红细胞是血液基本功能的重要组成,红细胞功能的异常直接影响到气体的运输及血液粘度,从而直接影响到机体的正常生理活动和运动能力。牛磺酸( Taurine,Tau) 又名为牛胆酸或牛胆素,是中药牛黄的主要成分,化学名称为 2 - 氨基乙磺酸( 2 - Amin-oethyanesulfonic acid) .国内外的学者对 Tau 的生理作用已经进行了充分的研究[1],其在抗氧化、缓解运动性疲劳[2 -4]和调节细胞稳态[5]方面发挥重要作用。在剧烈运动或者大负荷长时间过度训练中,由于机体运动量加大,呼吸活动及物质代谢和能量代谢加速,自由基水平升高,自由基介导的脂质过氧化速率加快,脂质过氧化过程中会产生以丙二醛( Malondialdehyde,MDA) 为代表的许多含有 α-、β-不饱和醛酮类物质( 也叫 TBARS) ,这些物质在机体的衰老和疲劳过程中有重要的毒副作用[6 -7].Tau 作为一种抗氧化剂和自由基清除剂及其抗疲劳作用在细胞和动物水平已经有了充分的研究,结果表明,Tau 可降低剧烈运动导致的自由基水平升高[8],可显著延长大鼠的游泳能力[9].
以往研究人员在机体的抗氧化研究中,MDA 往往只认为是氧化应激水平的指标,而忽视 MDA 本身也是导致机体运动能力下降的原因及造成机体细胞结构功能损伤的重要因素[10].本研究通过比较过度训练大鼠模型和喂食 MDA 大鼠,比较二者红细胞在结构上的损伤性变化,采用 Tau 保护过度训练大鼠与喂食 MDA 大鼠,通过扫描电镜观察其对红细胞形态结构的影响,其对红细胞溶血反应和膜脂流动性的影响,对红细胞膜蛋白电泳特征的影响,探讨 MDA 在运动性疲劳中的作用及 Tau在抗运动性疲劳中的可能机制。
1 材料与方法
1. 1 实验动物与过度训练方案
40 只成年 SD 大鼠购自中南大学湘雅医学院实验动物中心,雌雄不限,体重 220 ~275 g.在我校实验动物房中喂养 1 周后,行力竭运动3 d( 1 次/d) ,淘汰游泳能力最强的前5 只和游泳能力最差的后 5 只大鼠,余下 30 只随机分成 6组,其中 A、B 组各 3 只,其余各组 6 只。A 组为自来水对照组,常规饲养,不加其它干预措施; B 组为 PBS对照组,常规饲养,以 PBS 代替自来水; C 组为喂食MDA 组,MDA 以 PBS 为溶剂配制; D 组为喂食 MDA +Tau 组,MDA 和 Tau 以 PBS 为溶剂配制; E 组为过度运动训练组,常规饲养; F 组为过度运动训练 + 喂食 Tau组,Tau 以 PBS 为溶剂配制。
1. 2 研究方法
1. 2. 1 游泳过度训练方案
筛选的 30 只 SD 大鼠休息饲养 3 d 后对 E、F 组进行 4 周负重游泳过度训练,每周负重量依次为: 3% 体重、5% 体重、7% 体重、9%体重,每次游泳至力竭。力竭标准为大鼠头沉入水下持续 10 s 以上,捞起后呈低头俯卧位,呼吸深急。
1. 2. 2 Tau 配制、MDA 的制备
Tau 溶液的配制按照参考文献[9]提供的方法配制。先将 Tau 溶解于 1N 的盐酸中,然后用1N 的 NaOH 中和到 pH 7. 0 后,过滤除菌,最后用 PBS 配制成浓度为 10% 的 Tau 溶液。Tau喂食剂量为每 100 g 大鼠体重 10 mg( 即每 100 g 体重大鼠喂食 1 mL) .MDA 的制备按照文献[11]提供的方法配制,用 TMP 制成 10 mM 浓度的 MDA,喂食剂量为每 100 g 大鼠 1 mL.
1. 2. 3 仪器和试剂
主要仪器有日本日电 JSM -2840 型扫描电镜,日本 Hitachi F - 4500 型荧光分光光度计,美国 Bio-Rad 5070 型通用电泳仪。主要试剂有1,1,3,3 - 四甲氧基丙烷 ( 1,1,3,3 - tetramethoxypro-pane,TMP) ,DTT( 1,4 - dithiothreitol) ,DPH( 1,6 - di-phenyl - 1,3,5 - hexatriene) ,四氧化锇( osmium tetrox-ide) 购自 Sigma 公司; 蛋白裂解试剂盒购自上海润城生物科技公司; Tris,TEMED,SDS,Gly 等电泳用常规试剂购自 Amresco 公司,其它试剂购自上海生工生物工程公司;
1. 2. 4 红细胞扫描电镜观察样品的制备
处死大鼠从腹主动脉处收集全血,全血用10 倍体积的生理盐水洗涤后,2 000 r/min 离心去上清,重复 2 次即得压积红细胞,圧积红细胞加入到含 3% 的戊二醛的甲次砷酸盐的缓冲液( pH7. 4) 中固定至少 1 h 后,植入预先包被有多聚赖氨酸的玻片,经 1% 的四氧化锇固定后,乙醇梯度脱水,CO2临界点干燥后于扫描电镜下观察。
1. 2. 5 红细胞膜的制备
红细胞膜按照依照 Dodge等的方法制备[12].红细胞加入到在冰箱中预冷的生理盐水中( 1: 9 体积比) ,离心( 2 000 r/min,5 min) 洗涤 3 次后,按照红细胞与双蒸水体积比 1: 9 的比例低渗裂解红细胞,置 4 ℃ 冰箱中 3 h 后,离心( 6 000 r/min,10 min) ,去上清,用双蒸水离心洗涤 4 ~ 5 次后,直到收集的红细胞膜无红色。最后将收集的红细胞膜悬浮在 pH7. 4 的 PBS 缓冲液中备用。
1. 2. 6 红细胞膜流动性的测定
制备的红细胞膜按照按 Lowry 法[13]测定膜蛋白浓度,用 PBS 配成红细胞膜悬液( 膜蛋白浓度在 300 ~350 mg/L) .取红细胞膜悬液 2. 0 mL 加入含 DPH 浓度为 2. 0 × 10- 6mol / L 的PBS 2. 0 mL,在 25 ℃ 水浴 30 min 后,以激发和发射波长分别为 360 nm 和430 nm 测定计算各组荧光偏振度( P) ,P = ( Ivv-G × Ivh) / ( Ivv+ G × Ivh) ,G 为校正因子,G= Ihv/ Ihh.其中 Ivv为起偏器和检偏器为垂直时的荧光强度; Ivh为起偏器为垂直和检偏器为水平时的荧光强度; Ihv为起偏器为水平和检偏器为垂直时的荧光强度; Ihh起偏器和检偏器为水平时的荧光强度度。荧光偏振度( P) 与微粘度( η) 的定量关系[14]: η = P/( 0. 46 - P) .膜流动性用膜微粘度表示,膜微粘度越大,流动性越小。微粘度变化的数据为每组各动物 3次所测结果的平均值。
1. 2. 7 红细胞溶血度的测定
各处理组红细胞溶血度的测定参照文献[15]的方法。即取各处理组大鼠血清,用分光光度法在 415 nm 处测定其吸光度; 正常大鼠压积红细胞放入 37 ℃蒸馏水中( 体积比为 1∶ 1) 1 h后,离心取上清,用分光光度法测定该上清吸光度。用红细胞在蒸馏水中溶血后吸光度( 假设这时红细胞的溶血度为 100%) 作对比算出各处理组红细胞的溶血度。各处理组红细胞溶血度变化的数据为每组各动物 3 次所测结果的平均值。
1. 2. 8 红细胞膜蛋白
SDS-聚丙烯酰胺凝胶电泳( SDS-PAGE) 按照 Laemmli 法进行[16],即取红细胞膜样品,经裂解后释放膜蛋白,用 PBS 缓冲液稀释到上样蛋白浓度为 1. 0 mg/mL ( Lowry 法定量) .以7. 5% 的丙烯酰胺胶浓度,150 V 恒压 60 min 电泳后终止。凝胶以考马斯亮蓝( R -250) 染色显带。
1. 2. 9 数据分析
本研究中给出的所有数据为平均值 ± 标准差。不同组间的比较采用 SPSS13 统计软件进行方差分析( ANOVA) ,并用 Origin7. 5 软件进行统计绘图。P <0.05 代表统计学上有显著差异,P <0. 01代表统计学上有极显著差异。
2 结 果
2. 1 各不同处理组红细胞扫描电镜观察
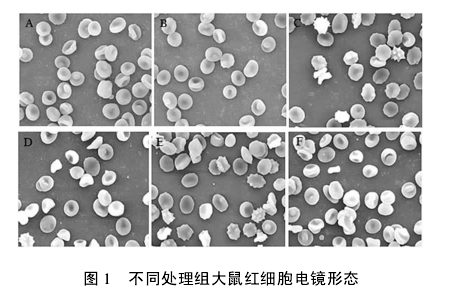
红细胞在受到环境异常刺激后,会出现各种不同异常形态。各处理组红细胞按要求制备成样品后,在扫描电镜下观察红细胞的形态。结果如图 1( 80 000 倍放大) .显然,丙二醛和过度训练导致了大鼠红细胞形态的异常,而 Tau 对红细胞形态的破坏有保护作用。
图 A、B 为分别用自来水和 PBS 喂食的大鼠红细胞,形态上为典型的双凹圆饼状; 图 C 为喂食 MDA 大鼠的红细胞,部分红细胞出现异常结构; 图 D 为喂食Tau 和 MDA 大鼠红细胞,基本上为结构异常的红细胞,少量红细胞周围出现毛刺; 图 E 为过度训练大鼠红细胞,不仅结构异常,还出现聚集现象; 图 F 为过度训练大鼠同时喂食 Tau 后的红细胞,结构大多完整,且为双凹圆饼状。
2. 2 各不同处理组大鼠红细胞膜流动性的变化
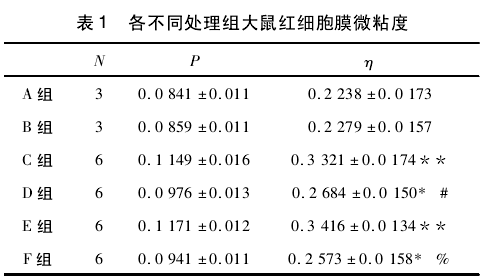
红细胞膜的流动性是红细胞发挥其运载功能的重要生理基础。红细胞膜的流动性以红细胞膜的微粘度来表示,红细胞膜微粘度升高,表明其流动性降低,红细胞变形性下降,是红细胞膜受损的重要指标。各不同处理组大鼠红细胞微粘度 η 的变化如表 1 所示。
与对照组( A 和 B) 比较,其余各不同处理组大鼠红细胞微粘度升高,流动性降低,且都表现出显著或极显著差异。C 组大鼠喂食 MDA 后导致红细胞微粘度升高,流动性降低,与同时喂食 MDA 和 Tau 的 D 组大鼠比较,D 组大鼠红细胞膜微粘度降低,流动性升高,它们之间存在显著差异( #P < 0. 05) ; E 组大鼠在剧烈运动后其微粘度升高,与剧烈运动同时喂食 Tau的 F 组比较,后者的微粘度降低,细胞膜流动性升高,它们之间也表现出了显著差异( %P <0.05) .
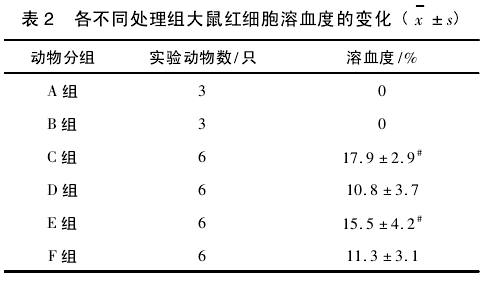
2. 3 各不同处理组大鼠红细胞溶血度的变化在红细胞处于应激状态下,红细胞膜的结构会发生改变,导致膜的完整性变差,红细胞膜容易破裂导致红细胞易于溶血。各不同处理组大鼠红细胞的溶血度变化如表 2 所示。
与对照组( A 组和 B 组) 比较,各种不同处理都导致红细胞溶血度升高,但不管是喂食 MDA 组后再喂食 Tau( C 组与 D 组比较) 或力竭运动后再喂食 Tau( E组与 F 组比较) ,都可显著降低红细胞的溶血度,显示Tau 能拮抗 MDA 或力竭运动所致的红细胞膜的损伤。
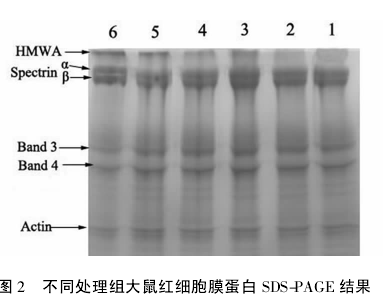
2. 4 各不同处理组大鼠红细胞膜蛋白凝胶电泳结果为进一步研究各不同处理对大鼠红细胞膜蛋白的影响,用生理盐水洗涤各组的压积红细胞后,按1.2. 5 的方法制备红细胞膜,红细胞膜用蛋白裂解试剂盒裂解后,经 SDS-PAGE 凝胶电泳,结果如图 2 所示。泳道 1~ 6 分别为 A ~ F 组大鼠红细胞膜蛋白在 SDS-PAGE中电泳的行为,泳道 1 和2 没有出现新的电泳条带,而其它各处理组都出现了高分子量聚集的新电泳条带( high molecular weight aggregation,HMWA) ,说明各种不同的应激都将导致红细胞膜蛋白结构的变化。
3 讨 论
MDA 作为自由基介导的脂质过氧化和非酶糖基化过程所产生的 α-、β-不饱和醛酮类物质的代表性中间产物[17],其对机体正常生理功能和稳态的破坏作用已被越来越多的科学工作者所重视[18 -20],甚至认为MDA 是导致机体运动性疲劳的重要“疲劳因子”[20].
牛磺酸作为一种非蛋白氨基酸,在临床医学和运动营养学上广泛用于调节机体正常生理功能和增强机体对抗各种应激状态的能力,以人和动物为模型对 Tau在抗运动性疲劳的作用和对红细胞 ATPase 活性的保护作用已有广泛的研究[9,22].然而 Tau 在抗疲劳中的生化机制并不十分明确。
机体在剧烈运动或大负荷长时间训练后,都会导致体适能的下降,而红细胞运载功能的下降与机体这种体适能的下降是密不可分的。由于红细胞长时间处于高氧分压环境,在剧烈运动或大负荷长时间的训练后,活性氧会引发红细胞膜脂成分的脂质过氧化,脂质过氧化反过来又会产生新的活性氧 自由基( ROS) ,在该过程中会产生大量的 MDA 等不饱和活性醛酮类物质[17,20],如此循环往复即脂质过氧化的放大过程加大对红细胞膜结构的损伤。在本研究中,在大鼠喂食 MDA 后红细胞形态发生了改变( 图 1C) ; 在红细胞膜流动性的实验中同样 MDA 喂食大鼠红细胞膜微粘度升高( 表 1C 组) ,且与对照组之间有极显著差异; 在红细胞溶血度实验中,MDA 喂食大鼠红细胞膜溶血度升高( 表 2C 组) ,与对照组间有显著差异。
而 Tau 的喂食,则在很大程度上能逆转 MDA 的这些作用( 图 1D,表 1D 组和表 2D 组) ,表现了 Tau 的抗MDA 作用。已有的研究表明,在人或动物运动性疲劳时 MDA 的水平也显著升高[23 -24],升高的 MDA 会攻击细胞膜上的亚油酸、亚麻酸、花生四烯酸等不饱和脂肪酸中的不饱和双键,进一步破坏细胞膜的脂双层结构,MDA 的这种作用可能是导致运动性疲劳的重要原因之一。从形态上观察,在 MDA 的作用下和过度训练大鼠,其红细胞形态都明显异常,出现了许多非典型形态的红细胞( 图 1C、1E) ; C 组和 E 组大鼠在红细胞膜流动性降低和红细胞的溶血度升高,在喂食 Tau作用后,都能逆转对红细胞的损伤,提示二者在导致红细胞膜结构破坏上可能存在共同的生化机制。在喂食 MDA 和 Tau 的 D 组及过度训练同时喂食 Tau 的F 组,其红细胞膜的结构明显得到保护,与相应的对照组比较红细胞膜的流动性升高,其溶血度也降低。杨爱华等[9]的研究也表明,Tau 的补充能明显延长动物力竭运动时间,其中的生化机制可能与 Tau 抗 MDA 的作用密切相关。
红细胞膜的蛋白成分,主要有细胞骨架蛋白( 血影蛋白、肌动蛋白、原肌球蛋白等) 、血型糖蛋白、离子通道蛋白和转运蛋白( 如 Na-K-ATPase) 等,其在细胞膜中含量有高有低,分子量有大有小。由于 MDA 的分子结构中含有 2 个醛基的双功能基团,能与蛋白质中的 α-NH2以形成 Shiff's 碱形式共价加成,在分子结构上会导致蛋白质间 ( 或者蛋白质内) 的分子交联[25 -26],因此在 SDS-PAGE 电泳条带上能产生大分子量的新的电泳条带。图 2 清楚地显示,喂食 MDA 组大鼠和过度训练疲劳大鼠红细胞蛋白电泳中都出现了高分子量条带 ( high molecular weight aggregation,HMWA) ( 图 2 条带 3 和条带 5) .这些 HMWA 条带的形成是红细胞膜上的各种蛋白质在丙二醛的交联作用下,它们之间以 Shiff's 碱形式共价加成聚集而成的大分子物质,因此在 SDS-PAGE 电泳图上显示出新的条带。而在喂食 Tau 后,这些的新的 HMWA 条带蛋白聚集量减少,推测由于对红细胞蛋白的损伤减少,显示了 Tau 在拮抗由于 MDA 作用或过度训练所导致的对红细胞膜蛋白损伤的保护作用。
4 结 论。
不管是喂食 MDA 或过度训练都可导致红细胞形态和结构的损伤,从而降低红细胞的运载功能,这是导致大鼠运动能力下降和运动性疲劳的原因之一。Tau 的补充则可对红细胞膜起保护作用,从而提高运动能力,提示 Tau 在抗运动性疲劳中的生化机制可能是通过拮抗 MDA 对红细胞的损伤而发挥作用。
参考文献:
[1] Bouckenooghe T,Remacle C,Reusens B. Is taurine afunctional nutrient[J]. Curr Opin Clin Nutr Metab Care,2006,9( 6) : 728 - 33.
[2]Aruoma OI,Halliwell B,Hoey BM. The antioxidant actionof taurine,hypotaurine and their metabolic precursors[J].Biochem J,1988,256,251 - 255.
[3] Galloway SD,Talanian JL,Shoveller AK,et al. Sevendays of oral taurine supplementation does not increasemuscle taurine content or alter substrate metabolism duringprolonged exercise in humans[J]. J Appl Physiol,2008,105,( 2) : 643 - 151.[4]魏源,李良鸣,王步标,等。 牛磺酸对运动小鼠骨骼肌自由基代谢的影响[J]. 中国运动医学杂志,2000,19( 2) : 225 -226.
[5]Dawson RJr,Biasetti M,Messina S,et al. The cytoprotec-tive role of taurine in exercise-induced muscle injury[J].Amino Acids,2002,22 ( 4) : 309 - 324.
[6]Esterbauer H,Schaur RJ,Zollner H. Chemistry and bio-chemistry of 4 - hydroxy-nonenal,malondialdehyde andrelated aldehydes [J]. Free Radic Biol Med,1991,11( 1) : 81 -128.
[7]李芳序。 羰基应激抑制红耳龟的脑活动[D]. 长沙: 湖南师范大学,2008.[8]张宜龙,陈吉棣,刘晓鹏,等。 牛磺酸对人体自由基代谢水平和运动能力的影响[J]. 中国运动医学杂志,1999,18( 1) : 73 - 75.
[9]杨爱华,申伟华,汤华,等。 牛磺酸对丙二醛诱导疲劳大鼠的保护作用[J]. 北京体育大学学报,2011,34( 3) : 71 -74.
[10]蔡建光。 运动性疲劳与衰老过程中的羰基应激[M].吉林: 吉林大学出版社,2013.
[11]Kikugawa Y,Machida M,Kida T,et al. Study on peroxi-dized lipids Ⅲ: Fluorescent pigments derived from the re-action of malonaldehyde and amino acids [J]. ChemPharm Bull,1981,29( 11) : 3003 – 3011.
[12] Dodge JT,Mitchell C,Hannahan DJ. The preparationand chemical characteristics of hemoglobin-free ghosts ofhuman erythrocytes [J]. Arch Biochem Biophy,1963,100( 1) : 119 - 130.
[13]张龙翔,张庭芳,李令媛,主编。 生化实验方法和技术[M]. 北京: 人民教育出版社,1981.
[14] Shinitzky M,Barenholz Y. Fluidity parameters of lipidregions determined by fluorescence polarization [J].