在离体脊髓横切片的细胞电生理研究中,除强直刺激背根可在腹角记录到场电位长时程增强(long-term potentiation, LTP) 现象外,背根传入和下行激活这两条通路也可诱发运动神经元 (moto-neuron, MN) 兴奋性突触后电位 (excitatory postsyn-aptic potential, EPSP) 的 LTP,而 MN 突触传递的LTP 现象可能是运动技能学习和记忆的生理机制之一。神经营养因子 -3 也可在 MN 诱发 EPSP 的类似LTP 现象,且 N- 甲基 -D- 门冬氨酸 (N-methyl-D-aspartate, NMDA) 受体在诱导而非维持中起重要作用。在 LTP 的分析中,一般用 EPSP 的幅度、曲线下面积、最大上升斜率以及上升相起始斜率等参数来定量描述突触效能的变化。尽管 LTP 所涉及的突触后受体机制已有大量研究,但递质与受体作用的受体动力学性质是否改变尚未见研究报道。鉴于分析药物 - 靶体关系的经时性动态过程,即可进行药物作用的受体动力学参数测定,本实验室已据此提出了突触反应的表观受体动力学分析方法,即通过分析突触传递过程中递质与突触后受体作用的历时性变化,以探讨递质与受体结合、解离的速率和受体的亲和力等受体动力学性质。
因此,本文应用新生大鼠 (8~14 d) 脊髓切片 MN细胞内记录技术,观察同侧腹外侧索 (ipsilateralventrolateral funiculus, iVLF) 电刺激诱发的 iVLF-EPSP 在其 LTP ( 即 iVLF-LTP) 过程中受体动力学性质的变化,为进一步研究脊髓 MN 等突触传递的可塑性机制提供新的方法。
1 材料与方法
1.1 动物及脊髓切片制备 动物实验方案经皖南医学院实验动物伦理委员会批准。取 8~14 d 龄新生Sprague Dawley (SD) 大鼠 15 只 ( 由南京江宁青龙山动物繁殖场提供 ),雌雄不拘。按报道方法,经乙醚麻醉后分离出脊髓,用振荡切片机 (Vi-bratome, Technical Products International Inc., USA)制备脊髓腰骶膨大部的横切片 (400~500 μm) 数片,置室温 (25 °C ± 3 °C) 下经 95% O2和 5% CO2混合气体饱和的人工脑脊液 (artificial cerebrospinal fluid,ACSF) 中备用。
1.2 细胞内记录及电刺激 用吸管将脊髓切片移入自制的全浸式记录浴槽,用上下两个丝网相夹固定,持续灌流经混合气体饱和的处于室温下的ACSF。 取 内 充 3 mol/L 乙 酸 钾、 尖 端 阻 抗 为80~140 MΩ 的微电极,在显微镜下对脊髓腹角进行穿刺。记录到的电信号由 Axoclamp-2B 微电极放大器 (Axon Instruments, USA) 放 大 后, 经 DigiData1200 转换接口 (Axon) 输入计算机,用 pClamp 7.0软件 (Axon) 进行采样、显示和贮存。细胞内电流注射由微机触发,经 DigiData 1200 联机接口到微电极放大器后,刺激电流由探头输出经微电极注入细胞内。iVLF及腹根电刺激,由三通道电刺激器(日本光电 ) 产生的方波脉冲,经隔离器以恒压模式至同芯双极刺激电极施加,分别用于激活下行纤维和腹根纤维。通过刺激电极在 iVLF 施加测试刺激 ( 单脉冲,波宽 0.1~0.3 ms,10 次 /min,10~100 V),在腹角MNs 记录 EPSP 反应,记录到稳定的 EPSPs 至少15 min 后,对 iVLF 施加强直刺激 (100 Hz,50 脉冲 / 串,波宽 0.4~1.0 ms,共 6 串,串间隔 10 s,电压强度同测试刺激 ),强直刺激后继续记录测试刺激 ( 参数同强直刺激前 ) 诱导的 iVLF-EPSPs 至少 30 min (10 次 /min)。
1.3 统计分析方法 对记录到的资料,用 Clamp-fit 10.1 软 件 (Axon Instruments/Molecular Devices,USA) 进行实验数据分析,其中 LTP 判定标准为 :EPSP 幅度增大到或超过基础值的 120%、增大的维持时间达 30 min 以上。测 EPSP 幅度时,取 1 min内 10 次记录的平均值,若 EPSP 上爆发动作电位(action potential, AP),则取首次出现的 AP 阈值水平。数据统计分析用 Origin 5.0 软件 (Microcal SoftwareInc., USA),结果以 mean ± SEM 表示,相关性采用直线相关分析,P< 0.05 时认为差异具有显着性意义。
1.4 受体动力学分析方法 按文献报道方法进行表观受体动力学分析,即依据突触反应大小依赖于受体激活数量,受体激活数量又依赖于递质的释放量,递质释放量则依赖于刺激强度大小的特性,将刺激强度的大小作为起作用的有效递质浓度高低的表观指标,将突触反应幅度大小作为递质结合突触后受体量的表观指标,进行表观受体动力学分析,此处的 “ 表观 ” 意为分别使用有效递质浓度、递质结合的突触后受体量的表观指标进行的受体动力学分析,以区别于直接测定递质浓度、递质结合受体数量的受体动力学分析。具体来说,就是将一定强度电刺激诱发的突触反应 EPSP 之上升相和下降相,与配体 - 受体的结合动力学过程和解离动力学过程相对应,以 EPSP 的幅度 (Veq,EPSP 峰值 ) 表征递质与受体的平衡结合浓度,以 EPSP 的不同时间膜电位值 (V) 表征递质结合的受体浓度,以电刺激强度 (S,以阈刺激强度 T 为单位 ) 表征有效递质浓度,再以 K1为表观结合速率常数、K2为表观解离速率常数,得 EPSP 上升相和下降相的直线化公式分别为 :ln[Veq/(Veq– V)] = (K1·S + K2)·t 和 ln(V/Veq) =K2·t,用 EPSP 的实测数据进行直线回归所得斜率分别为 K1S + K2和 K2。在求得 K2( 单位为 ms1)和 K1( 单位为 T1·ms1) 基础上,再进一步求得表观平衡解离常数 KT( 表征平衡解离常数 KD) :KT= K2/K1( 单位为 T),也称为表观半幅有效递质浓度 ( 表征 EC50)。KT代表受体的亲和力,KT值越小,说明递质与受体的亲和力越高。
2 结果
记录到静息电位负于 60 mV、AP 有超射的细胞后,通过置于腹根的同芯双极刺激电极对细胞进行逆行鉴定,记录到 “ 全或无 ” 特点的 AP,即可鉴定为 MN,再进行 iVLF-EPSP 的记录和下述实验分析。
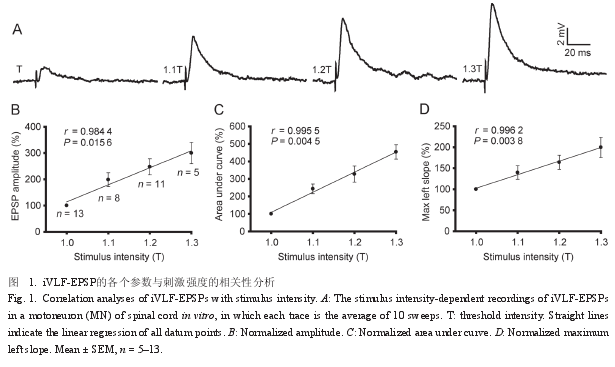
2.1 iVLF-EPSP的刺激强度依赖性通过置于 iVLF 的刺激电极进行不同强度 ( 单脉冲,波宽 0.1~0.3 ms,10 次 / 分,10~100 V) 的电刺激,刺激强度以阈刺激强度 (T) 的倍数表示,分析记录到的 iVLF-EPSP 的各个参数与刺激强度的相关性,在一个 MN 记录到的 iVLF-EPSP 呈刺激强度依赖性结果例示如图1A。直线相关分析结果显示,在 1.0T~1.3T 范围内,EPSP 的幅度、曲线下面积和最大上升斜率均与刺激强度呈正相关 (P < 0.05 或P < 0.01),且 1.3T 时 EPSP 幅度增大到 1.0T 时的300% 左右 ( 图 1B~D)。
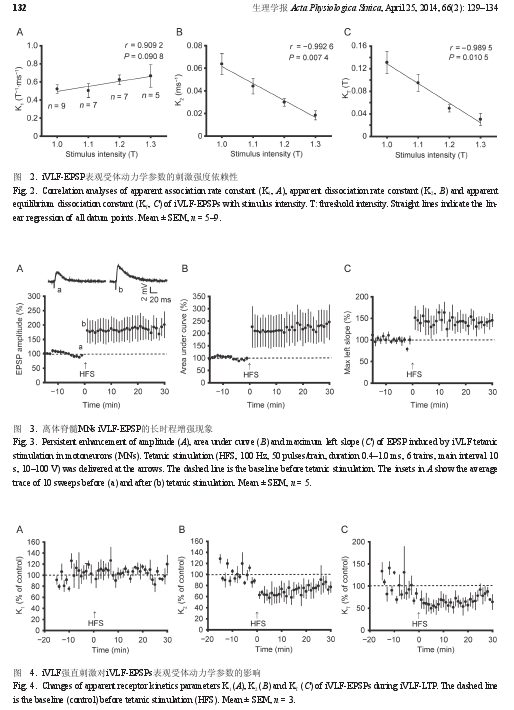
2.2 iVLF-EPSP表观受体动力学参数的刺激强度依赖性。对不同强度电刺激诱发的 iVLF-EPSPs 进行表观受体动力学分析,显示刺激强度在 1.0T~1.3T 时,表观受体动力学参数 K2和 KT与刺激强度均呈负相关 (P < 0.05,图 2)。
2.3 iVLF-EPSP的LTP现象通过置于 iVLF 的同芯双极刺激电极,给予iVLF 测试刺激 ( 单脉冲,波宽 0.1~0.3 ms,10 次 / 分,10~100 V) 记录到稳定的 EPSP 15 min 后再给予强直刺激 (100 Hz,50 脉冲 / 串,波宽 0.4~1.0 ms,共6 串, 串 间 隔 10 s,10~100 V), 在 测 试 的 11 个MNs 中,有 5 个 MNs 的 EPSP 幅度在强直刺激后增大到基础值的 120% 以上,且维持至少 30 min,可以被判定诱发了 LTP (iVLF-LTP)( 图 3A),同时EPSP 的曲线下面积 ( 图 3B)、最大上升斜率 ( 图3C) 均增大到基础值的 120% 以上。
2.4 iVLF-LTP过程中iVLF-EPSPs受体动力学的变化。选择诱发出 iVLF-LTP 的 5 个 MNs 进行表观受体动力学分析,结果表明在 iVLF-LTP 过程中,iVLF-EPSPs 的 K1未呈现规律性变化,而有 3 个 MNs 中K2和 KT明显减小,在强直刺激后 10 min 内减小到基础值的 80% 以下,后逐渐有所恢复 ( 图 4B、C),但在剩余 2 个 MNs 未发生明显变化。
3 讨论
本研究观察到的现象是 iVLF-EPSP 的表观受体动力学参数在 1.0T~1.3T 范围内,具有刺激强度依赖性,如表观解离速率常数 K2和表观平衡解离常数 KT均与刺激强度呈高度负相关。同时,iVLF-EPSP 的幅度在 1.3T 刺激强度时可增大到 1.0T 刺激强度时的 300% 左右,这在 LTP 分析中可能是比较强的突触效能增强作用。这些观察结果提示了 iVLF-EPSP 表观受体动力学参数分析,可能在 iVLF-LTP的受体动力学分析中有一定的参考价值。
就本研究的 iVLF-EPSP 而言,介导突触反应的离子通道型谷氨酸受体可能有 NMDA 受体、AMPA受体和 KA 受体。已有研究表明,KA 受体在 iVLF-EPSP 中的作用可能很小,因为 AMPA 受体拮抗剂和 NMDA 受体拮抗剂能够完全阻断刺激诱发的反应,而且本实验室前期研究结果提示,生理情况下 VLF-EPSP 可能主要由 non-NMDA 受体介导。
EPSP 具有刺激强度依赖性,在本实验中,EPSP 的幅度等参数都与刺激强度呈高度正相关。AMPA 受体的特点是快速脱敏和低亲和力;而 NMDA受体介导的电流不仅上升慢,衰减也特别慢,可能与 NMDA 受体的功能亚基 NR2 与谷氨酸的亲和力较高有关。本结果显示,K2值随着刺激强度的增大而减小,提示递质与受体分离减慢,且 KT值与刺激强度亦呈负相关,递质与受体作用的亲和力随刺激强度增大而增强。在 iVLF-EPSP,这种亲和力的增强可能与 NMDA 受体的激活有关 ,但其它可能的受体机制是否参与仍需进一步研究。
给予 iVLF 强直刺激,5 个 MNs 的 iVLF-EPSP幅度表现为显着增大到或超过基础值的 120%,并且至少维持 30 min,根据在其它脑区的 LTP 判断标准,可以判定发生 iVLF-LTP。在脊髓背角,NMDA 受体激活是触发中枢敏化的必要条件,高频刺激使突触后膜去极化,封闭 NMDA 受体的 Mg2+被移除,NMDA 受体激活,Ca2+内流激活钙敏感的胞内信号级联导致 NMDA 受体和 AMPA 受体的磷酸化,产生 LTP。使用 APV 阻断 NMDA 受体可以阻断 LTP 的诱导,而在 LTP 的维持中,突触后 AMPA 受体量明显上调。本研究选择诱发出iVLF-LTP 的 5 个 MNs 进行 EPSP 的表观受体动力学分析,其中 3 个 MNs 的 iVLF-EPSP 的表观平衡解离常数 KT值在强直刺激后 10 min 内明显减小,后逐渐部分恢复,提示在部分 iVLF-LTP 有递质与突触后受体的亲和力增大现象,这首先要考虑强直刺激后有大量高亲和力的 NMDA 受体被激活所致的可能性,其后 LTP 维持中 AMPA 受体数量的上调则可能是导致亲和力有所恢复的原因之一。另外,不同类型 NR2 亚基的 NMDA 受体,其失活时间不同,大小顺序为 NR2A < 2C = 2B << 2D。已有研究显示 NMDA-2B 受体拮抗剂 Ro 25-6981 能够明显减弱强直刺激后脊髓 C 纤维 LTP 的幅度,表明含NR2B 亚基的 NMDA 受体的激活与脊髓 LTP 的维持有关。更有报道,不仅在 LTP 的产生机制中,突触外 NR2B 的侧向位移至突触部位贡献其中,而且突触外的含 NR2D 的 NMDA 受体在 LTPNMDA中被补充到突触部位。尽管以上研究结果均提示iVLF-LTP 过程中突触后受体亲和力的提高可能与NMDA 受体的激活、NR2B、2D 亚基的参与有关,但局部神经元网络、LTP 产生机制的复杂性,以及发育过程中 NMDA 亚基表达有一个从 NR2B 到NR2A 的转换过程等,均强烈提示 iVLF-LTP 过程中突触后受体亲和力是否升高可能涉及更多因素,如其它神经递质的参与、受体的表达与修饰等,这些均值得进一步研究。
综上所述,本研究的受体动力学分析显示,在iVLF-LTP 过程中 iVLF-EPSP 表观受体动力学参数K2和 KT出现明显减小,可能表明突触后受体亲和力的升高,不仅提示了所涉及的受体机制以及其它相关机制需进一步的研究,也为相关突触传递的可塑性研究提供了一种可以参考的分析方法。
参考文献
1Pockett S, Figurov A. Long-term potentiation and depressionin the ventral horn of rat spinal cord in vitro. Neuroreport1993; 4(1): 97–99.
2 Su YY (苏彦艳), Jiang X, Wang MY. Long-term potentia-tion of synaptic transmission in motoneurons induced by te-tanic stimulation of ventrolateral funiculus of neonatal ratspinal cord in vitro. J Wannan Med Univ (皖南医学院学报)2007; 26(3): 161–164 (Chinese, English abstract).
3 Jiang X (江潇), Wang MY. Long-term potentiation of synap-tic transmission in motoneurons induced by tetanic stimula-tion in contralateral ventrolateral funiculus of neonatal ratspinal cord in vitro. J Wannan Med Univ (皖南医学院学报)2008; 27(6): 391–394, 398 (Chinese, English abstract).
4Arvanov VL, Seebach BS, Mendell LM. NT-3 evokes anLTP-like facilitation of AMPA/kainate receptor-mediatedsynaptic transmission in the neonatal rat spinal cord. J Neu-rophysiol 2000; 84(2): 752–758.
5 Jiang J (蒋娟), Sui N, Wang MY. Intracellular recordingsand multi-parameter analysis of long-term potentiation ofsynaptic responses in chick brain slices. Acta Physiol Sin (生理学报) 2009; 61(6): 577–584 (Chinese, English abstract).
6Yang SN, Tang YG, Zucker RS. Selective induction of LTPand LTD by postsynaptic [Ca2+]ielevation. J Neurophysiol1999; 81(2): 781–787.
7Kapur A, Yeckel MF, Gray R, Johnston D. L-Type calciumchannels are required for one form of hippocampal mossy fiberLTP. J Neurophysiol 1998; 79(4): 2181–2190.
8 Yang SL (杨守礼), Huang SL, Wen YY, Zhang SF, WangSZ. Kinetic method for receptor assay. Acta Acad Med Sin(中国医学科学院学报) 1984; 6(2): 153–155 (Chinese, Eng-lish abstract).
9 Luo H (罗浩), Zhang Y, Wang MY. Apparent receptor kinet-ics analyses of synaptic responses in spinal cord motoneu-rons in vitro. J Wannan Med Univ (皖南医学院学报) 2013;32(3): 173–177 (Chinese, English abstract).
10 Wang MY (汪萌芽). Responses of motoneurons to ventro-lateral funiculus stimulation in neonate rat spinal cord slices.Acta Physiol Sin (生理学报) 1994; 46(2): 148–153 (Chi-nese, English abstract).
11Dingledine R, Borges K, Bowie D, Traynelis SF. The gluta-mate receptor ion channels. Pharmacol Rev 1999; 51(1): 7–61.
12 Zheng C (郑超), Wang MY. Effects of etomidate on de-scending activation of motoneurons in neonatal rat spinalcord in vitro. Acta Physiol Sin (生理学报), 2012; 64(2):155–162 (Chinese, English abstract).
13Mayer ML, Armstrong N. Structure and function of gluta-mate receptor ion channels. Annu Rev Physiol 2004; 66:161–181.
14Tsvyetlynska NA, Hill RH, Grillner S. Role of AMPA recep-tor desensitization and the side effects of a DMSO vehicleon reticulospinal EPSPs and locomotor activity. J Neuro-physiol 2005; 94(6): 3951–3960.
15Dodt HU, Frick A, Kampe K, Zieglg nsberger W. NMDAand AMPA receptors on neocortical neurons are differentiallydistributed. Eur J Neurosci 1998; 10(11): 3351–3357.
16McAllister AK, Stevens CF. Nonsaturation of AMPA andNMDA receptors at hippocampal synapses. Proc Natl AcadSci U S A 2000; 97(11): 6173–6178.
17Anson LC, Schoepfer R, Colquhoun D, Wyllie DJ. Single-channel analysis of an NMDA receptor possessing a muta-tion in the region of the glutamate binding site. J Physiol2000; 527(Pt 2): 225–237.
18Lee CM, Stoelzel C, Chistiakova M, Volgushev M. Het-erosynaptic plasticity induced by intracellular tetanization inlayer 2/3 pyramidal neurons in rat auditory cortex. J Physiol2012; 590(Pt 10): 2253–2271.
19Mameli M, Bellone C, Brown MT, Lüscher C. Cocaine in-verts rules for synaptic plasticity of glutamate transmissionin the ventral tegmental area. Nat Neurosci 2011; 14(4):414–416.
20Woolf CJ, Salter MW. Neuronal plasticity: increasing thegain in pain. Science 2000; 288(5472): 1765–1769.
21Ikeda H, Heinke B, Ruscheweyh R, Sandkühler J. Synapticplasticity in spinal lamina I projection neurons that mediatehyperalgesia. Science 2003; 299(5610): 1237–1240.
22Ji RR, Kohno T, Moore KA, Woolf CJ. Central sensitizationand LTP: do pain and memory share similar mechanisms?Trends Neurosci 2003; 26(12): 696–705.
23Monyer H, Burnashev N, Laurie DJ, Sakmann B, SeeburgPH. Developmental and regional expression in the rat brainand functional properties of four NMDA receptors. Neuron1994; 12(3): 529–540.
24Pedersen LM, Gjerstad J. Spinal cord long-term potentiationis attenuated by the NMDA-2B receptor antagonist Ro256981. Acta Physiol (Oxf) 2008; 192(3): 421–427.
25Zhao J, Peng Y, Xu Z, Chen RQ, Gu QH, Chen Z, Lu W.Synaptic metaplasticity through NMDA receptor lateral dif-fusion. J Neurosci 2008; 28(12): 3060–3070.
26Harney SC, Jane DE, Anwyl R. Extrasynaptic NR2D-containing NMDARs are recruited to the synapse during LTPof NMDAR-EPSCs. J Neurosci 2008; 28(45): 11685–11694.
27Xing GG, Wang R, Yang B, Zhang D. Postnatal switching ofNMDA receptor subunits from NR2B to NR2A in rat facialmotor neurons. Eur J Neurosci 2006; 24(11): 2987–2992.