园艺学论文


番茄(Solanum lycopersicum L.)生产中番茄黄化曲叶病毒为害日趋严重(Czosnek & Ghanim,2011)。番茄黄化曲叶病毒(Tomato yellow leaf curl virus)简称 TYLCV,是单链环状 DNA 病毒(刘玉乐 等,1998),主要依靠烟粉虱进行传播(Stansly et al.,2012)。与传统育种相比,基因工程育种周期短,在育种过程中不会带入其他不良农艺性状的基因,在番茄抗病毒病育种中的应用日益受到重视。通过克隆与抗 TYLCV 有关的基因导入植物体内,有望提高植物的抗病性。
研究表明,R 基因对各种病原菌引发的病害均具有抗性,且在植物基因组中分布广泛(Flor,1971)。根据已克隆的 R 基因所编码的蛋白质保守结构及其在细胞中的位置,可以将 R 基因分为NBS-LRR 类、LRR-TM 类、STK 类、LRR-TM-STK 类和 CC 类抗病基因(Hammond-Kosack & Jones,1997)。NBS-LRR 类基因是已知的 R 基因家族中最大的一类(Dangl & Jones,2001)。比较基因组分析表明植物基因组可以编码数十个到数百个 NBS-LRR 基因,如木瓜和黄瓜的 NBS-LRR 基因有50 个,而水稻则多达 653 个(Shang et al.,2009)。在病毒侵染时,NBS-LRR 基因驱动植物表型反应,以过敏性抗性反应和极端抗性反应两种形式抵御病毒侵染(Hull,2002),且在极端抗性中病毒增殖被限制在单细胞水平,通过局部坏死病变使病毒不能在初始侵染位点扩增(Lukhovitskaya et al.,2005)。NBS-LRR 基因编码的 NBS-LRR 蛋白具有核苷酸结合位点(NBS)和富亮氨酸重复基序(LRR),其中 NBS 包含在信号转导中,具有高度保守且严格有序的多个功能基序(Tan &Wu,2012),LRR 为高度灵活的结构域,可能作为受体参与病原体的识别和信号转导,其激酶结构域在激活其下游转导元件中发挥作用(Jones & Dangl,2006)。目前已知 LRR 结构域通过调控 NBS 结构域的分子状态来发挥抗病功能(Takken et al.,2006)。
基于番茄响应 TYLCV 侵染的转录组结果,作者前期发现了 1 个 NBS-LRR 类抗病基因在抗病番茄材料中上调表达,并通过 VIGS 和在 TYLCV 侵染抗病番茄材料‘CLN2777A’后该基因表达量的变化表明该基因有一定的抗病功能(Chen et al.,2013)。本试验中进一步克隆该 NBS-LRR 类抗病基因 ClNLR,通过荧光定量 PCR 发现 ClNLR 在番茄‘CLN2777A’根和叶片中高水平表达,且通过抗、感番茄材料接种 TYLCV 后 ClNLR 表达量的对比和在本氏烟中的瞬时表达,进一步证明了该基因与番茄抗病性相关。同时再次通过 VIGS 试验对接病前后番茄抗病材料‘CLN2777A’的表型进行了观察,并通过荧光定量 PCR 验证了前期的试验结果。
1、 材料与方法
1.1 试验材料
抗 TYLCV 番茄材料‘CLN2777A’和感 TYLCV 番茄材料‘TMXA48-4-0’由江苏省农业科学院蔬菜研究所提供。携带TYLCV的烟粉虱为连续饲养于防虫温室番茄上的试验种群。pTRV1、pTRV2和 pTRV2-PDS 质粒由清华大学刘玉乐教授馈赠。大肠杆菌 DH5α、根瘤农杆菌 LBA4404、GV3101、质粒 pBI121 均为本实验室保存。表达载体质粒 CaMV35S 由本实验室在 pBI121 的基础上改造而成。
TA 克隆载体 PGEM-T 载体、限制性内切酶(XbaⅠ、BamHⅠ、SacⅠ)、T4DNA 连接酶、DNA 聚合酶、DNA 分子量标准均购自 TaKaRa 公司,其他试剂为国产分析纯试剂。TIANamp Genomic DNAKit、血液/细胞/组织基因组 DNA 提取试剂盒购自 TIANGEN 生物公司,质粒提取试剂盒、PCR 清洁试剂盒、DNA 片段快速纯化/回收试剂盒购于 AXYGEN 公司。引物序列由 Primer 5.0 引物设计软件评价生成,并委托上海英骏公司合成。
1.2 ClNLR 的克隆与序列分析
在前期的研究中,从‘CLN2777A’和‘TMXA48-4-0’番茄材料接种 TYLCV 前后的转录组(NCBIsra 数据库登录号 SRP028618)中发现 1 个响应 TYLCV 侵染而上调表达、注释为含有 NBS-LRR 结构域的抗性基因Solyc05g009760.1,通过初步的功能验证,表明该基因有一定的抗病功能(Chen et al.,2013)。
根据 Solyc05g009760.1 基因序列,用 Primer 5.0 软件设计扩增基因编码区全长的特异引物 P3:5′-TTCACAAGACTTTAGATGTGGCAT-3′及 P4:5′-AGTAGTTCTTGGGGATATGAAGTTG-3′,以抗病材料‘CLN2777A’cDNA 为模板,参照 PrimerSTAR 高保真酶反应体系扩增该基因序列。PCR 反应条件为:94 ℃预变性 3 min;94 ℃ 30 s,58 ℃ 40 s,72 ℃ 2 min 30 s,35 个循环;72 ℃ 10 min。
将回收的 PCR 产物连接至 PGEM-T 载体后转化大肠杆菌 DH5α,转化子经菌落 PCR 鉴定为阳性克隆后送至上海英骏公司测序。所获得的基因序列命名为 ClNLR 基因。用在线软件 Conserved Domains分析保守结构域。
1.3 ClNLR 在 TYLCV 诱导下的表达分析
将番茄材料‘CLN2777A’和‘TMXA48-4-0’以携带 TYLCV 的烟粉虱进行 TYLCV 接种 5 d后,提取叶片 RNA,M-MLV Reverse Transcriptas(eMBI 公司)反转录合成 cDNA。以 Actin(AB199316)为内参,荧光定量 PCR 检测 ClNLR 基因的表达情况。ClNLR 和 Actin 引物分别为 ClNLR-forward:5′-CTTTGCGGGTTCGTTCATCTTAT-3′;ClNLR-reward:5′-CGTTTATGTCCACATGCCTCAAC-3′;Actin-forward:5′-TGGTCGGAATGGGACAGAAG-3′;Actin-reward:5′-CTCAGTCAGGAGAACAGGGT-3′。荧光定量 PCR 反应体系为 20 μL:2× SYBR Premix Ex Taq?Ⅱ10 μL,引物各 1 μL,cDNA模板 1 μL,超纯水 7 μL,混合加样。PCR 反应程序为 96 ℃预变性 1 min,95 ℃变性 15 s,60 ℃退火 15 s,72 ℃延伸 45 s,40 次循环。试验设置 3 个生物重复。
1.4 ClNLR 的组织表达分析
提取番茄抗病材料‘CLN2777A’根、茎、叶片、花蕾和果实 RNA,每个样品取 500 ng RNA,反转录成 cDNA 后稀释 1 000 倍,取 2 μL 进行荧光定量 PCR 反应。基因特异引物、内参引物、荧光定量 PCR 的反应体系和反应程序参照 1.3 的方法。试验进行 3 个生物学重复。
1.5 ClNLR 在本氏烟的瞬时表达
以获得的ClNLR 基因为模板,通过 PCR 扩增在ClNLR基因两端引入BamHⅠ和SacⅠ酶切位点。PCR 引物为 P5:5′-CATGGATCCGGCATTGTTAATCATTAGAGTATG-3′(引入 BamHⅠ酶切位点)和 P6:5′-ACTGAGCTCTTAGTAGTTCTTGGGGATATGAAG-3′(引入 SacⅠ酶切位点)。植物表达载体 pBI121 和 PCR 产物分别经用 BamHⅠ和 SacⅠ双酶切,于 16 ℃连接过夜。取 5 μL 连接产物转化大肠杆菌 DH5α,在固体 LB(附加 50 mg · L-1卡那霉素)培养基上 37 ℃培养过夜,单克隆提取质粒 DNA 并经酶切验证后命名为 pBI121-ClNLR。pBI121-ClNLR 质粒通过冻融法转化农杆菌LBA4404。
将含有pBI121-ClNLR的农杆菌接种于液体LB(附加50 mg · L-1卡那霉素和25 mg · L-1利福平)、28 ℃培养过夜。取培养液离心弃上清,沉淀重悬于缓冲液(10 mmol · L-1MES,10 mmol · L-1MgCl2,200 μmol · L-1乙酰丁香酮)内,调整 OD600值至 0.5,用无针头 1 mL 无菌注射器将农杆菌悬浮液压入5 ~ 6片真叶期的本氏烟的半边叶片,另一半边叶片注射以同样方法获得的pBI121-GFP作为对照。
农杆菌注射后的烟草在培养箱中(22 ℃,75%空气湿度,黑暗)培养 2 d,转入人工气候室培养(28 ℃,16 h/8 h 光照周期),观察表型变化。
1.6 VIGS 沉默 ClNLR 基因
以pBI121-ClNLR为模板,PCR克隆出283 bp的ClNLR基因片段(P7:5′-GTATCTAGAGTGTTGTATGTCCGTGGCTGAC-3′和 P8:5′-CATGGATCCGAGAACCCAATGAC TCCCTGC-3′)。用 XbaⅠ和BamHⅠ酶切 PCR 产物和病毒载体 pTRV2,然后将该基因片段克隆至 pTRV2 中,构建 ClNLR 基因的沉默载体 pTRV2-ClNLR。
质粒 pTRV1、pTRV2、pTRV2-PDS 和 pTRV2-ClNLR 采用冻融法转化农杆菌 GV3101。将转化成功的农杆菌 28 ℃过夜培养,离心弃上清后重悬于缓冲液(10 mmol · L-1MES,10 mmol · L-1MgCl2,200 μmol · L-1乙酰丁香酮)中,调节 OD600值至 2.0,室温放置 4 ~ 6 h 后,将含有 pTRV2-ClNLR、pTRV2-PDS 和 pTRV2 空载体的农杆菌悬浮液分别与含有 pTRV1 的农杆菌悬浮液按 1︰1 的比例混合,用无针头 1 mL 无菌注射器将农杆菌悬浮液注入抗病番茄材料‘CLN2777A’子叶期幼苗的子叶中。其中,注射 pTRV1 + pTRV2 空载体的植株为阴性对照,注射 pTRV1 + pTRV2-PDS 的为阳性对照,番茄幼苗一直在 22 ~ 25 ℃、相对湿度 50%、16 h/8 h 的光照周期下生长。
当注射 pTRV1 + pTRV-PDS 的番茄叶片上出现光漂白现象时,提取 pTRV1 + pTRV2-ClNLR 的番茄上端嫩叶总 RNA;并用 oligo(dT)合成第一链 cDNA,然后通过荧光定量 PCR 检测 ClNLR 基因的沉默效果,定量引物及反应条件参照 1.3。
1.7 ClNLR 基因沉默对番茄体内 TYLCV 含量的影响
选取 ClNLR 基因沉默的番茄植株和注射 pTRV1 + pTRV2 空载体的番茄植株同时置于密闭的防虫温室,烟粉虱接种 TYLCV 后 7 d 时,用 CTAB 法提取番茄叶片总 DNA。用 SHIMADZU 公司的BioSpec-nano/230v 型号的微量分光光度计测定总 DNA 浓度后,将其稀释为终浓度 300 ng · μL-1备用。荧光定量 PCR 检测番茄植株体内的 TYLCV 积累,以 Actin 为内参。其中检测 TYLCV 病毒含量的引物为 TY-AV494:5′-GCCYATRTAYAGRAAGCCMAG-3′和 TY-COPR:5′-GANGSATGHGTRCADGCCATATA-3′,Actin 引物及荧光定量 PCR 的反应条件参照 1.3。
2、 结果与分析
2.1 ClNLR 基因的克隆
作者前期通过比较番茄材料‘CLN2777A’和‘TMXA48-4-0’接种 TYLCV 前后的转录组(NCBIsra 数据库登录号 SRP028618),发现在接种 TYLCV 后,1 个 NBS-LRR 类抗性基因(Solyc05g009760.1)在抗病材料‘CLN2777A’中上调表达,而在感病材料‘TMXA48-4-0’中则无明显变化(Chen et al.,2013)。
本试验中进一步通过 PCR 克隆该基因,鉴于该基因的注释功能为 NBS-LRR 抗病基因将之命名为 ClNLR。结构分析表明 ClNLR 基因编码蛋白在 N 端第 43 ~ 323 位氨基酸为保守结构域 NB-ARC,480 ~721 位氨基酸序列为典型的 LRR 结构域(图 1)。

2.2 ClNLR 基因的表达分析
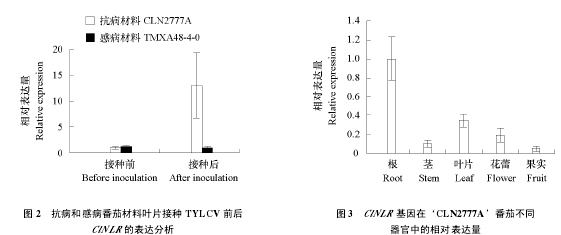
为了验证 ClNLR 基因的转录组表达结果,利用荧光定量 PCR 检测 ClNLR 响应 TYLCV 侵染的表达情况。抗病番茄材料‘CLN2777A’和感病番茄材料‘TMXA48-4-0’通过烟粉虱接种 TYLCV,5 d 后 ClNLR 在抗病材料中表达量相比接种前上调了约 12 倍,而在感病材料中则无明显变化(图 2),基因表达变化趋势与转录组结果(Chen et al.,2013)基本一致。荧光定量 PCR 进一步检测 ClNLR 基因在抗病番茄‘CLN2777A’中的组织表达情况,发现 ClNLR在根部表达最强,其次为叶片,在果实中表达最弱(图 3)。
2.3 ClNLR 瞬时表达引起本氏烟过敏性反应
将含有 ClNLR 基因植物表达载体的农杆菌通过农杆菌浸润法注射本氏烟叶片 15 d 后,注射 ClNLR 基因的一侧叶片上出现了坏死斑,而注射 GFP 基因的另一侧叶片没有任何变化(图 4),表明 ClNLR 基因诱导了本氏烟过敏性反应,可能与植物抗病防卫反应相关。

2.4 沉默 ClNLR 基因影响番茄 TYLCV 抗性
为了研究ClNLR在番茄中抗TYLCV的作用,本试验中通过TRV沉默抗病番茄的ClNLR基因。番茄幼苗注射 TRV 沉默载体 15 ~ 20 d后,注射 pTRV1 + pTRV2-PDS 的阳性对照植株新叶出现光漂白现象,随着时间推移,叶片出现光漂白现象愈明显(图 5),表明 VIGS 沉默成功。

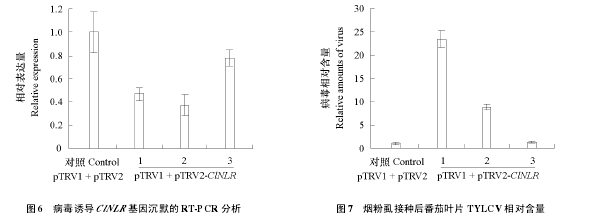
此时提取 pTRV1 + pTRV2-ClNLR 处理的番茄嫩叶总 RNA,通过荧光定量 PCR 检测发现 3 株独立 VIGS 处理的番茄植株 ClNLR 基因的表达水平下降至阴性对照的 40% ~ 80%(图 6)。这些 pTRV1 + pTRV2-ClNLR 处理的番茄通过烟粉虱接种 TYLCV 7 d 后,荧光定量 PCR 检测发现抑制ClNLR基因表达可使番茄体内的TYLCV含量增加,且ClNLR基因表达水平与其体内TYLCV积累量成反比(图 6,图 7),但 ClNLR 基因表达下降的番茄植株接种后没有出现典型的叶片黄化、卷曲等番茄黄化曲叶病病症(图 5),这可能是由于 CLNLR 的沉默效率未能达到使基因表达完全被抑制的水平,低表达的 ClNLR 仍能维持植株一定的抗性。
3、 讨论
植物对病毒的抗性大概可以描述为基因网络中基因的相互作用和信号传递途径抑制了病毒的复制和移动,以及诱导产生的小分子介导了抗性基因的活化(Culver & Padmanabhan,2007)。抵抗TYLCV 的分子和生化机制仍不清楚(Gorovits et al.,2007),因此描绘 TYLCV 的抗性网络并发掘抗性基因是至关重要的。为了达到这个目标,开展了番茄响应 TYLCV 侵染的转录组研究,还发现ClNLR 基因(Solyc05g009760.1)可能参与抵抗 TYLCV 侵染的防卫反应(Chen et al.,2013)。在此基础上,本试验中进一步克隆了该 ClNLR 基因(图 1)。荧光定量 PCR 证实 ClNLR 基因在抗病番茄材料上调表达,而在感病材料中无明显表达变化(图 2),初步说明 ClNLR 基因在抗病番茄中可以响应 TYLCV 的侵染。ClNLR 基因在本氏烟叶片上的瞬时表达可产生过敏性反应(图 4),VIGS 抑制抗病番茄材料 ClNLR 基因的表达提高了 TYLCV 在番茄叶片中的积累量,且番茄 ClNLR 基因表达水平与其体内 TYLCV 积累量成反比(图 5、图 6、图 7),表明该基因可能参与了番茄抗 TYLCV 的防卫反应。ClNLR 基因在番茄的根和叶片中表达最高(图 3),也可能与其抗病功能相关。
ClNLR 编码的蛋白质含有 NBS-LRR 抗病蛋白的特征结构域,其 N 端为 1 个典型的 NB-ARC 结构域,C 端含有 4 个 LRR 保守结构域,是典型的 NBS-LRR 类抗病基因的结构(图 1)。NBS 在细胞的生长与分化、小泡运输、细胞骨架构成及防御反应中都具有重要作用(Pan et al.,2000)。NB-ARC结构域分布于真核生物的许多蛋白中,如 GTP 结合蛋白、ATP 合成酶 β 亚基、核糖体延伸因子(elongation factors)异质三聚体、R 基因编码蛋白等且它在蛋白与蛋白互作中作为一个功能组件发挥作用,比如嘌呤核苷酸的结合通常被认为是改变了 R 蛋白与防御信号传导途径中其它成员间的相互作用(Bent,1996)。而具有特异识别的氨基酸残基大部分位于 LRRs,可形成多种配基结合域作为 R 蛋白的受体区域,配基结合、参与蛋白与蛋白间的互作、蛋白与碳水化合物结合,并能特异性识别病原物来源的信号分子。在对拟南芥 NBS-LRR 类抗病基因 RPS5 的 LRR 突变体研究中发现,它不仅参与信号的识别还参与了下游信号的传导(Wang et al.,1998)。根据 ClNLR 的结构特征,推测该基因可能通过与病原的效应子进行结合而抑制病原在植物细胞中定植和扩展,从而在番茄抗病防御过程中发挥作用。
在缺少相应 Avr 产物的情况下,有时候 R 基因的过表达也会引起 HR 反应。例如番茄的 Pto 基因(Tang et al.,1999),拟南芥的 RPS2 基因(Tao et al.,2000)和烟草的 L 基因(Frost et al.,2004)。
这些数据表明信号级联反应可以在缺少病原物的情况下被激活。这种强烈的过敏性坏死反应在 R 蛋白 RPS2 和 RPS5 中通过 CC 区域和 NB-ARC 片段(CC-NB-ARC)诱导产生(Tao et al.,2000;Juleset al.,2007)。而在 RPS4 和 RPP1A 抗性蛋白中,TIR-NB-ARC 便足以诱导 HR(Zhang et al.,2004;Weaver et al.,2006)。ClNLR 编码的蛋白不包含 TIR 和 CC 结构,哪个结构域引发了 HR 反应以及从 R 蛋白到 HR 免疫反应的信号转导途径仍有待进一步的研究。
病毒诱导的基因沉默已经发展为研究植物基因沉默的有效工具(Purkayastha & Dasgupta,2009)。
如今最常用的一种病毒载体为烟草脆裂病毒(Liu et al.,2002;Brigneti et al.,2004)。Czosnek 等(2013)通过比较抗感番茄材料的 cDNA 文库和 VIGS 的方法,筛选并鉴定出 25 个在抗病番茄材料中优先表达的基因,其中 5 个基因的沉默使抗性瓦解,并指出番茄对 TYLCV 的抗性存在多层次关系,其中SlVRSLip 基因在 LEHT1 基因的下游发挥作用(Sade et al.,2013)。从图 6 和图 7 中可以看到,VIGS抑制 ClNLR 基因表达的 1、2 植株接种 TYLCV 后,带毒量都显著增加,植株 3 的 ClNLR 基因表达水平和接病后的带毒量与对照相差不大,可能是由于该基因在植株 3 中没有被有效沉默。值得注意的是,烟粉虱接种供试番茄植株后,即使显著沉默的番茄也未出现番茄黄化曲叶病病症,可能是由于ClNLR基因的沉默效率不高,未能使基因的表达被完全抑制或抑制到感病的水平,也可能是ClNLR基因只是抗病反应中的下游防卫基因并非抗病关键基因。通过接种其他病原物鉴定该基因是否具有广谱抗性可以更好阐释 ClNLR 基因在抗病反应中的作用。

启动子是调控基因表达过程中重要的转录调节因子,分析启动子时空表达模式可以精确可靠地定位基因的表达部位,对研究基因的时空表达模式和验证基因功能具有重要意义(AyoubivanDeVen,1996;聂丽娜等,2008)。DUF231家族是一类植物特有但功能未知的蛋白...