama T,Sasaki K,Yoshida Y,Wuriyanghai Y,Chen JR,Hattori T,Ohno S,Kita T,Horie M,Yamanaka S,Kimura T.Ultrastructural maturation of human-induced pluripotent stem cell-derived cardiomyocytes in a long-term culture.Circ J,2013,77(5):1307-1314.
[83] Nunes SS,Miklas JW,Liu J,Aschar-Sobbi R,Xiao Y,Zhang B,Jiang J,MasséS,Gagliardi M,Hsieh A,Thavandiran N,Laflamme MA,Nanthakumar K,Gross GJ,Backx PH,Keller G,Radisic M.Biowire:a platform for maturation of human pluripotent stem cell-derived cardiomyocytes.Nat Methods,2013,10(8):781-787.
[84] Pearson BL,Simon JM,Mc Coy ES,Salazar G,Fragola G,Zylka MJ.Identification of chemicals that mimic transcriptional changes associated with autism,brain aging and neurodegeneration.Nat Commun,2016,7:11173.
[85] Miller JD,Ganat YM,Kishinevsky S,Bowman RL,Liu B,Tu EY,Mandal PK,Vera E,Shim JW,Kriks S,Taldone T,Fusaki N,Tomishima MJ,Krainc D,Milner TA,Rossi DJ,Studer L.Human i PSC-based modeling of late-onset disease via progerin-induced aging.Cell Stem Cell,2013,13(6):691-705.
[86] Kondo T,Asai M,Tsukita K,Kutoku Y,Ohsawa Y,Sunada Y,Imamura K,Egawa N,Yahata N,Okita K,Takahashi K,Asaka I,Aoi T,Watanabe A,Watanabe K,Kadoya C,Nakano R,Watanabe D,Maruyama K,Hori O,Hibino S,Choshi T,Nakahata T,Hioki H,Kaneko T,Naitoh M,Yoshikawa K,Yamawaki S,Suzuki S,Hata R,Ueno S,Seki T,Kobayashi K,Toda T,Murakami K,Irie K,Klein WL,Mori H,Asada T,Takahashi R,Iwata N,Yamanaka S,Inoue H.Modeling Alzheimer's disease with i PSCs reveals stress phenotypes associated with intracellular Aβand differential drug responsiveness.Cell Stem Cell,2013,12(4):487-496.
[87] Soldner F,Stelzer Y,Shivalila CS,Abraham BJ,Latourelle JC,Barrasa MI,Goldmann J,Myers RH,Young RA,Jaenisch R.Parkinson-associated risk variant in distal enhancer ofα-synuclein modulates target gene expression.Nature,2016,533(7601):95-99.
[88] Gandre-Babbe S,Paluru P,Aribeana C,Chou ST,Bresolin S,Lu L,Sullivan SK,Tasian SK,Weng JL,Favre H,Choi JK,French DL,Loh ML,Weiss MJ.Patient-derived induced pluripotent stem cells recapitulate hematopoietic abnormalities of juvenile myelomonocytic leukemia.Blood,2013,121(24):4925-4929.
[89] Malkin D.p53 and the Li-Fraumeni syndrome.Cancer Genet Cytogenet,1993,66(2):83-92.
[90] Lee DF,Su J,Kim HS,Chang B,Papatsenko D,Zhao RY,Yuan Y,Gingold J,Xia WY,Darr H,Mirzayans R,Hung MC,Schaniel C,Lemischka IR.Modeling familial cancer with induced pluripotent stem cells.Cell,2015,161(2):240-254.
[91] Mc Cauley HA,Wells JM.Pluripotent stem cell-derived organoids:using principles of developmental biology to grow human tissues in a dish.Development,2017,144(6):958-962.
[92] Dutta D,Heo I,Clevers H.Disease modeling in stem cell-derived 3D organoid systems.Trends Mol Med,2017,23(5):393-410.
[93] Lee G,Papapetrou EP,Kim H,Chambers SM,Tomishima MJ,Fasano CA,Ganat YM,Menon J,Shimizu F,Viale A,Tabar V,Sadelain M,Studer L.Modelling pathogenesis and treatment of familial dysautonomia using patient-specific i PSCs.Nature,2009,461(7262):402-406.
[94] Lancaster MA,Renner M,Martin CA,Wenzel D,Bicknell LS,Hurles ME,Homfray T,Penninger JM,Jackson AP,Knoblich JA.Cerebral organoids model human brain development and microcephaly.Nature,2013,501(7467):373-379.
[95] Quadrato G,Brown J,Arlotta P.The promises and challenges of human brain organoids as models of neuropsychiatric disease.Nat Med,2016,22(11):1220-1228.
[96] Sloan SA,Darmanis S,Huber N,Khan TA,Birey F,Caneda C,Reimer R,Quake SR,Barres BA,Pa?ca SP.Human astrocyte maturation captured in 3D cerebral cortical spheroids derived from pluripotent stem cells.Neuron,2017,95(4):779-790.
[97] Gabriel E,Wason A,Ramani A,Gooi LM,Keller P,Pozniakovsky A,Poser I,Noack F,Telugu NS,Calegari F,?ari?T,Hescheler J,Hyman AA,Gottardo M,Callaini G,Alkuraya FS,Gopalakrishnan J.CPAPpromotes timely cilium disassembly to maintain neural progenitor pool.EMBO J,2016,35(8):803-819.
[98] Bian S,Repic M,Guo ZM,Kavirayani A,Burkard T,Bagley JA,Krauditsch C,Knoblich JA.Genetically engineered cerebral organoids model brain tumor formation.Nat Methods,2018,15(8):631-639.
[99] Rashid ST,Corbineau S,Hannan N,Marciniak SJ,Miranda E,Alexander G,Huang-Doran I,Griffin J,Ahrlund-Richter L,Skepper J,Semple R,Weber A,Lomas DA,Vallier L.Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells.J Clin Invest,2010,120(9):3127-3136.
[100] Ogawa M,Ogawa S,Bear CE,Ahmadi S,Chin S,Li B,Grompe M,Keller G,Kamath BM,Ghanekar A.Directed differentiation of cholangiocytes from human pluripotent stem cells.Nat Biotechnol,2015,33(8):853-861.
[101] Sampaziotis F,de Brito MC,Madrigal P,Bertero A,Saeb-Parsy K,Soares FAC,Schrumpf E,Melum E,Karlsen TH,Bradley JA,Gelson WT,Davies S,Baker A,Kaser A,Alexander GJ,Hannan NRF,Vallier L.Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation.Nat Biotechnol,2015,33(8):845-852.
[102] Voges HK,Mills RJ,Elliott DA,Parton RG,Porrello ER,Hudson JE.Development of a human cardiac organoid injury model reveals innate regenerative potential.Development,2017,144(6):1118-1127.
[103] Pagliuca FW,Millman JR,Gürtler M,Segel M,Van Dervort A,Ryu JH,Peterson QP,Greiner D,Melton DA.Generation of functional human pancreaticβcells in vitro.Cell,2014,159(2):428-439
[104] Hoang P,Wang J,Conklin BR,Healy KE,Ma Z.Generation of spatial-patterned early-developing cardiac organoids using human pluripotent stem cells.Nat Protoc,2018,13(4):723-737.
[105] Abilez OJ,Tzatzalos E,Yang HX,Zhao MT,Jung G,Z?llner AM,Tiburcy M,Riegler J,Matsa E,Shukla P,Zhuge Y,Chour T,Chen VC,Burridge PW,Karakikes I,Kuhl E,Bernstein D,Couture LA,Gold JD,Zimmermann WH,Wu JC.Passive stretch induces structural and functional maturation of engineered heart muscle as predicted by computational modeling.Stem Cells,2018,36(2):265-277.
[106] Musah S,Mammoto A,Ferrante TC,Jeanty SSF,Hirano-Kobayashi M,Mammoto T,Roberts K,Chung S,Novak R,Ingram M,Fatanat-Didar T,Koshy S,Weaver JC,Church GM,Ingber DE.Mature induced-pluripotentstem-cell-derived human podocytes reconstitute kidney glomerular-capillary-wall function on a chip.Nat Biomed Eng,2017,1:0069.
[107] Brown JA,Pensabene V,Markov DA,Allwardt V,Neely MD,Shi MJ,Britt CM,Hoilett OS,Yang Q,Brewer BM,Samson PC,Mc Cawley LJ,May JM,Webb DJ,Li DY,Bowman AB,Reiserer RS,Wikswo JP.Recreating blood-brain barrier physiology and structure on chip:a novel neurovascular microfluidic bioreactor.Biomicrofluidics,2015,9(5):054124.
[108] Mc Cauley KB,Hawkins F,Serra M,Thomas DC,Jacob A,Kotton DN.Efficient derivation of functional human airway epithelium from pluripotent stem cells via temporal regulation of Wnt signaling.Cell Stem Cell,2017,20(6):844-857.
[109] Jacob A,Morley M,Hawkins F,Mc Cauley KB,Jean JC,Heins H,Na CL,Weaver TE,Vedaie M,Hurley K,Hinds A,Russo SJ,Kook S,Zacharias W,Ochs M,Traber K,Quinton LJ,Crane A,Davis BR,White FV,Wambach J,Whitsett JA,Cole FS,Morrisey EE,Guttentag SH,Beers MF,Kotton DN.Differentiation of human pluripotent stem cells into functional lung alveolar epithelial cells.Cell Stem Cell,2017,21(4):472-488.
[110] Liu C,Oikonomopoulos A,Sayed N,Wu JC.Modeling human diseases with induced pluripotent stem cells:from 2D to 3D and beyond.Development,2018,145(5):dev156166.
[111] Sturgeon CM,Ditadi A,Awong G,Kennedy M,Keller G.Wnt signaling controls the specification of definitive and primitive hematopoiesis from human pluripotent stem cells.Nat Biotechnol,2014,32(6):554-561.
[112] Wang LS,Li L,Menendez P,Cerdan C,Bhatia M.Human embryonic stem cells maintained in the absence of mouse embryonic fibroblasts or conditioned media are capable of hematopoietic development.Blood,2005,105(12):4598-4603.
[113] Barker RA,Parmar M,Studer L,Takahashi J.Human trials of stem cell-derived dopamine neurons for Parkinson’s disease:dawn of a new era.Cell Stem Cell,2017,21(5):569-573.
[114] Espuny-Camacho I,Michelsen KA,Gall D,Linaro D,Hasche A,Bonnefont J,Bali C,Orduz D,Bilheu A,Herpoel A,Lambert N,Gaspard N,Péron S,Schiffmann SN,Giugliano M,Gaillard A,Vanderhaeghen P.Pyramidal neurons derived from human pluripotent stem cells integrate efficiently into mouse brain circuits in vivo.Neuron,2013,77(3):440-456.
[115] Yuan T,Liao W,Feng NH,Lou YL,Niu X,Zhang AJ,Wang Y,Deng ZF.Human induced pluripotent stem cell-derived neural stem cells survive,migrate,differentiate,and improve neurologic function in a rat model of middle cerebral artery occlusion.Stem Cell Res Ther,2013,4(3):73.
[116] Sundberg M,Bogetofte H,Lawson T,Jansson J,Smith G,Astradsson A,Moore M,Osborn T,Cooper O,Spealman R,Hallett P,Isacson O.Improved cell therapy protocols for Parkinson’s disease based on differentiation efficiency and safety of h ESC-,hi PSC-,and non-human primate i PSC-derived dopaminergic neurons.Stem Cells,2013,31(8):1548-1562.
[117] Donegan JJ,Lodge DJ.Cell-based therapies for the treatment of schizophrenia.Brain Res,2017,1655:262-269.
[118] Mansour AA,Gon?alves JT,Bloyd CW,Li H,Fernandes S,Quang D,Johnston S,Parylak SL,Jin X,Gage FH.An in vivo model of functional and vascularized human brain organoids.Nat Biotechnol,2018,36(5):432-441.
[119] Takebe T,Sekine K,Enomura M,Koike H,Kimura M,Ogaeri T,Zhang RR,Ueno Y,Zheng YW,Koike N,Aoyama S,Adachi Y,Taniguchi H.Vascularized and functional human liver from an i PSC-derived organ bud transplant.Nature,2013,499(7459):481-484.
[120] Yang JY,Wang Y,Zhou T,Wong LY,Tian XY,Hong XY,Lai WH,Au KW,Wei R,Liu YQ,Cheng LH,Liang GC,Huang ZJ,Fan WX,Zhao P,Wang XW,Iba?ez DP,Luo ZW,Li YY,Zhong XF,Chen SH,Wang DY,Li L,Lai LX,Qin BM,Bao XC,Hutchins AP,Siu CW,Huang Y,Esteban MA,Tse HF.Generation of human liver chimeric mice with hepatocytes from familial hypercholesterolemia induced pluripotent stem cells.Stem Cell Reports,2017,8(3):605-618
[121] Chao MP,Gentles AJ,Chatterjee S,Lan F,Reinisch A,Corces MR,Xavy S,Shen J,Haag D,Chanda S,Sinha R,Morganti RM,Nishimura T,Ameen M,Wu HD,Wernig M,Wu JC,Majeti R.Human AML-i PSCs reacquire leukemic properties after differentiation and model clonal variation of disease.Cell Stem Cell,2017,20(3):329-344.
[122] Stricker SH,Feber A,Engstr?m PG,Carén H,Kurian KM,Takashima Y,Watts C,Way M,Dirks P,Bertone P,Smith A,Beck S,Pollard SM.Widespread resetting of DNA methylation in glioblastoma-initiating cells suppresses malignant cellular behavior in a lineagedependent manner.Genes Dev,2013,27(6):654-669.
[123] Lee DF,Su J,Kim HS,Chang B,Papatsenko D,Zhao RY,Yuan Y,Gingold J,Xia WY,Darr H,Mirzayans R,Hung MC,Schaniel C,Lemischka IR.Modeling familial cancer with induced pluripotent stem cells.Cell,2015,161(2):240-254.
[124] Kiskinis E,Eggan K.Progress toward the clinical application of patient-specific pluripotent stem cells.JClin Invest,2010,120(1):51-59.
[125] Lee AS,Tang C,Rao MS,Weissman IL,Wu JC.Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies.Nat Med,2013,19(8):998-1004.
[126] Lund RJ,N?rv?E,Lahesmaa R.Genetic and epigenetic stability of human pluripotent stem cells.Nat Rev Genet,2012,13(10):732-744.
[127] Hotta A,Yamanaka S.From genomics to gene therapy:induced pluripotent stem cells meet genome editing.Annu Rev Genet,2015,49:47-70.
[128] Okita K,Matsumura Y,Sato Y,Okada A,Morizane A,Okamoto S,Hong H,Nakagawa M,Tanabe K,Tezuka K,Shibata T,Kunisada T,Takahashi M,Takahashi J,Saji H,Yamanaka S.A more efficient method to generate integration-free human i PS cells.Nat Methods,2011,8(5):409-412.
[129] Kim D,Lee DR,Kim HS,Yoo J,Jung SJ,Lim BY,Jang J,Kang HC,You S,Hwang DY,Leem JW,Nam TS,Cho SR,Kim DW.Highly pure and expandable PSA-NCAM-positive neural precursors from human ESC and i PSC-derived neural rosettes.PLo S One,2012,7(7):e39715.
[130] Doi D,Samata B,Katsukawa M,Kikuchi T,Morizane A,Ono Y,Sekiguchi K,Nakagawa M,Parmar M,Takahashi J.Isolation of human induced pluripotent stem cell-derived dopaminergic progenitors by cell sorting for successful transplantation.Stem Cell Reports,2014,2(3):337-350.
[131] Finkbeiner SR,Hill DR,Altheim CH,Dedhia PH,Taylor MJ,Tsai Y,Chin AM,Mahe MM,Watson CL,Freeman JJ,Nattiv R,Thomson M,Klein OD,Shroyer NF,Helmrath MA,Teitelbaum DH,Dempsey PJ,Spence JR.Transcriptome-wide analysis reveals hallmarks of human intestine development and maturation in vitro and in vivo.Stem Cell Reports,2015,4(6):1140-1155.
[132] Mandai M,Watanabe A,Kurimoto Y,Hirami Y,Morinaga C,Daimon T,Fujihara M,Akimaru H,Sakai N,Shibata Y,Terada M,Nomiya Y,Tanishima S,Nakamura M,Kamao H,Sugita S,Onishi A,Ito T,Fujita K,Kawamata S,Go MJ,Shinohara C,Hata KI,Sawada M,Yamamoto M,Ohta S,Ohara Y,Yoshida K,Kuwahara J,Kitano Y,Amano N,Umekage M,Kitaoka F,Tanaka A,Okada C,Takasu N,Ogawa S,Yamanaka S,Takahashi M.Autologous induced stem-cell-derived retinal cells for macular degeneration.N Engl J Med,2017,376(11):1038-1046
[133] Kuriyan AE,Albini TA,Townsend JH,Rodriguez M,Pandya HK,Leonard RE,Parrott MB,Rosenfeld PJ,Flynn HW Jr,Goldberg JL.Vision loss after intravitreal injection of autologous“Stem Cells”for AMD.N Engl JMed,2017,376(11):1047-1053.
[134] Cell Stem Cell Editorial Team.10 Questions:Clinical Outlook for i PSCs.Cell Stem Cell,2016,18(2):170-173.
[135] Themeli M,Kloss CC,Ciriello G,Fedorov VD,Perna F,Gonen M,Sadelain M.Generation of tumor-targeted human T lymphocytes from induced pluripotent stem cells for cancer therapy.Nat Biotechnol,2013,31(10):928-933.
[136] Nianias A,Themeli M.Induced pluripotent stem cell(i PSC)-derived lymphocytes for adoptive cell immunotherapy:recent advances and challenges.Curr Hematol Malig Rep,2019,14(4):261-268.
[137] Minagawa A,Yoshikawa T,Yasukawa M,Hotta A,Kunitomo M,Iriguchi S,Takiguchi M,Kassai Y,Imai E,Yasui Y,Kawai Y,Zhang R,Uemura Y,Miyoshi H,Nakanishi M,Watanabe A,Hayashi A,Kawana K,Fujii T,Nakatsura T,Kaneko S.Enhancing T cell receptor stability in rejuvenated i PSC-derived T cells improves their use in cancer immunotherapy.Cell Stem Cell,2018,23(6):850-858.e4.
[138] News and events.Ci RA,Kyoto University.2019.Available at:www.cira.kyoto-u.ac.jp/e/pressrelease/news/190716-150000.html (accessed September25,2019).
[139] Morvan MG,Lanier LL.NK cells and cancer:you can teach innate cells new tricks.Nat Rev Cancer,2016,16(1):7-19.
[140] Lupo KB,Matosevic S.Natural killer cells as allogeneic effectors in adoptive cancer immunotherapy.Cancers(Basel),2019,11(6):769.
[141] Chen S,Sun H,Miao K,Deng CX.CRISPR-Cas9:from genome editing to cancer research.Int J Biol Sci,2016,12(12):1427-1436.
[142] Li Y,Hermanson DL,Moriarity BS,Kaufman DS.Human i PSC-derived natural killer cells engineered with chimeric antigen receptors enhance anti-tumor activity.Cell Stem Cell,2018,23(2):181-192.e5.
[143] Lo Sardo V,Ferguson W,Erikson GA,Topol EJ,Baldwin KK,Torkamani A.Influence of donor age on induced pluripotent stem cells.Nature Biotechnology,2017,35(1):69-74.
[144] Hanahan D,Weinberg RA.Hallmarks of cancer:the next generation.Cell,2011,144(5):646-674.