分子生物学论文


真菌毒素( mycotoxin) 是由真菌产生的次级代谢产物,到目前为止已经发现超过 1000 多种真菌毒素,这些毒素仅由 350 多种真菌产生。真菌在陆地和海洋均广泛分布,在热带分布最广,因此污染也最重[1].目前,全球关于真菌毒素污染的报道日益增多,且受污染的也不局限于大米、玉米、小麦、大麦等主要粮食作物,还有药用植物、水果、坚果、牛奶等。毒性较强的真菌毒素有玉米赤霉烯酮( zearalenone,ZEN) 、伏马毒素( fumonisins,FB) 、黄曲霉毒素( aflatoxins,AFT) 、赭曲霉毒素 A( ochratoxin A,OTA) 、脱 氧 雪 腐 镰 刀 菌 烯 醇( deoxynivalenol,DON) 、镰刀菌素( fusarin) 、拟茎点霉毒素( phomopsins) 、桔霉素( citrinin) 等。不同的真菌毒素其致毒机制不同,表现为致癌毒性、细胞毒性、遗传毒性等多个方面。
由于真菌毒素对人类健康有巨大危害,因此国内外科研人员研发了多种检测真菌毒素的技术,主要有仪器确证法和利用免疫学原理筛选检测的方法。仪器法主要是高效液相色谱法[2]( HPLC) 、质谱分析[3]( MS) 、高效液相色谱-多级质谱法[4]等,这些方法显着的优势是能进行定性和定量分析且灵敏度高,但检测过程复杂、耗时、仪器设备十分昂贵、对实验室环境和实验员技能有严格的要求,从而限制了其用于大批量样品的筛选。利用免疫学原理进行大批量样品筛选检测的方法主要有酶联免疫吸附法[5]( ELISA) 和免疫层析试纸法[6],因其特异性高、灵敏度强、操作简单,甚至可以实现现场检测而发展迅猛。但上述方法均使生产人员和操作人员须长期接触真菌毒素标品,对其健康造成潜在危害,并存在二次污染的隐患。因此,近年来新兴的噬菌体随机肽库展示技术( phage display technology) 引起许多学者关注,并以此技术研究毒素的替代品及抗体参与免疫化学反应,建立了相关的无毒检测免疫学体系。
1 噬菌体展示技术简介
SMITH[7]在 1985 年首次证实外源 DNA 可以插入丝状噬菌体基因Ⅲ中,并与 pⅢ蛋白融合展示,从而创建了噬菌体展示技术。由于这一技术具有库容量大,结合特异性强等特点,促使科研工作者对噬菌体展示技术进行了深入的探索研究。
这些研究促进了无毒检测食品及饲料中真菌毒素体系的发展及昂贵真菌毒素抗体的替代。
1. 1 噬菌体展示系统的原理
噬菌体展示系统是一项选择技术,它将外源多肽或蛋白与噬菌体的一种衣壳蛋白融合表达,融合蛋白展示在病毒颗粒的表面,而编码这个融合子的 DNA 则位于该病毒粒子内。其原理是基于配体和受体反应的亲和选择性[8],即利用展示在噬菌体表面的多肽或重组抗体( 配体) 与目标抗体 或 抗 原 ( 受 体) 的 亲 和 能 力 进 行 淘 选( panning) ,淘选到与受体具有高亲和性的、空间构象与目标配体结构类似的模拟表位,从而可用这模拟表位取代相应的有毒抗原或不易获得的抗体。常用丝状噬菌体病毒包括 f1、fd 和 M13 三类,其基因组编码 11 种蛋白质,其中 5 种为结构蛋白,与噬菌体展示密切相关的是两种不同的结构蛋白 pⅧ和 pⅢ。pⅧ蛋白的 N 端( 1-5 氨基酸残基区域) 为可活动的、外露在噬菌体表面的肽段,是插入外源基因的最佳位置; pⅢ蛋白最外露的是穿膜区( N 端) ,因而亦是插入外源基因的理想位置,形成的融合蛋白表达在噬菌体颗粒的表面,不影响和干扰噬菌体的生活周期,同时也保持了外源基因蛋白的天然构象,并可被相应的受体所识别。利用固定于固相支持物的靶分子,采用适当的淘洗方法,洗去非特异结合的噬菌体,筛选出目的噬菌体,使得各种靶分子( 抗体、酶、细胞表面受体等) 的多肽通过体外选择程序得以快速鉴定。而其编码基因作为病毒基因组中的一部分可通过分泌型噬菌体的单链 DNA 测序推导出来。
该技术实现了基因型和表型的转换。
1. 2 噬菌体展示系统的分类
噬菌体展示系统分为: 丝状噬菌体展示系统、T7 噬菌体展示系统、T4 噬菌体与 λ 噬菌体展示系统; 其所展示内容包括随机肽段、天然肽段、cDNA、抗体、抗体片段等。根据展示内容所建立的文库又可分为随机肽库、抗体库、cDNA 文库等,其中用于筛选真菌毒素的模拟表位和不易获得抗体的常用文库分别是随机肽库和抗体库。
1. 2. 1 噬菌体随机肽库 随机肽库是将编码外源肽的 DNA 短序列插入到噬菌体编码外壳蛋白的基因组中,使各种组合的随机肽段与噬菌体外壳蛋白融合表达于噬菌体颗粒表面,被展示的肽可保持相对独立的空间构象和生物活性。而这特定的空间构象和生物活性能模拟小分子抗原,因此随机肽库经过 3 ~ 4 轮的筛选就能获得小分子的模拟表位。用于淘选的噬菌体展示肽库是基于完全随机的肽库序列,目前商业肽库主要有 7 肽库( Ph. D. -7) 、12 肽库( Ph. D. -12) 与环 7 肽库( Ph. D. -c7c) 等。随着研究的深入,为了获得亲和性更好的模拟表位,相关研究人员分析比较从随机肽库淘选到的阳性克隆噬菌体的氨基酸序列,获得其共有氨基酸序列,然后在共有序列两侧分布随机氨基酸残基构建二级肽库[9-10].从概率论和竞争关系的角度分析,二级肽库比随机肽库更易筛选到理想的模拟表位[11-12],是解决目前采用噬菌体文库筛选到的模拟表位与抗体的亲和性较低这个瓶颈的一个突破口。
1. 2. 2 噬菌体抗体库 传统的免疫检测抗原的抗体为多克隆抗体和单克隆抗体两种。多克隆抗体比较易于快速生产,但是它并不适合小分子检测,非特异性反应也较多; 而基于杂交瘤技术生产的单克隆抗体有很高的专一性,但单克隆抗体的生产费时、应用昂贵,且伤害动物。而基于噬菌体1抗体的优势又克服了其缺点[13],且为制备小分子抗体提供了有效手段。该技术能够将抗体功能片段有效地展示于噬菌体表面,利用靶抗原对噬菌体抗体库进行亲和筛选,从而获得针对该抗原的重组噬菌体抗体。
目前噬菌体展示技术可展示单链抗体( singlechain variant fragment,scFv) 及基于克隆重链抗体可变区构建的只由一个重链可变区组成的单域抗体 ( variable domain of heavy chin of heavy-chainantibody,VHH) 等。scFv 编码基因由重链可变区基因和轻链可变区基因通过人工合成一条寡核苷酸序列( linker) 相连融合表达在噬菌体载体的表面[14].重链抗体发现于羊驼、骆驼及护士鲨等动物体内,与普通抗体相比缺少了轻链,只保留了对抗原的结合能力的可变区; 基于其构建的 VHH保留了重链抗体的分子量小、物理稳定性高、易表达等 特 点,且 亲 和 力 高、不 易 聚 集 而 被 广 泛应用[15].
2 噬菌体展示技术在真菌毒素检测领域的研究与应用
噬菌体展示技术的不断发展,为真菌毒素检测领域的延伸和扩展带来新的方向。噬菌体展示技术主要应用于筛选真菌毒素的模拟表位和抗真菌毒素的重组抗体。
2. 1 噬菌体模拟表位肽在真菌毒素检测的研究与应用
近几年科研人员淘选各种真菌毒素模拟表位的工作情况见表 1.获得的模拟表位可用于ELISA、试纸条、快速试纸法和胶体金快速斑点结合免疫分析等方法检测真菌毒素。YUAN 等[16]淘选出 2 个较理想的模拟表位,合成了其中的一条模拟序列 SWGPFPF,用于检测小麦中 DON,其线性范围为 100 ~ 10 μg/ml; 同时,将合成多肽做了细胞毒理实验,结果表明对细胞没有明显的毒害,因此可利用模拟表位肽建立无毒免疫学检测体系。HE 等[17-18]将在 2011 年用 7 肽库筛选到的 1 个模拟表位与在 2014 年用 12 肽库淘选到的一个模拟表位进行了比较分析,发现在胶体金快速斑点结合免疫分析检测 ZEN 的临界值都是 50ng /ml,表明噬菌体展示肽的长度可能对小分子模拟表位的亲和性影响较小,决定性因素还是展示肽的序列及其构象。黄思敏等[19]
分别用 12 肽库和 7 肽库淘选桔霉素的模拟表位,最低检测限( LOD) 均为 10 ng/ml,进一步证明模拟表位亲和性的决定性因素主要是展示肽的序列及其构象,与其长短关系不大。WANG 等[20]对其实验室生产的单抗所淘选的 AFB1模拟表位与邓省亮等[21]使用中国协和医科大学的单抗所筛出的模拟表位进行了比较,结果显示前者的最低检测限和线性范围比后者提高了 40 倍。除了抗体本身的因素外,前者是使用 10% 甲醇进行竞争性洗脱淘选AFB1模拟表位,而后者使用酸洗脱淘选,表明洗脱试剂对能否淘选到理想的模拟表位也是一个关键影响因素。LIU 等[22]筛出伏马毒素 B1的模拟表位,将合成模拟表位的 7 个氨基酸偶联至 BSA并建立了 ELISA 方法检测伏马毒素 B1,结果显示无交叉反应,与商品化 ELISA 试剂盒和 HPLC 检测结果一致,表明模拟表位肽完全可以取代毒素标准品。YU 等[23]
获得了一个拟茎点霉毒素模拟表位并建立了相应的 ELISA 方法,与传统的ELISA 检测方法相比检测灵敏度提高了 100 倍。
同时,近年文献报道显示环肽库比非环肽库淘选出来的模拟表位半抑制率( IC50) 更低,分析其原因可能是成环的氨基酸序列更能模拟抗原的表位[24].2013 年,HE 等[26]构建二级肽库获得了 OTA的理想模拟表位。该课题组利用商业 12 肽库和环 7 肽库淘选获得阳性克隆噬菌体,分析了这些噬菌体的氨基酸序列,得出共有序列 FQLH,固定共有序列并在其两侧分布其他任意氨基酸残基构建了二级 12 肽库; 用这二级 12 肽库淘选出来的模拟表位建立 ELISA 方法检测 OTA,结果显示检测 OTA 的 LOD 达到 6 pg/ml,其检测灵敏度与用商业肽库直接淘选的模拟表位建立的 ELISA 方法的 LOD 150 pg/ml 相比极大提高。这些数据表明了二级肽库的优越性,因此在随机肽库的基础上构建二级肽库为今后获得检测灵敏度更高的真菌毒素模拟表位开辟了一条新途径。
以上研究显示,采用噬菌体展示技术淘选的真菌毒素模拟表位完全可以成为真菌毒素的替代品,用其建立的免疫学检测方法检测灵敏度可能更高且对生产和操作人员的伤害可降到最低。【1】

2. 2 噬菌体抗体库在真菌毒素检测的研究与应用
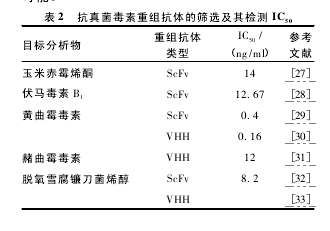
近几年利用噬菌体抗体库筛选的抗真菌毒素重组抗体并用其检测相应的真菌毒素的 IC50研究结果见表 2.YUAN 等[27]获得了抗 ZEN 的 scFv,用 scFv 建立了 ELISA 检测 ZEN,结果显示与单克隆抗体建立的 ELISA 方法检测结果一致,但 scFv与 ZEN 的类似物有更高的交叉反应,且不耐受甲醇溶液。ZOU 等[28]淘出的 scFv 与单克隆抗体相比在检测伏马毒素 B1的 IC50接近,无交叉反应,与 HPLC 对伏马毒素 B1的回收率实验结果相似,适合快速、大量、低成本的检测伏马毒素 B1.YANG 等[29]筛到了抗 AFB1的 scFv,表现为与AFB1类似物交叉反应低,优于传统的 ELISA 检测方法; 但与 WANG 等[30]淘选到 AFB1的 VHH相比,VHH 的 IC50更低且能耐受 20% 的甲醇,唯一不足的是与 AFB1类似物交叉反应率较高; 分析其原因可能与 VHH 缺少轻链更易与小分子抗原结合有关。HOUWELINGEN 等[31]筛到了一株与 OTA 类似物无明显的交叉反应、亦能耐受甲醇的 VHH.VHH 重组抗体比 ScFv 重组抗体在检测真菌毒素时具有更高的灵敏度和稳定性,究其原因可能是 VHH 重组抗体仅具有 3 个而不是 6 个互 补 性 决 定 区 环 ( complementaritydeterminingregion loop,CDR loop) 构象。重组抗体具有单克隆抗体相似的检测灵敏度,且拥有单克隆抗体不具备的价格低、易生产的优势,因此利用噬菌体展示技术筛选的重组抗体建立相应的检测方法来检测真菌毒素可以大大降低成本,为更全面地检测食物链中的各个环节的污染提供了可能。【2】
3 结语
真菌毒素对人类健康危害很大,所以加强检测食物链中的真菌毒素的污染是当务之急。然而,目前所报道的真菌毒素的检测成本高昂、同时对检测人员及生产人员都存在潜在的危害。噬菌体展示技术的出现及其在真菌毒素检测中应用,使得成本低廉且无毒的检测体系成为当今真菌毒素检测的新手段。但此方法涉及到展示的多肽在体外需要实现正确的折叠才能模拟真菌毒素,因此在实际应用中并不能完全替代真菌毒素标品且存在与抗体亲和力相对弱的情况。针对上述缺陷,目前已有科研工作者利用计算机软件进行抗体与模拟表位的分子对接分析,通过对模拟肽中的氨基酸残基进行替换来提高抗体与模拟肽的亲和力及提高肽的稳定性。相信科研工作者对噬菌体展示技术库的构建和淘选方法的不断探索及多学科的交叉,必然会克服不足,使利用无毒检测真菌毒素体系的检测方法灵敏度更高,特异性更好,成本进一步降低。

随着各种管家基因研究的深入, 在现有高通量测序和快速检测基础上, 结合痕量生物样本采集富集技术, 通过广谱覆盖的、不同特异性种属管家基因检测分析, 将促进各种复杂生物样品的基因分析、分类、鉴定和溯源的深入研究, 可能成为复杂样本分析研究热点之一。...

沙门氏菌是一种常见的食源性致病菌,种类繁多,目前已检测出2 500多种沙门氏菌血清型。沙门氏菌是一种革兰氏阴性的肠道杆菌,不但能显性或隐性感染动物,而且能通过污染的动物源性食品造成人的食物中毒,严重危害公共健康安全。...

非编码RNA在基因调控方面有着重要功能,可广泛参与基因调控的各个层面,在发育、癌症、神经功能紊乱等生物过程中都发挥重要作用[1]。在长非编码RNA中有一重要亚类,天然反义转录本,是从已知转录序列的互补链转录的转录物[2]。有学者提出一些分子模...
基因定点敲除技术通过对染色体上基因特异位点进行定点突变,使其功能丧失,进而达到研究其功能的目的。一些利用基因敲除技术创建的动物疾病模型对医学研究的发展做出了重要贡献。传统基因敲除技术于上世纪80年代末由Smithies等[1]创立,通过同源重组手段...
...

随着个体化医疗的不断发展,分子生物检测技术已广泛应用于科学研究和临床诊疗中。肿瘤分子生物检测能够为临床提供准确的病理诊断,有利于患者的精准治疗和个体化靶向治疗。...

丝状真菌与人们的生活密切相关,广泛应用于工业、农业和医药生产等领域,但这都离不开对丝状真菌的基础研究。然而在丝状真菌转化过程中,要从大量非转化子的背景中筛选目标转化子必须依赖遗传筛选标记。...