园艺学论文


全世界约有蔷薇属(Rosa L.)植物 200 多个种和变种,广泛分布于亚、欧、北非、北美等寒温带至亚热带地区。中国分布有 95 种蔷薇,分为 2 个亚属 10 个组(Ku & Robertson,2003),是重要的月季种质资源。进行准确的核型分析是将其有效应用于月季遗传改良的前提。由于蔷薇属植物染色体较小,缺乏明显可见具有随体的染色体(李懋学和张赞平,1996;Crane & Byrne,2003;蹇洪英 等,2009),传统的核型分析难以做到准确的同源配对。以核糖体 RNA 基因(rDNA)为探针的染色体荧光原位杂交是对染色体较小且形态相近的物种进行染色体分析的有效手段(Liu & Wendel,2003;徐延浩 等,2007)。rDNA-FISH 应用于蔷薇属植物的研究已有一些报道(Ma et al.,1997;Fernàndez-Romero et al.,2001;Akasaka et al.,2002,2003;Lim et al.,2005;田敏 等,2012,2013a,2013b;Jian et al.,2013b)。田敏等(2012,2013a,2013b)和 Jian 等(2013b),采用荧光原位杂交技术检测了 45S rDNA 在部分野生种、古老月季品种及现代月季品种中数量和信号强弱的差异,但中国应用荧光原位杂交技术对蔷薇属植物进行研究尚属起步阶段。
多苞蔷薇种内具有丰富的表型变异(中国农业科学院中国植物志编辑委员会,1985),川滇蔷薇耐旱和耐贫瘠(周志琼 等,2009),金樱子具有较好的抗虫性(魏开炬 等,2013),目前尚未见rDNA 在多苞蔷薇和川滇蔷薇染色体上定位的研究;Ma 等(1997)仅报道了金樱子的染色体组有 1对 45S rDNA 位点,并未进行核型分析。本研究中利用双色荧光原位杂交技术对 45S 和 5S rDNA 在多苞蔷薇、川滇蔷薇和金樱子的体细胞染色体上进行定位,为这 3 种蔷薇属植物的染色体识别提供明确有效的标记,为进一步利用这 3 种蔷薇对现代月季进行遗传改良提供分子细胞遗传学背景资料。
1、 材料与方法
1.1 材料
材料分别为桂味组(Section Cinnamomeae)的多苞蔷薇(Rosa multibracteata Helm. et Wils.)、合柱组(Section Synstylae)的川滇蔷薇(R. soulieana Crép.)及金樱子组(Section Laevigatae)的金樱子(R. laevigata Michx.),均保存于云南省农业科学院花卉研究所的月季种质资源圃中。本试验于2012 年 8 月采集嫩芽预处理后进行染色体制片,原位杂交试验于 2013 年 1 月完成。
1.2 探针标记
45S rDNA 克隆自番茄,质粒由武汉大学生命科学学院李立家教授提供,用 EasyPure PlasmidMiniPrep Kit 提取(北京全式金生物技术有限公司),以生物素(Biotin-16-dUTP)用缺口平移法标记(Roche,Biotin-Nick Translation Mix,No.11745824910);5S rDNA 以 PCR 方法获得,参照 Akasaka等(2002)的方法制备,并以 PCR 产物纯化试剂盒纯化(上海杰瑞生物工程有限公司),以地高辛(Dig-11-dUTP)用缺口平移法标记(Roche,Dig-Nick Translation Mix,No. 11745816910),按说明书的方法操作。
1.3 染色体制片、原位杂交、图像检测及分析
染色体制片参照 Ma 等(1997)的方法。原位杂交流程参照 Akasaka 等(2003)的方法进行。
杂交图像在 Zeiss 荧光显微镜(Axiophot 2)下观察并获取,用 Zeiss 系统软件及 Photoshop 软件对图像进行处理分析。核型分析采用李懋学和陈瑞阳(1985)的标准,核型类型根据 Stebbin(1971)的分类标准划分,染色体排序参考 Jian 等(2013a)的常规核型分析的结果排列。
2、 结果与分析
2.1 多苞蔷薇 45S rDNA 和 5S rDNA 的双色荧光原位杂交
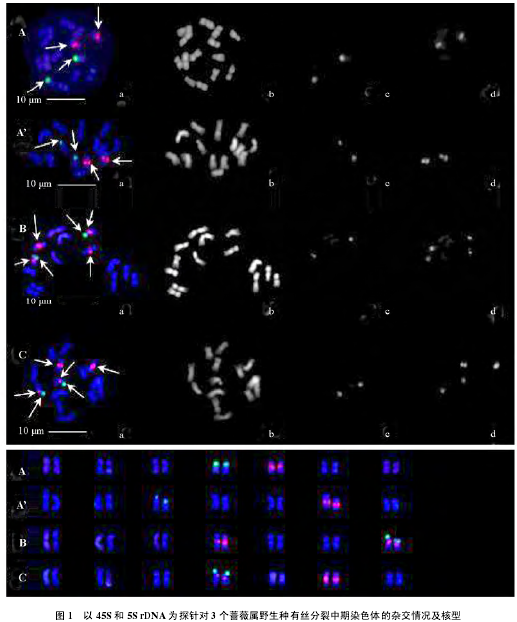
以 45S rDNA 和 5S rDNA 为探针的 3 种蔷薇属植物的荧光原位杂交结果及核型见图 1 和表 1。
45S rDNA 和 5S rDNA 在多苞蔷薇上(图 1,A、A’)各检出 1 对位点。本试验中发现多苞蔷薇有两种核型,一种(图 1,A)核型公式为 2n = 2x = 14 = 12m + 2sm,核型为 1A 型,除 4 号染色体为亚中部着丝点染色体(sm)外其它都是中部着丝点染色体(m),此类型中 45S rDNA 定位在 4号染色体的短臂上(S),1 个杂交信号比另 1 个稍弱;5S rDNA 位于 5 号染色体长臂(L)的近着丝点端,1 个杂交信号比另 1 个稍弱。另一种类型(图 1,A’)核型公式为 2n = 2x = 14 = 10m + 4sm,核型为 2A 型,除 3 号和 6 号为亚中部着丝点染色体(sm)外,其余均为中部着丝点染色体(m),而且 3 号和 6 号都是异形同源染色体,此类型中 45S rDNA 定位在 3 号染色体的短臂上(S),杂交信号较长染色体的稍弱;5S rDNA 位于 6 号(sm)染色体的长臂(L)的近着丝点处,杂交信号较长染色体的稍弱一些。
2.2 川滇蔷薇 45S rDNA 和 5S rDNA 的双色荧光原位杂交
45S rDNA 和 5S rDNA 在川滇蔷薇上(图 1,B)分别检出 1 对和 2 对位点。基于 rDNA-FISH的川滇蔷薇核型公式为 2n = 2x = 14 = 10m + 4sm,核型为 2A 型,4 号和 7 号染色体为亚中部着丝点染色体(sm),其余为中部着丝点染色体(m),其中 7 号为异形同源染色体。45S rDNA 位于 7 号染色体的短臂上(S),较短染色体的信号稍弱一些。5S rDNA,1 对位于 4 号染色体长臂(L)的近着丝点端,1 个杂交信号较另 1 个强很多;另 1 对位于 7 号染色体长臂(L)的近着丝点端,较短染色体的信号稍弱一些,其信号强度介于 4 号染色体两个信号的强度之间。
2.3 金樱子 45S rDNA 和 5S rDNA 的双色荧光原位杂交
45S rDNA 在金樱子上(图 1,C)检出 1 对位点。5S rDNA 检出 2 对位点。基于 rDNA-FISH的金樱子核型公式为 2n = 2x = 14 = 10m + 4sm,核型为 2A 型,4 号和 5 号为亚中部着丝点染色体(sm),其余为中部着丝点染色体(m),其中 4 号为异形同源染色体。45S rDNA 位于 4 号染色体的短臂上(S),杂交信号强弱一致;5S rDNA 1 对位于 4 号染色体的长臂(L)的近着丝点处,另 1对位于 6 号染色体长臂(L)的近着丝点处,位于 4 号染色体的 1 对杂交信号比 6 号染色体的弱。

3、 讨论
本研究中基于 rDNA-FISH 的核型分析发现多苞蔷薇有两种核型类型,一种与 Jian 等(2013a)基于常规核型分析的结果相同,属较为原始的 1A 型,另一种为更进化的 2A 型(Stebbins,1971)。
前者没有异形同源染色体,且 5S rDNA 位点染色体不是 sm 染色体;后者的两对 rDNA 染色体都是异形同源染色体。本课题组保存的多苞蔷薇来自 3 个居群,本研究中发现其具有两种核型,可能是由于两种核型的多苞蔷薇来自不同居群,它们之间存在核型差异,需要对不同居群的多苞蔷薇分别取样研究。这也从细胞水平上反映了为何多苞蔷薇种内形态变异丰富(中国科学院中国植物志编辑委员会,1985)。唐开学(2009)认为川滇蔷薇种内存在着丰富的居群间和居群内表型变异。本研究中川滇蔷薇核型及核型公式与 Jian 等(2013a)基于常规核型分析的结果相同,但两对 sm 染色体的序号相差较大,这可能是由于研究材料来源不同所致。金樱子在核型和核型公式上与 Crane 和 Byrne(2003)及 Jian 等(2012)基于常规核型分析的结果相同,但在染色体排序上稍有差别。因此,蔷薇属植物不仅在种间存在丰富的核型多样性(唐开学,2009;Jian et al.,2013a),种内也存在丰富的变异,应采用荧光原位杂交等较精确的方法对一些分布较广、种内存在丰富形态变异的蔷薇进行进一步的核型研究。本研究中还发现这 3 种蔷薇属野生种均存在异形同源染色体,它们分别是多苞蔷薇(图 1,A’)3 号和 6 号染色体、川滇蔷薇 7 号染色体和金樱子 4 号染色体,异形同源染色体在相对长度上存在明显差异,由于较大的长度差异使研究人员在通过常规染色体核型分析时很容易将其错配。可见基于 rDNA-FISH 的蔷薇属植物染色体核型分析更加准确可靠。
核糖体 RNA 基因(ribosomal DNA,rDNA)是具有重要功能的保守重复序列,成簇分布于 1对或多对染色体上(Pedersen & Linde-Laursen,1994)。45S rDNA 被认为在进化过程中相当保守,与核仁形成有关,一般在染色体的次缢痕部位,也可存在于非次缢痕位点(徐川梅 等,2007;轩淑欣 等,2007;徐晶,2009),李懋学和张赞平(1996)也指出,次缢痕既 NOR,但 NOR 不一定在次缢痕区。蔷薇属植物的染色体很小,在常规染色体分析中看不到次缢痕,无法识别核仁组织区,也很难识别其同源染色体。Akasaka 等(2002,2003)研究 rDNA 在蔷薇属 9 个二倍体野生种的分布中发现 45S rDNA 均定位在 1 对异形同源 sm 染色体短臂的近端部。本研究中多苞蔷薇、川滇蔷薇和金樱子都只有 1 对 45S rDNA 位点,大都位于 1 对异形同源 sm 染色体的短臂上。一般认为,随体是高度惰性的异染色质,着丝点附近区域亦是异染色质集中分布的区域。对其他物种的研究表明,45S rDNA 位点可能位于长臂或短臂上,但基本分布于近着丝点处(轩淑欣 等,2007)。Fransz 等(2002)认为,在间期核中常染色质与异染色质都是具有特定空间结构的,从而顺利实现基因的表达调控。那么蔷薇属植物中 45S rDNA 位点分布于 1 对异形同源 sm 染色体的短臂上应当与其特定功能有关。目前已有的报道(Ma et al.,1997;Fernàndez-Romero et al.,2001;Akasaka et al.,2002,2003;Lim et al.,2005;田敏 等,2012,2013a,2013b;Jian et al.,2013b)表明有 20 个蔷薇属二倍体野生种的 45S rDNA 位点数与其染色体倍性相同,即 1 个染色体组有 1 个 45S rDNA 位点,但卡罗莱纳组(Sect. Carolinae)的 R. foliolosa(2n = 2x = 14)有 6 个 45S rDNA 位点,分别位于 3 对sm 染色体短臂的端部,可见在蔷薇属二倍体野生种中的 45S rDNA 数存在明显差异。需要积累更多野生蔷薇 rDNA 的研究资料以便探讨它们的系统关系。
5S rRNA 是核糖体大亚基的组成成分之一,每个 5S rDNA 单元包括 120 bp 左右的转录区和 100 ~700 bp 的非转录区(Appels & Baum,1992)。Mantovani 等(2005)发现在大部分已进行了 5S rDNA定位的物种中,其数量以 1 对或 2 对为主,但在染色体上没有固定的分布模式。Akasaka 等(2002;2003)在研究中发现,蔷薇属 9 个二倍体野生种中 5S rDNA 位点数有 2 个(1 对)、3 个和 4 个(2对),其中只有 1 对的种的 5S rDNA 位点位于非 45S rDNA 染色体长臂的近着丝点处;有 3 个或 2对的种其 5S rDNA 有 1 对与 45S rDNA 位于同一染色体的长臂或短臂上,其余位点位于另外染色体的长臂的近着丝点处。Lim 等(2005)的研究表明 5S rDNA 在五倍体狗蔷薇中有 8 个位点均位于染色体长臂的近着丝点处,其中有 3 个位点与 45S rDNA 位于同一染色体,另外 5 个位点位于另外的染色体上。本研究中多苞蔷薇有 1 对 5S rDNA 位于非 45S rDNA 染色体的长臂的近着丝点处,川滇蔷薇和金樱子有 2 对 5S rDNA 位点,其中 1 对位于 45S rDNA 染色体的长臂的近着丝点处,另 1 对位于其它染色体长臂的近着丝点处。可见,蔷薇属植物中 5S rDNA 位点按是否与 45S rDNA 有共线性分为两种:只有 1 对 5S rDNA 位点的与 45S rDNA 没有共线性,且位于 1 对染色体长臂的近着丝点处;有 3 个或 2 对 5S rDNA 位点的,与 45S rDNA 有共线性的 5S rDNA 位点可位于染色体短臂或长臂的近着丝点处,另外的位点位于其它染色体长臂的近着丝点处。
本研究中通过双色荧光原位杂交技术对多苞蔷薇、川滇蔷薇和金樱子 3 种蔷薇野生种的 45S 与5S rDNA 在体细胞中期染色体上进行物理定位,不但能准确地识别各自染色体组中的同源染色体,而且还能通过杂交位点数量、位置和强弱体现各自染色体的结构特征,为这 3 种植物的染色体识别提供了明确有效的标记。rDNA 在蔷薇属植物上的应用已从分子细胞遗传学层面对部分野生种、古老月季及现代月季间的关系作了一些阐述(Ma et al.,1997;Fernàndez-Romero et al.,2001;Akasakaet al.,2002,2003;Lim et al.,2005;田敏 等,2012,2013a,2013b;Jian et al.,2013b),为了更好地将蔷薇野生资源应用于现代月季遗传改良工作中,需要积累更多种蔷薇的 rDNA-FISH 资料。已有的研究也表明以 rDNA 为探针还不能完全区分蔷薇属植物的每对同源染色体,应从蔷薇属植物的基因组中分离出其它特异重复序列作为探针以区分各对同源染色体,这不仅可用于探讨蔷薇属植物的演化,还可在现代月季遗传育种工作中明确染色体的去向,从而提高育种效率。

百合(Liliumspp.)是世界着名切花之一,具有很高的观赏价值,一直深受人们喜爱,是重要的商品花卉和园林绿化植物。自Robb首次离体培养百合鳞茎以来,组织培养技术在百合上的应用已取得了显着的进展,但百合种类繁多,类型各异,目前很多品种尚未形成规模化生...