��������ѧ����


����Ghrelin ���������شٷ���������( growth hor-mone secretagogue receptor��GHS-R) ����Դ�����壬��Ҫ��θ�Ĥ��֯�е�����ϸ�����ڡ�Ghrelin ������ѧ����ʮ�ֹ㷺���ɴٽ��������ط���; ������ʳ��������л; �̼�θ������θ�����; ������Ѫ�ܹ��ܵȡ��½��о����֣�ghrelin ���������˶���ϵ���У����ܺ�������� ghrelin �ɼ���θ�ſգ����������裬�ٽ�θ��ʮ��ָ���������ڸ��ϼ�����Gong ���õ���ӡ�������;ۺ�ø����Ӧ( polymerase chain reaction��PCR) �������� GHS-R�ڴ�����������б���; Gnanapavan ���� PCR ����֤ʵ��ghrelin �� GHS-R ���˵�θ��С�����᳦�����١����ҡ����ࡢ���塢�ѳ���Ѫ�ܵȴ����б���;Dass�� Xu ������ӫ��������֯��ѧ�������ˡ�����������θ�ͽ᳦Ҳ������ ghrelin �������塣Ȼ�������� ghrelin �� GHS-R ���������֯ϸ��ѧ�ֲ����������ıȽ���δ�����������⣬������������֯�Ƚ������ж������ŵľ�ȷ���ߣ�PCR������ӡ������ֻ�ܷ��������IJ�������( �纣����������) �����ܷ�������а�����С���ź��Ǻ��ţ������б������Ƚϡ���ˣ����߲���������֯��ѧ��ͼ����������۲�������� ghrelin ��GHS-R �ķֲ����ص�����Ƚ϶�����������������������ı��Ϊ����������� ghrelin �Ĵ�θ�����������ṩ�������ݡ�
����1 �����뷽��
����1�� 1 ʵ�鶯�� �������� Sprague-Dawley ���� 8ֻ�����۲��У������� 220 ��240 g����������ͨ��ѧҽѧԺʵ�鶯�������ṩ��
����1�� 2 ��Ҫ�Լ������� �ÿ��� GHS-R ���幺������ Santa Cruz ��˾���ÿ��� ghrelin ���幺���������������Ʒ����˾����ù����-������-��������ø������( strept-avidin-biotin complex��SABC) ����������֯��ѧ�Լ��й����人��ʿ�¹�˾��������������( diaminobenzidine��DAB) ��ɫ�Լ��й��Ա�����ɽ���\������˾��Q550CW ��ͼ��ɼ������ϵͳ���Ե¹� Leica ��˾��
����1�� 3 ȡ����걾�Ʊ� ʵ������ú� 100 g·L�� 1��ˮ����ȩ400 mg·kg�� 1��ǻע���������������Ҵ���������������������ˮ���ٳ�Ѫ�ܣ��� 40 g·L�� 1�Ķ�ۼ�ȩ����Һ������������ע��ȡ�����Ժ�С�ԣ��гɺ�4 mm С�飬40 g·L�� 1�Ķ�ۼ�ȩ����Һ�̶�8 h���ú��������� 20% �� 30% ��������Һ�ݶ���ˮ��Һ���ٶ�����֯����������������Ƭ����Ƭ����Ƭ��������Ƭ����15 μm��
����1�� 4 ������֯��ѧȾɫ �� SABC ������ ghrelin�� GHS-R ��������֯��ѧȾɫ�����岽������: �䶳��Ƭ�������λ���Һ( phosphate buffered saline��PBS) ���� 5 min�����������Ỻ��Һ( pH 6�� 0) ���������������£�PBS ��ϴ3 �Σ�ÿ�� 5 min; �������3% �Ĺ���������Һ( hydrogen peroxide��H2O2) ����30 min��PBS ��ϴ 3 �Σ�ÿ�� 5 min; ������� 0�� 5% �ľ��Ҷ�������������( triton X-100) ���� 20 min��PBS��ϴ3 �Σ�ÿ��5 min��������� 10%����ɽ��Ѫ����( 37 �棬30 min) ��˦ȥ����Һ�壬��Ƭ�ֱ�μ��ÿ��� ghrelin ����( 1: 2 500) ���ÿ��� GHS-R ����( 1:100) ��4 ���ҹ��PBS ��ϴ3 �Σ�ÿ��5 min���μ����ÿ���( 37 �棬40 min) ��PBS �� ϴ 3 �Σ�ÿ ��10 min���μ������ػ���������������ø������( avidin-biotinylated horseradish peroxidase complex��ABC) ( 37 �� ��20 min) ��PBS ��ϴ 3 �Σ�ÿ�� 10 min��DAB ��ɫ������������Ƭ����ͼ�����ϵͳ�۲� ghre-lin �� GHS-R ���Ա���� PBS ����һ�������Զ��գ����ಽ�費�䡣
������ʵ��ѡȡ������������صĺ��Ž����ص�۲죬�������� ghrelin �� GHS-R ���Է�Ӧϸ���ڸ����Ų�ͬ�����ı����������ϸ������: ÿֻ������� 5 ����Ƭ��ÿ����Ƭ������ϸ�����ܼ������ѡȡ 5 ����Ұ������ȡƽ��ֵ��Ϊ����Ƭ������ϸ��������ÿֻ����ͬһ������ ghrelin �� GHS-R����ϸ������ʱ����ѡ������ͬƽ�档
����1�� 5 ͳ��ѧ���� Ӧ�� SPSS 13�� 0 ͳ�������������ݷ��������߷�Ӧ����ϸ�����Ծ��� ± ����( x�� ± s) ��ʾ�������Ƚϲ��÷��������P ��0. 05 Ϊ������ͳ��ѧ���塣
����2 ���
����2�� 1 Ghrelin �� GHS-R ��������ϸ����Ⱦɫ�����ڴ���������ϵͳ��ghrelin �� GHS-R ��������ϸ���㷺�ֲ��ڶ�����������ߵ����Ա������ƣ�����ϸ��Ϊϸ��Ĥ��( ��) ϸ�������ػ�ɫ�������ţ�ϸ�������ػ�ɫ�������š����ߵ�����ǿ���Է�Ӧ�ڴ���Ƥ�ʡ������ԡ��������ܺˡ�����( hippocamp��HIP) �����ʺˡ�С�Լ��ԸɵȲ�λ���ɼ� ( ͼ 1��ͼ 2) ����ͼ1-2�ԡ�
����
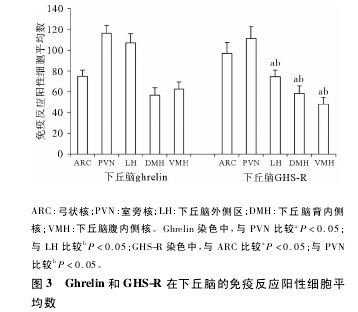
����2�� 2 Ghrelin �� GHS-R ��������ϸ������������������������ı��� �������������Ժ�( periven-tricular nucleus��PVN ) ���� ״ �� ( arcuate nucleus��ARC) ����ǰ��( preoptic nucleus��PON) �����Ϻ�( su-praoptic nucleus��SON) �������������( lateral hypo-thalamic area��LH) �������Ը��ڲ�� ( ventromedialhypothalamic nucleus��VMH) �������Ա��ڲ��( dor-somedial hypothalamic nucleus��DMH) �������Ա����( dorsal hypothalamic nucleus��DO) �ȴ����ɼ� ghre-lin �� GHS-R ���Է�Ӧϸ��������ϸ����ϸ��ĤΪ�ػ�ɫ�������ţ�ϸ�������ػ�ɫ�������š����У�ghrelin �� PVN �� LH ���Է�Ӧ��ǿ�����߷�Ӧ����ϸ�����ֱ�Ϊ116�� 31 ±7�� 76 ��107�� 02 ±8�� 80�����Ը��� ARC( 75�� 12 ± 5�� 59) ��DMH( 56�� 84 ± 7�� 07) ��VMH( 62�� 54 ± 6�� 93 ) ��������ͳ��ѧ���� ( P ��0. 05) ( ͼ 1��ͼ 3) ; GHS-R �� PVN �� ARC ���Է�Ӧǿ�����߷�Ӧ����ϸ�����ֱ�Ϊ 111�� 06 ± 11�� 80 ��97�� 06 ± 10�� 37�����Ը��� LH( 74�� 70 ± 6�� 04) ��DMH( 58�� 65 ±7�� 28) �� VMH( 48�� 24 ± 6�� 58) ��������ͳ��ѧ����( P �� 0�� 05) ; ����ϸ��Ⱦɫ���Ŀ�࣬ϸ�����Բ�λ���Բ�Σ�����ϸ���дֶ�ͻ�������������ϸ����Ŀ���٣��ֲ�ϡ�裬Ⱦɫ�ʵ���ɫ( ͼ 2��ͼ 3) ����ͼ3��
����
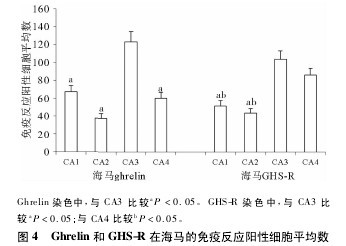
����Ghrelin �� GHS-R �� HIP �� CA1��CA2��CA3��CA4 ���Ŀ�������зֲ���ϸ����������Բ�Σ�ϸ��Ĥ������ɫ��ϸ�����С���ֲ��ܼ������У�ghrelin�� CA3 �����Է�Ӧ��ǿ�����߷�Ӧ����ϸ����Ϊ123�� 20 ± 11�� 17�����Ը��� CA1( 67�� 46 ± 6�� 93) ��CA2( 37�� 91 ±5�� 21) ��CA4( 59�� 96 ±6�� 73) ����������ͳ��ѧ����( P �� 0�� 05) ( ͼ 1��ͼ 4) ; GHS-R �� CA3 ��CA4 ������ǿ���Է�Ӧ�����߷�Ӧ����ϸ�����ֱ�Ϊ 103�� 79 ± 9�� 23 �� 86�� 48 ± 7�� 25�����Ը��� CA1( 51�� 16 ±6�� 56) ��CA2( 43�� 39 ±5�� 11) ����������ͳ��ѧ����( P ��0�� 05) ����ϸ����ɼ����ߵ�ǿ���Է�Ӧ( ͼ 2��ͼ 4) ��ϸ��Ĥ������ɫ��ϸ����ϴ�Բ�Ρ���Բ�λ��Σ�ͻ��ֶ̣���ɫ���ԣ�ϸ���ʵ�Ⱦ�������ڷ��Ӳ��δ����ɫ����ͼ4��
 ����
����
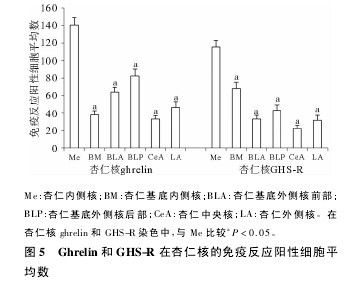
���������ʺ�����ghrelin �� GHS-R ����ϸ��ϸ��Ĥ��( ��) ϸ����Ϊ��ɫ�������ţ�ϸ���˲���ɫ�����߾��������ڲ��( medial amygdaloid nucleus��Me)��ǿ �� �� �� Ӧ���� �� �� Ӧ �� �� ϸ �� �� �� �� Ϊ140�� 22 ± 9�� 07 �� 115�� 66 ± 7�� 35�����Ը������ʻ����ڲ�� ( basomedial nucleus of the amygdale��BM)( 67�� 84 ±7�� 36) �����ʻ�������ǰ��( anterior divi-sion of the basolateral nucleus of the amygdale��BLA)( 33�� 46 ±4�� 24) �����ʻ������˺�( posterior di-vision of the basolateral nucleus of the amygdale��BLP)( 43�� 06 ±6�� 24) �����������( central amygdaloid nu-cleus��CeA) ( 22�� 58 ± 3�� 60) ����������( lateral a-mygdaloid nucleus��LA) ( 31�� 95 ± 5�� 77) ��������ͳ��ѧ����( P ��0�� 05) ( ͼ 1��ͼ 2��ͼ 5) ��ϸ�����������Բ�Σ�ϸ��ĤΪ����ɫ������ϸ���ж̴�ͻ��
����Ghrelin �� BLA �� BLP Ҳ�н϶�����ϸ����ϸ����ϴ��ж̴�ͻ����̬��Բ�ξӶ࣬ϸ��ĤȾɫ�ʵ���ɫ��GHS-R �� BM �н϶�����ϸ����ϸ�����С����Բ�λ���Բ�Σ�Ⱦɫ���ػ�ɫ���� CeA ��LA�����ߵ�����ϸ����Ŀ���٣�ϸ�����С��Ⱦɫ�ʵ���ɫ��
����2�� 3 Ghrelin �� GHS-R ��������ϸ��������������λ�ı��� �ڴ���Ƥ������������� ghrelin ��GHS-R ���Է�Ӧϸ���������ڷ��Ӳ㡢������㡢����ϸ�������Է�Ӧ��������Ⱦɫ�ʵ���ɫ������ϸ�������������������С����̬��Բ�κ���Բ�ξӶ࣬���ɼ�����ϸ�������ڿ����������ϸ���㣬ghrelin���Է�Ӧ��ǿ������ϸ�����ػ�ɫ��������������̬��Բ�λ���Բ�Σ���ͻ��Ⱦɫ��ϸ���˲���ɫ��GHS-R���Ա���������ϸ����϶࣬�ڿ����������������ϸ���㣬GHS-R ��ghrelin ���Է�Ӧ����ǿ( ͼ1��ͼ2) ��ϸ�����ػ�ɫ��ͻ��Ⱦɫ���ԣ�ϸ���˲���ɫ����̬��Ϊ�Ρ���ͼ5��
 ����
����
������С���ѿ�Ұϸ����ghrelin �� GHS-R ����ǿ���Է�Ӧ( ͼ 1��ͼ 2) ��ϸ����������Σ�ϸ��Ĥ��ϸ����������ɫ��ͻ����������ɫ���������Է�Ӧ��С�Եķ��Ӳ�Ϳ����������ϸ��С��Բ��ϸ��Ĥ��ϸ���ʳ��ػ�ɫ�����⣬��ǰͥ���Ϻˡ����ˡ��з�ˡ�Сϸ����״�˺����˵ȴ����ɼ�ghrelin �� GHS-R ����ϸ����
���������裬ghrelin �� GHS-R ����ǿ���Է�Ӧ( ͼ 1��ͼ 2) ��ϸ��Ĥ��ϸ���ʳ�����ɫ��ϸ����ʶ����ԣ�������ͻ���ڹ����ˡ��������ˡ�������״�˼�������ȴ�Ҳ�ɼ� ghrelin �� GHS-R ����ϸ����
����3 ����
�����½��о����֣�ghrelin ���������˶���ϵ���У��ɴٽ���ʳ��θ�ſգ�����θ����ڣ�����С��ת�ˣ���Ч���������衣�������о����ע��ghrelin ����֯�ֲ��������о����֣�ghrelin ��θ������������ϵͳ���зֲ�����Ҫ��θ�Ĥ�ڷ���ϸ��( ��ҪΪ X/A ��ϸ��) ���ڣ���С�������ࡢ�����ԡ��ٴ��塢���١���״�١����ࡢ�ѳ���̥�̵ȴ�Ҳ���������ڣ���ͨ���ڷ��ڻ��Է�����ʽ���������壬�ھֲ��������á����������� GHS-R �ֲ����о�������֡������о����֣�GHS-R ��Ҫ�ֲ��ڴ��塢�����Ժ�θ�������ࡢ�Ρ����١�����֬����֯����Ҳ�зֲ�����������ڷ�����������л��Ŀǰ�����ڶ������������������֯ѧ��λ��С���ź��Ǻ��ŵı���ˮƽ�Ƚ���δ�����������߾ʹ˽����˽�һ���о������ص�����˶���������������������������ı���ˮƽ�Ƚϡ�
�����о������ʾ��GHS-R �ڴ�������㷺���ڣ�����ϸ��ϸ���˲���ɫ��ϸ��Ĥ�Ż�ɫ���ػ�ɫ��ϸ������������ɫ����Ҫ�ֲ��ڴ���Ƥ�ʡ������ԡ��������ܺˡ�HIP�����ʺˡ�С�Լ��ԸɵȲ�λ�������о��õ���ӡ������ PCR ����֤ʵ GHS-R ����������ܾ��б����ӫ��������֯��ѧ�������ˡ�����������θ�ͽ᳦����Ҳ���� ghrelin ��GHS-R�����о������������������ڴ˻����Ͻ�һ���۲��˶��������������������������( �������ż��Ǻ���) �ı��
����������λ�ڼ��Եĸ��ಿ�����ɵ� 3 �����²��ĵױںͲ�ڡ����������ڷ��ڡ�ˮ����ʴ�л�����µ��ڵȹ��ܵĵ�����������Ҳ�ǵ�����ʳ���������˶�����Ҫ���ࡣ�����Դ�ǰ����Ϊ 3 ����: ���ϲ�λ���ӽ����Ϸ����� SON �� PVN���; ��ڲ�λ��©���ĺ�; ��ͷ��λ����ͷ�塣
���������������������и��Ӷ��㷺����ά��ϵ���������������Դ���Ƥ�ʡ���Եϵͳ���ԸɵĴ�����ά����������������ά�������Ƥ�ʡ���Եϵͳ�����ԡ����ź����衣��������ͨ�� 3 ��;���Ի�����е���: ( 1) �������Ժ˷��������д����������Ըɺͼ����ֲ���������࣬��ͨ��ֲ������������; ( 2) �����Ե� SON �� PVN ��������ά����������-�������������壬���˷��ڵļ�ѹ��( ��������) �ʹ߲����ش����������������棬���������ͷ���ѪҺѭ��; ( 3) �����Կɷ��ڶ��ֶ��������ض��ٴ���ķ����������Դ̼����û��������ã���Ϊ�ͷż��ػ������ͷż��ء�
���������о����֣�ghrelin �Ĵ�θ������������������������أ������� ARC �ɲ��������ٶ������á������о�֤ʵ���������ڴ��� ghrelin ��GHS-R����Ҫ�ֲ��������ԣ�����δ�о�������������Ҫ�Ǻ��ŵı�����о������ʾ��ghrelin�� GHS-R ��������ϸ���������� PVN��ARC��LH��VMH��DMH �ȴ����зֲ���Ghrelin ��������ϸ���� LH �� PVN �Ӷ࣬GHS-R ��������ϸ���� PVN ��ARC �Ӷࡣ����ʾ���������˶��ĵ����У������Է�������Ҫ���ã�ghrelin ����ͨ�� PVN��ARC �� LH����������ѧ���á�
����HIP ��������֪������ѧϰ����С������йأ�Ҳ�ǵ�����������Ҫ���࣬���������ԡ����ʺˡ������������ż��临�ӵ����ϵ���ڵ��ڲ��鶯��Ľ�ʳ��Ϊ�����м�θ���˶��з�������Ҫ���á�HIP �ṹ�� HIP ��������״�ء����С���״�غͻұ���ɡ������Ҽ�״������������½Ŷ�����ȫ�γʹ��Ρ����� HIP ������ϸ���������ٽ��仮��Ϊ 4 ���������ֱ�����Ϊ CA1��CA2��CA3 �� CA4����CA4 �������������ӡ���״�ص�Ƥ�ʷ�Ϊ���Ӳ㡢������Ͷ��β㡣��ʵ���о������ʾ��GHS-R��������ϸ���� HIP �������зֲ���HIP �ڽ��ʽṹ�����ڱ�Եǰ��Ƥ�ʽṹ����Եϵͳ����һ�������Ľ���ѧ������ʵ�壬���ǹ���ѧϰ���顢�����½���������飬ͬʱ�����͵���������Ϊ����з�Ӧ�ĸ�����·����Ҫ��һ���֡��½��о����֣�ghrelin ��ͨ�� HIP ���Ӵٽ�θ�������á������о����֣�ghrelin ��������ϸ���� HIP �� CA3 ��������࣬GHS-R ��������ϸ���� CA3��CA4 �����й㷺�����ˣ��������Ʋ��� ghrelin ������������������У�HIP ���ܷ�����Ҫ���á�
�������ʺ��DZ�Եϵͳ����Ҫ��Ƥ���º��ţ�λ���Ҷǰ�����������½Ǽ���Ϸ����ɶ�����ʺ��ܲ�ͬ�ĺ�����ɣ���Ҫ��ΪƤ���ڲ��Ⱥ���������Ⱥ���Լ�ǰ��������Ƥ�������� 2 ������������ӽ���������Ƥ���ڲ��Ⱥ�ȽϹ��ϣ���������Ⱥ���¡�
�������ʺ����Ըɡ����ԡ������Լ�Ƥ��о����ȴ��ڹ㷺����ά��ϵ���Ӷ��漰����������Ҫ�������ܵĵ��ڻ��ƣ�����������Ӧ��ѧϰ���䡢��ֳ��Ϊ�ȸ�����Ϊ�����ϣ�����ʳ��Ϊ��θ���˶��ĵ�������Ҫ���á��½��о�֤ʵ�����ʺ�����ע�� ghrelin���������ƴ���ʳ�����ٽ���ʳ�����о�ʵ����������ghrelin �� GHS-R ��������ϸ�������ʺ˸��Ǻ��ž��б���� Me ��࣬CeA ����; ���ʻ�������Ҳ�н϶� ghrelin ��������ϸ������ˣ���ʾ Me�����ʻ������˺ܿ����� ghrelin ����θ���˶�����Ҫ���ಿλ֮һ��
��������������ghrelin �������� GHS-R ������������㷺���ڣ���Ҫλ�ڴ���Ƥ�ʡ������ԡ�HIP�����ʺ˵Ȳ�λ��Ghrelin ������Ϊ���ڵ��ʹ㷺��������������������Ӹ��������������á�������������������ص����������߱������϶࣬�� HIP�����ʺ˼������Ծ���ǿ���Ա������ʾ ghrelin����ͨ�� GHS-R ����Щ���������������������˶���

��ԭ�������е�ѹ���������Ắ�ȶ����ص�,��Щ����Ӧ���ض����»������������л�ı仯[1].���о�����,�����ڸ�ԭ�ϵ���Ѫ��ˮƽһ��ϵ�,���ķ�����Ҳ��Խϵ�[2].ƽԭ�����������쳣���߽����ԭ����,������״�������仯,��ҩ����Ӧ����ʵ�����,...
����ER������������幹��������rBMSCs�еı�
0����.�Ƽ��������ǴƼ��������ڰ����ٵ���Ҫ·��֮һ.�Ƽ���ˮƽ���Ƽ����������IJ���,����ʹϸ�������ԴƼ��صIJ�ͬӦ��[1-3].�Ƽ�������Ϊ������������̴��������Ա֮һ,��,2������,���и߶�ͬԴ��,�����ܲ�ͬ[4].������ʸ�ϸ�����и߶�����...
������״������������������Ԫ�ķֲ�����
�����ڵ��ڻ����˶�����֪���ܷ�����м�����Ҫ������[1].��״���ǻ��˵���Ҫ���ţ���������ѧϰ������ص���֪����[2],��㷺���ܴ���Ƥ�ʡ����Ժ����Ե��˷��Դ���[3].��״��������ܲ�ͬ����Ϣ����ζ�Ÿ����в�ͬ����������[4].��״��������������...
Ӧ��RNAi����NSCs��NgR�ı���������������
Ŀǰ��Ӧ������֯����ѧ�ķ������ٽ��������������ѳ�Ϊ�����о������ȵ㣬������ǰ��ʵ����Ӧ����ϸ����������������������Σӣã���ֲ�������յ�����������ȱ��ȡ�õIJ�����Ч��������ʵ���з�����Դ���������...