发育生物学论文


实蕨科( Bolbitiaceae) 是蕨类植物分类系统中出现较晚,且物种相对较少的科,最早是由秦仁昌于1978 年依据孢子体特征,将其从藤蕨科( Lomariopsidaceae) 中独立出来的,在我国仅有实蕨属和刺蕨属,目前,已经被确认的种类为 30 余种,广泛分布于华南及西南地区,不少种类具有药用价值或较高的观赏性。尽管吴兆洪、董仕勇等先后对该科种类进行了修订,肯定了该科在分类系统中的合理性,但是,在实蕨科的分类问题上还一直存有争议,HOLTTUM认为实蕨属、刺蕨属以及舌蕨属应归入舌蕨科( Elaphoglossaceae) ,NAYAr则把它们归入藤蕨科,近代分子系统学则支持将实蕨科包含于鳞毛蕨科( Dryopteridaceae)。配子体发育是真蕨类生活史中进行有性生殖的必经过程,承担着孕育胚的重要使命,因蕴藏着古老的遗传信息而对阐明蕨类科属的亲缘关系具有重要的价值。前人对配子体的发育过程做了大量的研究工作,就实蕨科而言,MOMOSE对华南实蕨成熟配子体做了较细致的观察,NA-YAR对实蕨科某些种的配子体早期发育进行了简要描述,郭严冬对刺蕨属的中华刺蕨和长耳刺蕨的配子体发育进行了研究,但对边缘细胞的观察和假根特征的观察与描述仍不够完善。笔者选取实蕨科中分布广泛、特征典型的多羽实蕨( Bolbitis angustipinna ( Hayata) H. Ito) 为材料,用人工培养法,全面观察了其配子体各发育阶段的形态特征,尤其是假根和边缘细胞的多样性,旨在为深入研究实蕨科配子体发育提供参考。
1、 材料与方法
1. 1 材料 多羽实蕨( Bolbitis angustipinna ( Hayata) H. Ito) 孢子,由张宪春于 2004 - 12 - 25 采自海南七仙岭,标本凭证保存于哈尔滨师范大学植物标本室( HANU) 。
1. 2 方法 在采集地将含有孢子囊尚未开裂的成熟孢子叶羽片剪下,暂时存放于洁净的硫酸纸袋内,密封后放于干燥、无风、阴凉的环境中。3 ~ 4 d 后,孢子囊自然开裂,收集洒落的棕褐色粉末状孢子,置于- 80 ℃ 的冰箱内保存或直接播种在琼脂含量为 w = 1. 2% 的改良 Knop’s 平板培养基上。培养条件: 温度( 22 ±2) ℃,光照强度150 ~200 μmol·m- 2·s- 1,每天光照14 h,相对湿度85%以上。重复培养3 ~6 次。根据各阶段的发育特征,定期选取典型且稳定的活体材料制成临时水装片,在 Olympus-BX53 光学显微镜下观察,所有观测数据均采用随机测得的 20 个数的平均值。
2 结果与分析
2. 1 孢子及孢子萌发 孢子棕褐色较小,两侧对称,极面观椭圆形,赤道面观近圆形,周壁薄而透明,外壁上有扁圆形较整齐的疣状纹饰。单裂缝,裂缝长度约为孢子长度的1/2 ~3/4( 图版: 1) 。接种7 ~10 d,孢子吸水膨涨,孢子内部含有黄色球状油滴和叶绿体( 图版: 2) ,随着孢子萌发黄色油滴逐渐减小,这表明,脂肪类物质随孢子萌发而产生,但在萌发后不久就迅速转变为非脂肪类即碳水化合物或蛋白质。在单裂缝处伸出 1 条假根,有的假根内部含少量叶绿体且出现顶端膨大( 图版: 3) ,这种初生假根就出现顶端膨大的现象,是本种的1 个稳定特征,继续培养2 ~3 d,初生假根伸长,同时,基部也明显膨大( 图版:4) 。
2. 2 原丝体阶段 孢子培养约 17 d,原叶体原始母细胞经过连续横向分裂,发育成具有 3 个单列细胞的原丝体,此阶段的细胞分裂面都相互平行,且垂直于原丝体的长轴,表现出了颈卵器植物原丝体的共性。基部细胞伸出 2 条互成 90°的无色透明的假根,细胞中均匀分布着大量鲜绿色的叶绿体。中部细胞中的叶绿体较基部细胞中的叶绿体少,且分布不均匀,多沿细胞壁分布。上部细胞中的叶绿体集中分布在细胞的顶端,使顶端细胞呈深绿色( 图版: 6) 。原丝体不同部位细胞的光合能力是不同的,有些顶端细胞的生长方向发生弯曲( 图版: 5) ,具有明显的向光性。有少数原丝体在 2 ~3 个细胞时,前端会纵裂为双列丝状体,细胞呈圆柱状紧密排列,细胞质浓厚,富含叶绿体( 图版: 7) ,这是由丝状体向片状体过渡的表现。
2. 3 片状体阶段 孢子培养约 30 d,进入片状体时期。通常片状体的前端会偏向一侧,形成最初的生长点。生长点处的细胞体积小,且数量多,形状不规则,叶绿体数目多,而生长点周围细胞多呈圆柱形,体积较大,叶绿体数目较少,因而,生长点处为深绿色,且呈四周高中部凹状态。此时,假根数量增加到 7 条左右( 图版: 8) 。片状体阶段的生长速度明显快于丝状体阶段。
2. 4 原叶体阶段 孢子培养约 40 d,形成幼原叶体。这一时期细胞横向分裂次数多于纵向分裂次数,使幼原叶体形状不规则,两翼渐上延,生长点渐下陷且偏生。细胞形状呈多样性、大小不一,生长点处的细胞小且排列紧密,由生长点向外细胞体积逐渐增大。生长点附近的细胞边缘生出少量无色透明的乳突状毛状体。毛状体先端有少量叶绿体,顶端无腺状分泌物( 图版: 9) 。继续培养 5 ~6 d,由于生长点细胞的不断分裂调节,形成左右对称的狭心形原叶体,生长点下陷程度加深,上方的凹陷呈“V”型( 图版: 10) 。
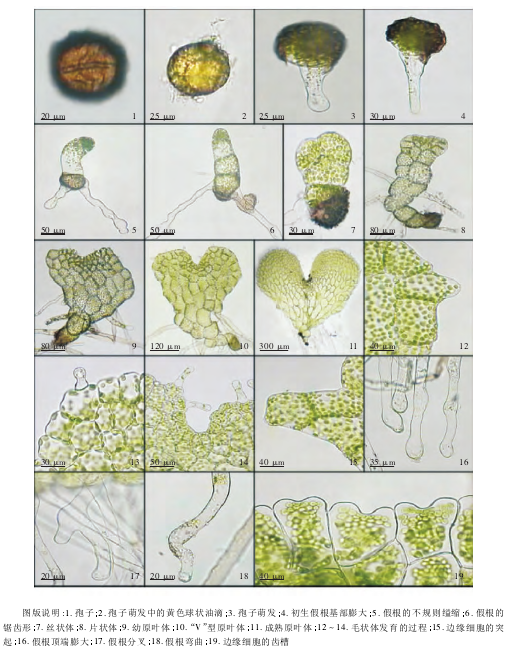
孢子培养 53 d 后,原叶体上出现精子器,标志着原叶体进入成熟阶段,其发育方式为槲蕨型( Drynaria-typa) 。成熟的原叶体呈阔心形且左右对称,基部稍尖,两侧微向外展,生长点位于原叶体中心 1 /3 处。基部及中部细胞较长,多呈六边形,两翼细胞大小均匀,近圆形,生长点处的细胞小而密集。在生长点及翼缘分布着较多的毛状体,原叶体背腹面密生几十条无色透明的假根( 图版: 11) 。
2. 4. 1 毛状体 多羽实蕨毛状体一般为单细胞棒状,最早由片状体细胞外壁向外突起形成透明帽状结构,还有叶绿体向其内部移动( 图版: 12) 。随着原叶体的发育,生长点附近陆续生出长棒状的毛状体且内有叶绿体和无色透明的分泌物( 图版: 14) 。当毛状体逐渐发育成熟,其内部的叶绿体几乎全部解体,在其顶部出现透明的帽状物质( 图版: 13) 。从毛状体的数量、细胞结构、分泌物的多少等方面能够看出,多羽实蕨具有较强的生理调节功能和环境适应能力。
2. 4. 2 边缘细胞 配子体边缘细胞存在多种形态: 外壁向外形成具有 3 细胞的突起,叶绿体均匀地分布在细胞内( 图版: 15) ; 外壁由中间或偏离中部向内凹陷形成齿槽,细胞多呈长方形,叶绿体集中分布在细胞内且胞壁上无叶绿体( 图版: 19) ; 两翼上缘细胞呈不规则六边形,翼缘细胞较小且有缺刻,使翼缘有明显的波状起伏( 图版: 11) 。这 3 种边缘细胞类型反映出不同的生理功能,其系统学意义也值得探讨。
2. 4. 3 假根 假根是由营养细胞向外突起形成的管状结构,直径约为 13 μm。幼时无色透明,含有少量叶绿体。随着配子体的发育,假根数量逐渐增多,形态也发生多种变化。初期长出的假根较短,且发生不规则缢缩( 图版: 5) ,或呈明显的锯齿形( 图版: 6) ,后期假根顶端或近顶端多膨大且顶端外有无色透明的分泌物( 图版: 16) ,偶有分叉现象,且分叉位置往往稍有膨大( 图版: 17) 。成熟配子体的假根有时出现弯曲( 图版: 18) 。假根的主要生理功能是固着和吸收,缢缩、锯齿形、顶端膨大、分叉、弯曲等形态都强化了其固着和吸收能力。
3、 讨 论
NAYAR指出,孢子的萌发类型、成熟原叶体形态、假根特征等有助于阐明类群之间的亲缘关系,是蕨类植物分类的重要特征。笔者观察到多羽实蕨( Bolbitis angustipinna ( Hayata) H. Ito) 的孢子为单裂缝,外壁有疣状纹饰,萌发方式为书带蕨型( Vittaria-typa) ,原丝体发育到 4 ~6 个细胞时,进入片状体阶段,发育类型为槲蕨型( Drynaria-typa) 。这既与鳞毛蕨科普遍存在的叉蕨型( Tectaria-typa) 发育方式不同,又与舌蕨属带状的原叶体不同,还与 VASHISHTA 所报道的藤蕨属的特征不同,本研究结果支持孙淑红等对实蕨( Bolbitis hainanensis) 孢子萌发方式和原叶体发育类型的研究结果,认为秦仁昌将实蕨属和刺蕨属独立为实蕨科更为合理。
NAYAR通过对一些实蕨种类的研究,发现大部分原叶体成熟时,两翼分布着发达的毛状体; 戴绍军等在报道鹿角蕨( Platycerium wallichii Hook) 配子体发育时指出,毛状体着生在原叶体边缘和表面是较进化的特征,毛状体顶端分泌物能免疫和抵抗不良环境,利于在自然界中生存,从而保证了物种的延续。笔者认为,多羽实蕨的毛状体也应该具有上述特征。
笔者不仅观察到了 MOMOSE描述的有关实蕨科配子体边缘具有的平坦、波状、内陷等类型的细胞形态,还新观察到了突起和齿槽等边缘细胞形态。MOMOSE曾提出,不同类群的蕨类植物,其原叶体的边缘细胞形态也是不相同的。因此,笔者新观察到的这种突起和齿槽的边缘细胞形态也应该具有相应的系统分类学参考价值。
就假根而言,多羽实蕨的假根具有 5 种不同的特征,分别是: 分叉、顶端膨大、弯曲、不规则缢缩和锯齿形。刘保东等对蹄盖蕨科( Athyriaceae) 、张开梅等对鳞毛蕨科( Dryopteridaceae) 、郭治友等对水龙骨科( Polypodiaceae) 等配子体发育研究过程中,也观察到假根有分叉或顶端膨大或弯曲等现象,可见这些特征在蕨类植物中还是较普遍的。刘保东等认为,假根形态特征的多样化不仅反映了固着能力的加强、吸收表面的增加,也反映出复杂的形态结构比简单的管状结构具有更进化的系统学意义。因此,本研究支持刘保东的观点,即多羽实蕨假根形态的多样性不仅在生态生理上具有较强的固着和吸收功能,在系统演化上也反映出了较高的进化程度。此外,对于笔者观察到假根的不规则缢缩和锯齿形,在现有蕨类配子体发育的文献中尚未见报道。此 2 种特征的功能与意义等都还有待进一步研究。
参考文献:
[1]吴兆洪,秦仁昌. 中国蕨类植物科属志[M]. 北京: 科学出版社,1991: 455 -459.
[2]吴兆洪. 中国植物志·第 6 卷( 第 1 分册) [M]. 北京: 科学出版社,1999: 104 -124.
[3]董仕勇,张宪春. 中国实蕨属的分类修订[J]. 植物分类学报,2005,43( 2) : 97 -115.

蹄盖蕨科(Athyriaceae)为中小型、少数为大型土生蕨类植物,约有20属500种,广布全世界热带至寒温带各地,尤以热带与亚热带山地为多。我国各属均产,约400种。蕨类植物孢子萌发和配子体发育的特性可以为分类学和种的研究提供有效的依据。近年来,对蹄...

本文重点对TET3介导甲基化调控在配子和胚胎发育中的作用进行阐述,以期促进TET3的功能性研究,完善在生殖发育中TET3介导的表观遗传学调控机制研究。...