基因工程论文


兰尼定受体( Ryanodine Receptors)在肌肉收缩中起关键作用。兰尼定受体是细胞内质网膜上介导细胞内钙信号转导的离子通道.兰尼定受体是由分子量为560 kDa 的相同亚单位组成的四聚体,整个蛋白质的分子量超过 2000 kDa,是目前已知最大的膜蛋白。
兰尼定受体基因的突变可以导致人和动物的一些病症,例如恶性高热症( Malignant Hyperthermia).人类的骨骼肌型兰尼定受体基因( RYR1) 中已经发现 22 个能导致恶性高热症的突变.猪的恶性高热症( 又称为猪应激综合症) 表现为猪受到应激刺激出现肌肉紧张、呼吸促迫、心跳加快,体温升高,严重者可致突然死亡。这种猪宰后会产生肉色苍白、质地松软、切面渗出的劣质肉.研究证实,猪对恶性高热症的易感性是由于主要原因是骨髂肌型兰尼定受体( RYR1) 基因的突变导致的。猪 RYR1 基因的 1843 碱基处发生的 C→T 碱基突变,使兰尼定受体由第 615 位的精氨酸变为半胱氨酸,从而导致结构和功能的改变。这样使应激状态下 Ca2 +大量释放,引起电解质代谢紊乱,肌肉持续收缩,同时引起肌肉 pH 值异常变化,导致猪由正常的应激抵抗转变为对应激敏感,进而引发了繁殖、生长、胴体和肉质性状等发生全方位的变化.因此,在猪的选育工作中,已经广泛应用兰尼定受体基因作为筛选良种的指标之一。
目前许多脊椎动物的兰尼定受体基因已经被克隆,无脊椎动物如果蝇、线虫和海胆等的兰尼定受体基因也已经被克隆。近期笔者课题组克隆了凡纳滨对虾( Litopenaeus vannamei) 的兰尼定受体基因.本研究旨在查找凡纳滨对虾兰尼定受体基因的 SNP 位点,并进行 SNP 与对温度变化敏感性的关联分析,结果有助于了解对虾应激易感性的分子机制。
1 实验材料与方法
实验选用无特定病原( SPF) 凡纳滨对虾的平均体重大约为 3 g,来自广西水产研究所良种场。
1. 1 温度变化刺激试验 凡纳滨对虾共 192 尾,暂养于塑料桶的海水中,水温 25 ℃。养殖 24 h 后,迅速转入 38 ℃水温的塑料桶中,观察对虾的行为反应。出现痉挛现象的对虾归入应激敏感组,不出现痉挛现象的对虾归入应激抵抗组。随后,用 DNA 提取试剂盒( 天根生物技术公司) 提取各对虾的基因组 DNA.
1. 2 单核苷酸多态性( SNP) 位点的查找及基因频率分析 根据笔者前期获得的凡纳滨对虾兰尼定受体基因序列( Genbank: HM367069) ,使用 Primer Premier 5.0软件设计 1 对引物: 5' - ATCATTGTTGCTGTTATTGT-TATTAT - 3'; 5' - TCTTCTTCGTCTTCACTGAGGTACTT- 3',预计扩增 538 bp 基因片段。PCR 反应体系包含50 ng 基因组 DNA,12. 5 μL Taq Master Mix( 天根生物技术公司) ,1 μL 正向引物( 浓度 10 μmol/L) ,1 μL 反向引物( 浓度 10 μmol/L) ,补充无菌纯水至 25 μL.反应条件为: 先 94 ℃变性3 min; 然后94 ℃ 30 s,60 ℃ 30s,72 ℃ 30 s,共 35 个循环; 最后 72 ℃ 延伸 10 min.随机取 24 个对虾样品基因组 DNA 进行 PCR 扩增,扩增产物经胶回收纯化后,进行双向测序( 由天根生物技术公司完成) ,用 DNAMan 软件比对测序结果,查找 SNP位点。发现 SNP 后,用同样的引物对 192 尾凡纳滨对虾基因组 DNA 进行 PCR 扩增,然后进行单向测序,根据测序图谱判断 SNP 位点的基因型,统计各组基因型频率以及等位基因频率,然后用卡方检验其是否符合哈迪 - 温伯格平衡定律( Hardy - Weinberg Equilibri-um,HWE) .用 SPSS 13. 0 软件对各组对虾的基因型频率和等位基因频率进行单因素方差( ONE - WAYANOVA) 分析.
2 结果与分析
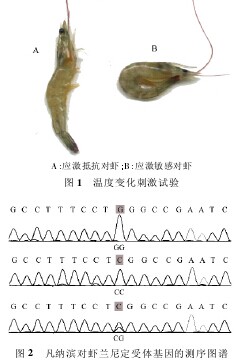
2. 1 温度变化刺激试验 试验观察到,当凡纳滨对虾从 25 ℃海水中被迅速转移到 38 ℃ 海水中后,所有的凡纳滨对虾都快速游动、腹部收缩、弹跳和身体失去平衡。18 尾凡纳滨对虾被观察到不可逆转的尾痉挛并且很快死亡( 图 1 - B) ,被归入应激敏感组。而其余的凡纳滨对虾( 174 尾) 尾痉挛症状较温和并且可以恢复正常( 图 1 - A) ,被归入应激抵抗组。随后,分别提取所有的凡纳滨对虾基因组 DNA 用来检查 SNP 及其基因型。
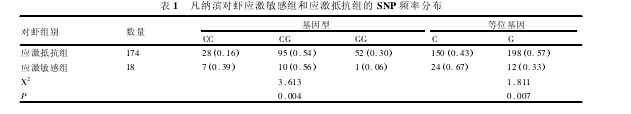
2. 2 SNP 检测及基因频率分析 首先对随机抽取的24 个对虾样品进行 PCR 扩增兰尼定受体基因片段,扩增产物纯化后直接进行双向测序,用 DNAMan 软件比对测序结果,结果发现了 1 个 C/G SNP( 图 2) ,位于第409 碱基的位置,是 Leu→Leu 同义突变。随后,同样用直接单向测序的方法对 192 尾凡纳滨对虾进行 SNP 基因型的检测。用 Chromas 软件查看测序图谱,根据图谱人工判断基因型,单峰为纯合基因型,套峰为杂合基因型( 图 2) ,统计计算应激敏感组和应激抵抗组的基因型频率和等位基因频率( 表 1) .卡方检验结果表明应激敏感组和应激抵抗组的基因型频率分布都符合哈迪 - 温伯格平衡定律。对应激敏感组和应激抵抗组对虾基因型频率单因素方差分析结果 P =0. 004,等位基因频率单因素方差分析结果 P =0. 007,说明两组间的基因型频率及等位基因频率差异显着。【图1.图2】
3 讨论
凡纳滨对虾是世界最重要的对虾养殖品种之一.以往凡纳滨对虾的研究主要集中在生物学和生态学方面。近年来,越来越多的学者对凡纳滨对虾进行分子生物学方面的研究.和其它动物一样,对虾受到物理或化学因子的刺激会产生应激反应。例如,当环境温度剧烈变化时,对虾会产生应激反应,应激敏感的对虾全身或局部收缩,尾部弯曲后大部分不能自由伸展,腹肌白浊,甚至会死亡.此外,高温、强光、过度拥挤、捕捞、换水、受惊吓等外部条件刺激等也常常会引起对虾的应激反应.但是,通常产生应激反应的对虾只占一部分比例,而另一部分对虾不会出现应激反应,其原因目前还没有研究报道。
为了研究对虾应激反应易感性与兰尼定受体基因的相关性,笔者首先在凡纳滨对虾的兰尼定受体基因片段中查找 SNP 位点。PCR 产物直接测序是检测未知 SNP常用的方法,具有方便、快捷和准确等优点.随着测序成本的降低,直接测序法也经常用于已知 SNP 的基因型检测。
本研究在凡纳滨对虾兰尼定受体基因序列中发现了 1 个 SNP 位点,是外显子中的同义突变,它没有改变编码的氨基酸,没有对兰尼定受体的生理功能直接产生影响,但是由于它和兰尼定受体基因可能存在的错义突变位点在染色体上距离很近,通常会连锁遗传,因此可以作为连锁标记进行性状关联分析。如果将来的研究发现更多的 SNP 位点,用多个 SNP 的单倍型进行连锁分析则结果更加稳定、可靠。当然,最好是能找到直接影响应激敏感性状的错义突变位点。然后,笔者进行了凡纳滨对虾的温度变化刺激试验,观察对虾的应激反应。比较分析应激敏感组和应激抵抗组对虾的SNP 频率分布,结果表明: 应激敏感组和应激抵抗组基因型及等位基因频率分布差异显着( P <0.01) ,说明试验对虾的应激敏感性与兰尼定受体基因的多态性相关。因此可以推测,应激敏感的对虾兰尼定受体基因中可能存在改变氨基酸序列的错义突变点而导致对应激的敏感性,这有待于将来在全长兰尼定受体基因序列中查找突变点和进行更多的应激试验和关联分析来证实。【表1】
参考文献:
[1]Franzini - Armstrong C,Protasi F. Ryanodine receptors of stri-ated muscles: a complex channel capable of multiple interac-tions[J]. Physiol Rev,1997,77( 3) : 699 - 729.
[2]Wang R W,Zhang L,Bolstad J,et al. Residue Gln4863 withina predicted transmembrane sequence of the Ca2 +release channel( ryanodine receptor) is critical for ryanodine interaction[J]. JBiol Chem,2003,278: 51557 - 51565.
[3]Loke J,MacLennan D H. Malignant hyperthermia and centralcore disease: disorders of Ca2 +release channels[J]. Am JMed,1998,104: 470 - 486.
[4]Jurkat - Rott K,McCarthy T,Lehmann - Horn F. Genetics andpathogenesis of malignant hyperthermia[J]. Muscle Nerve,2000,23( 1) : 4 - 17.
[5]Mitchel G,Heffron J J A. Porcine stress syndromes[J]. AdvFood Res,1982,28: 167 - 230.
[6]Fujii J,Otsu K,Zorzato F,et al. Identification of a mutation inthe porcine ryanodine receptor that is associated with malignanthyperthermia[J]. Science,1991,253( 5018) : 448 - 451.
[7]Zhao F Y,Li P,Chen S R Wayne. Dantrolene inhibition of ry-anodine receptor Ca2 +release channels [J]. J Biol Chem,2001,276( 17) : 13810 - 13816.