����������
�������������ҽѧ�����Ľ��������Խṹ���ܳ�����Ի�ø��Ӿ�ȷ�����ݣ�ͬʱ�Դ��Ե��о����پ����ڴ����ӽṹ����̬���ܣ�����ע�ظ��ӽṹ֮�����ϵ����ˣ������������۳�Ϊ�о����Ե��������ߣ���������������Դ�ڶԹ�����������������о���С����ģ�ͺ��ޱ��ģ�͵����ʹ���������������չ��ʹ��������Ĺ۵㿪ʼ�������ǵ������������۵���Ϊ����������Է�Ϊ�ṹ����������磮����������ʱ�����������Ϊ�㼯�ͱ����ڵ�Ķ����Ƕ��εģ���Ԫ����Ԫ��Ⱥ����Ƥ�㹦������ȶ����Գ���Ϊ����Ľڵ㣬����ı߱�ʾ�����ڵ�֮����ڵ�ij�ֹ�ϵ���ṹ���練ӳ����Ƥ�㲻ͬ��������̬ѧ�ϵ������Ի������ӵĶ��٣��������練ӳ����Ƥ���������Ԫ��Ķ�̬Э���ԣ��ڵ�Ķȡ�ƽ���ȡ��ȷֲ�������·�����ȡ�����ϵ���ͽ������Ƕ�������������������о��������ԵĽṹ�������綼���ֳ�С�������ԣ���ƽ��·�����Ⱥ̣ܶ��ӽ���������ƽ��·�����ȣ�������ϵ��ȴ���������ߵö࣬�ӽ���������ľ���ϵ�������ҽṹ����Խڵ�����ֳ��������ޱ������Ŀ����ԣ����������о���һ����Ҫ����������ṹ���ܵĹ�ϵ���������о�Ҳ�����⣮����Ƥ����������Ҫ����֯ԭ�ṹ�ָ�������ϣ���������֯ԭ��ʹ���Դﵽһ���ֲ�רע������Э����“����”�Ĺ���ģʽ��ʵ��������ԭ��Ҫ��ṹ�����������֮���Э����ʹ���Գ�Ϊͳһ���壬��˶��������о����ԣ��ṹ����֮��Ĺ�ϵʮ����Ҫ���Թ�����������˽ṹ���Ի���Ϊ���������仯������ﻼ�������δ����ʱ����������������������ԣ�����ʱ�����������Ϊ���ڹ�����������ԣ����Ⱥ�Ĭ֢���ߵ��Թ��������������������˵�������˻�������ԵĽṹ�����������Ĺ�ϵ�Դ��Լ������о�Ҳ������Ҫ���壮�ԵĽṹ���������ϵ�о���Ҫ�����ַ����������Խṹ���ܳ����о�ͬһ����Ľṹ����������磬�������ڳ����ֶκͱ��о����壮���ڽṹ����ģ�ͺ���Ԫģ�͵��о����������ַ�����ȫ����ģ�ͣ�����ȱ���������壮�����ۺ����������ַ���������ͬʱ�����Ե�ʵ��ṹ�������Ԫ����ģ�͵ķ��������ַ��������ǰ������˼·���ŵ㣬��˵õ��Ľ��Ҳ���������ŷ����о��в��õĽṹ������ͨ������ѧ�õ���è��Գ����Ƥ�����������磮�����еĽڵ������ԪȺģ�ͣ�ͨ����ԪȺģ�����Խṹ�����б��ֳ��ķֲ�ͬ���о��Խṹ��������Ĺ�ϵ��
������è��Գ�����Ƥ��ṹ����
���������Խṹ������Է�Ϊ�������裺
��������ڵ㣺��������ͼ����Ƥ����з�����ÿ��������Ϊһ���ڵ㣮�����������ӣ����ڽṹ�Ź���Ľṹ���磬��������ڵ���̬ѧָ��֮���ͳ�ƹ�ϵȷ�����ߣ�������ɢ�Ź���Ľṹ���磬������ά���ټ���ȷ������ڵ�֮��������ӣ����ڽ���ѧ�Ľṹ���磬���ݷ���֮��������Ӷ���ȷ�����ߣ����IJ��õ��Խṹ������ͨ������ѧ�õ��IJ��鶯�����Ƥ�����������磮����è���Խṹ�������ɣ�����������������ͬ�ܶȵ���������ɵ�һ�������Ȩ�������磬�ߵ�Ȩ��Խ��˵�������������������άԽ�࣮Գ����Ӿ�����ṹ������ͨ��ע�������õĽ���ѧ�ṹ���磬���У�����������������ͻ���ӣ�Գ����Ƥ���Ӿ�����ṹ������Գ����Ƥ��ṹ�����һ�������磬ͬ����ͨ����ʾ����Ĺ۲���Խ���Գ����Ƥ�����磬��������У������ڵ㡢�����������ߣ�Գ�����Ƥ��ṹ���綼��������Ȩ���磬չ�ֳ��Ե����ϵ���ͨ�Բ㼶�����۲����������õ���Ԫ�������磬���м��������ͻ���;ֲ��������������磻�ں�۲�����ɴ������������άͻ�ֹ������磬��һ����߶Ȳ��Ҿ߱��㼶�ṹ�����磮
������������ԪȺģ�͵��Թ��ܻģ��
������������Ԫ������֮���ϡ�����ӹ��ɣ����ж��ص�ʱ�䶯̬���ԺͿռ�߶ȣ����۵���۵ĸ���ηֱ�����Ԫ����Ԫ��Ⱥ����Ƥ���ض�������Ƥ����ģ��֯��ȫ�ԣ�ÿһ����ζ����������� ʱ�����ԣ���˴Ӷ��ز�����֣��÷� ��ͬ���ķ���������ͬ���������Ľṹ�Ͷ���ѧ��Ϊ������������ع۲����˽ṹ���������Լ��˽�����֮��Ĺ�ϵ���ö���ѧ�����о��ڲ�ͬʱ�ڵ��Ի������Ҫ����һ���Եļ���ģ�ͣ��۲���ʵ��Ƥ�����������ͬ����Ϊ�Լ���ϵͳ�����ϵ�ȫ�ֶ�̬��֯�ͻ֮��Ĺ�ϵ�����IJ�����ԪȺģ��������Ԫ��Ⱥ��ƽ�����Ȼ��ͨ��ģ����ʾһ��������ȷ�ĵ���ʱ��߶ȵĶ���ѧϵͳ����ڵ������ǿ����ѭȫ���������˲�����һ������ѧģʽ���������̲������¶��ϵķ�����ͨ���Ӱ�����Ԫ��Ⱥģ������ڵ㣬ͨ���ֲ�ͬ����ʾ����ѧ�������Խṹ��������Ĺ�ϵ��
������������ԪȺģ��
������ԪȺģ�͵���Ҫ�����Ƿ�ӳ��Ԫ��Ⱥ����ѧ��Ϊ��һ����Ԫ��Ⱥ���������Ӽ����Ӽ���������ϸ���飬���������Ӽ�����ϸ���źŷ������Ӽ������ɾֲ�������м���Ԫ��ɣ�����ȡ�˷��źź������˷��źŵ�ϸ�����������մ̼��������ģ����Ҫ������Ⱥ�ĺ�۱�����
���������ʾ��ϸ��ͻ�����ƽ��Ĥ���ƣ������ʾ�����˷ܵ��м���Ԫ��ƽ��Ĥ���ƣ������ʾ�����˷ܵ��м���Ԫ��ƽ��Ĥ���ƣ��������ԪȺģ�ͺ�Զ�������ԪȺģ�Ͷ�����һ��ȫ���ԵĴ̼��У���������������DZ�ܵ�ƽ���ܶȣ����뾲̬�����ԣӣ������亯���棨������ƽ��Ĥ��λDZ��ת��Ϊ��ӦDZ�ܵ�ƽ�������ܶȣ�
����ʽ�У��声�ͣ�����Ӧ�ڵ�λ�ķ����ʱ�ʾͻ�����DZ�ܣ����ʾ�ӣ������亯���Ķ��ȣ�ģ������ϸ�����м���Ԫ֮��Ľ�������ͨ���ĸ����Ӳ����ã����ã����ã����ã������������Ǵ���ͻ�����ӵ�ƽ����Ŀ���˷ܺ����Ƶ�����ת������������ͬ����ʽ���������£�
����
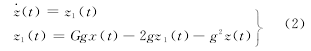
����ʽ�У����˷�״̬�£��ǣ������磽�����״̬�£ǣ��£��磽�⣮�ڶ��Թ��ܻ����ģ��ʱ�����Խṹ�����еĽڵ㿴������Ԫ��Ⱥģ�ͣ����ڵ�֮�����Ϲ�ϵ���Խṹ����ȷ�����ɣθ������ԪȺģ���ɵ�ϵͳ�У��ڣɸ����ӵĶ���ѧ����Ϊ��
����
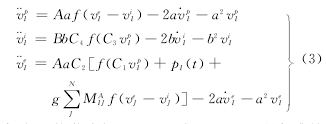
����ʽ�У����ӵļ���Ϊ��ɣ��������𣰣�ξ�ɣ�������ξ�ɣ������DZ����Ϊ���ĸ�˹���������𣰣�����������Ϊ���Ӽ�����ǿ�ȣ���ԪȺģ���еĸ�����ȡֵ�������ʾ��
�����������������ӵĶ���
�����Թ�������������Dz�ͬ��Ƥ��������ź�֮��Ķ�̬��һ���ԣ�����һ���Կ������ʱ���ϵĹ����Ժ�ͳ�������Զ�������Ҫ��Ϊ���Ժͷ��������࣬���Է�������Ƥ��ɭ��ء�ƫ��أ�ʱ��ƫ��ɣ�Ƶ�ȣ������Է�����Ҫ��ͬ����Ȼ�ԡ�����Ϣ�ȣ����Ľڵ��ͣ�֮��Ĺ������Ӷ���Ϊ��Ӧ��ɢʱ�����У֣ɣ�����ɣ�����ɺ֣ͣʣ�����ʣ�����ʵ�Ƥ��ɭ������
����ʽ�У��ֺ�σ�ֱ��ǽڵ��ʱ�����Уֵ�ƽ��ֵ�ͱ��
������������ֵģ����
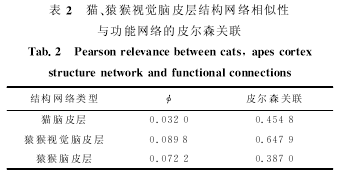
�������Ľ�����è��Ƥ�㡢Գ���Ӿ�Ƥ���Գ����Ƥ��ṹ�����ϵ��ģ�⣮�����������ԪȺģ�͵ij�ֵȡ�����Ͼ��ȷֲ������ֵ��ͨ�����ףң����壭�ˣ��������㷨������ֵ��⣬����ʱ����Ϊ����������ģ���ˣ�����ʱ�䳤�ȵ������ͼ����ͼ����ͼ���ֱ������è��Ƥ�㡢Գ���Ӿ�Ƥ���Գ����Ƥ��ṹ����Ͷ�Ӧ���Թ������Ӿ���
è��Գ�����Ƥ��ṹ���綼���ֳ�����ԣ����Է�Ϊ�����ڲ����ӽ��ܵ������磬��Щ�����������ִ����ͬ���ܵ���Ƥ�������ڶ�Ӧ�Ĺ������Ӿ����У��������ڲ��ڵ�֮���кܸߵĹ���ǿ�ȣ�������Щ�������ڲ����ͬ���̶ȸ��ߣ�������֮���ͬ���̶Ƚϵͣ���ԪȺģ�ͱ��ֳ��ֲ�ͬ����Эͬ����ѧ��Ϊ����ԪȺģ�ͷֲ�ͬ������Ƥ��ṹ�����νṹ���ǺϽ�ʾ����Ƥ��ṹ������������һ���ԣ��Խṹ����������������Թ��ܵĻ�����Ϊ�˽�һ���о��ṹ������������νṹ֮���һ���ԣ����Ļ��ڽṹ�����нڵ�������Զ��������˶���֮������Ƴ̶ȣ��̣������Ȼ��ڽڵ��ھӵ��غϳ̶ȶ����˽ڵ�������ԣ�����������Ծ���ӿ���ͨ����ʽ���㣺
����
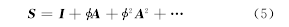
����ʽ�У�����������ڽӾ����ǵ�λ���ǿ��Ʋ�����Ϊ��֤����������ӦС�ڻ���ھ�������������ֵ�ĵ��������Ƿֱ����è��Գ���Գ���Ӿ���Ƥ��ṹ����������Ծ���Ȼ�����д洢Ϊһά���м���Ƥ��ɭ�������������ߵĽṹ���������Ծ����������֮���һ���ԣ�������ʾ���ڲ�ͬ�Ħ��ֱ�ȡè��Գ���Գ���Ӿ���Ƥ��ṹ�����������ֵ����ʱ�������Ծ����������һ���Ե��������������ӱ������Կ����������Ծ����������֮�����������Ҿ��нϸߵĹ���ֵ��˵�����߾��кܸߵ����ƶȣ���ֵʵ��Ľ��˵����������ͬ����Ƥ������ͨ��һ�µ���ΪģʽЭ�������ض������ͬ���ܵ�����֮����ֳ���ͬ����Ϊģʽ����Ƥ��ṹ���ܵķָ�������ͨ���ֲ�ͬ����Ϊ����ʵ�֣�
����
����������
��������ͨ�������ԪȺģ������Ƥ��ṹ�����ϵ�Эͬ����ѧ��Ϊ�о����Խṹ��������֮��Ĺ�ϵ����è��Ƥ��ṹ���硢Գ���Ӿ���Ƥ��ṹ�����Գ����Ƥ��ṹ�����ϣ���ԪȺģ�ͱ��ֳ��˷ֲ�ͬ����Ϊ��һ���̶��ϱ����ֲ�ͬ���е�ͬ������ṹ�����νṹһ�£�֤���ԵĽṹ����������Ļ������Թ��ܵķָ������ͨ����Ԫ��Эͬ�ʵ�֣������ܹ�����˵���ԵĽṹ�����빦������Ĺ�ϵ�������������Ե��������Ҫ���壮
�����ο����ף��ң���������
�����ۣ��ݷ�С�ᣬ�ں�࣮�����������о���չ�ۣʣݣ���ѧ��չ����������������������������������������
�����ۣ�����С�������裬�¹��٣������������ۼ���Ӧ�ãۣͣݣ��������廪��ѧ�����磬����������
�����ۣ��ݣӣ����ϣ��ԣ����顡�ǣ��ˢ�������ң��ԣ�塡�������������������壺������������������졡��������������桡����塡�������������ۣʣݣ��У̣�ӣã���������������졡�£��������������������������������������������
�����ۣ��ݣ٣����硡�͡��У��ԣ�塡����������������桡�������졡������������������������塡��������졡���������ۣʣݣ��У���������£���������ӣ�����������������������������������������������
�����ۣ��ݣף����塡�ʡ��ǣ��ӣ�����������塡�ţ��ԣ�������ʡ��Σ��������죮�ԣ��������������塡��桡����塡���������������������桡����塡��������������������������������ۣʣݣ��У������������졡�ԣ������������£��������졡�ӣ�����������������������������������������������
�����ۣ��ݣӣ�������ң��ӣ�������硡�ʣ��ã�������͡��ң��������죮�Σ������������������졡���������������塡��桡������������졡�������������������塡��������桡�������������ۣʣݣ��ã������졡�ã������������������������������������������������������
�ۣ��ݣӣ������졡�ʡ��ף��٣����硡�͡��У� �ԣ�塡�����������������������������桡�������졡���������������塡��������������졡���������ۣʣݣ��ã����������£��������������������������������������������
�����ۣ��ݣӣ�������ˣ֣��ʣ���ң��ӣ���������ӣ��������죮�ӣ����죭���������������롡��������������桡������������졡���������졡���������롡�����������������������������������塡�����������������졡��桡����������ۣʣݣ��ţ��������ʣ������졡��桡�Σ�����������壬������������������������������������������������
�����ۣ��ݣ������䡡�ӣ��£��������塡�ţ� �ţ��������������䡡��������������������졡�����������������졡�����������ۣʣݣ��У̣�ӣã���������������졡�£��������������������������������������������
�����ۣ����ݣУ������ӡ��ã��ģ����������������ȣ��������䡡�����������죮�ԣ��������������𡡣����������������������졡���䡡������������졡�������������������ǣ���衡�������������졡����������桡���ţţǡ��������졡������������ۣʣݣ��Σ���ɣ����壬��������������������������������������
�����ۣ����ݣӣ������졡�ʡ��ף��£����ǡ������С��ã��ȣ�������硡�á��ã��������죮�ԣ�塡�ã�����������졡����������������桡����塡�������������������������������桡����塡�����ۣʣݣ��ã������졡�ã��������������������������������������������
�ۣ����ݣ٣����硡�͡��У��ԣ�塡���������������塡��桡��������졡�������������������������졡������������������ۣʣݣ��ӣ������졡�֣����������������������������������������������
�����ۣ����ݣƣ��������ġ��ʣ������ţ����ġ��ã��ģ������������䡡�ȣ�����������У��������硡������塡�У�������塡�ã������졡�ã�������ۣʣݣ��ã������졡�ã��������������������������������������
�����ۣ����ݣף������硡�ƣ��£�������ʡ��ʣ��£����������顡�ƣ��������죮�ң��������塡��桡����������������䣭��������������������������������桡������裭�ţţǡ�����������㡡��������ۣʣݣ��£���������ã������������������������������������������������
�����ۣ����ݣڣ�������ᡡ�ͣ��ã��ᡡ�ƣ��գ�����ͣ������������졡������������������������塡������������������������ᡡ���������졡�������ۣʣݣ��ã���������������졡�ɣ�����������塡���䡡�Σ�����������壬���������������������������������������������������ۣ����ݣȣ����ģ������������������桡������������������������졡��������������ۣʣݣ��ӣ̣��͡��ң�����������������������������������������������ۣ����ݣ̣��������š������ȣ����塡�У��Σ�������͡��š��ʣ��֣����������������������������������ۣʣݣ��У�������졡�ң���������ţ�������������������������������������������������