生理学论文


近年来,随着分子生物学的发展,情绪个体差异的遗传背景受到越来越多的关注。研究发现,情绪的产生和调节与神经递质代谢酶基因多态性有关。单胺氧化酶( MAO) A 是单胺类神经递质的主要代谢酶类,MAO A 多态性与攻击行为、焦虑症等关系密切,动物研究也证实 MAO A 多态性与攻击性有关。本课题组前期研究也发现 MAO A 多态性与愤怒情绪的产生与表达相关。攻击性行为和愤怒、抑郁情绪等的产生均与人的执行功能( EF) ,尤其是抑制控制密切相关,那么 MAOA 多态性对愤怒等负性情绪的影响是否因为其对抑制控制能力的调节呢? 本文选择事件相关电位技术( ERPs) 考察不同MAO A 基因型的被试者抑制控制能力是否不同。
1 材料与方法
1. 1 对象 以某学院在校正常大学生为研究对象,经伦理委员会批准后招募被试,填写知情同意书及一般情况调查表。
1. 2 试验程序和任务 刺激由 E-prime 2. 0 系统控制呈现。参考文献,以黑色单/双三角形构成 Go/Nogo 任务刺激图形,共 8 组刺激,每组含 100 个试次,其中 Go 任务为 60 个,Nogo 任务为 40 个。要求被试 Go 任务时做按键反应。按键左右手在被试者中交叉平衡。整个实验约需要 20 min,实验完成后给予被试一定酬劳。
1. 3 数据采集与处理 采用 NuAmps40 导系统 ( NeuroscanInc) 记录脑电。同步记录连续脑电图( EEG) 与行为学数据,用Neuroscan4. 5 软件对脑电数据进行离线分析。对反应正确的EEG 进行分类叠加,得到 Go 和 Nogo 刺激产生的两类 ERP 数据。对得到的 ERPs 进行 30 Hz( 24 dB/oct) 无相移低通数字滤波。
1. 4 基因分型
1. 4. 1 多态性位点的选择 根据报道,从人类基因组单体型图( HapMap) 中选出能代表亚洲人 MAO A 基因单核苷酸多态性( SNP) 阳性信号的标签 SNP rs5905613 基因多态性位点,考察其多态性( CC、CT、TT) 对抑制控制能力的影响。最终各基因型入组被试 CC 11 人,CT 9 人,TT 16 人。
1. 4. 2 PCR 扩增目的片段 以白细胞基因组 DNA 为模板,扩增目的基因片段。正义引物 CCACAATAGATAACCTTGGGC,反义引物 CCTGGGCCAAAAATGCTATTC,扩增片段长度 222 bp.PCR 反应体系: 模板 DNA 1 μl,10 倍 PCR 缓冲液 1. 5 μl,MgCl2( 25 mmol/L) 1. 5 μl,dNTP( 10 mmol/L) 0. 3 μl,Taq 酶 1. 25 U,引物各 0. 25 μl,使用去离子水补足体积至 25 μl.反应条件:94℃ 预变性 2 min; 94℃ 变性 15 s、60℃ 退火 15 s、72℃ 延伸 30 s共 35 个循环; 72℃延伸 3 min.扩增完成后 1. 2% 琼脂糖凝胶电泳获得目的核苷酸片段。各组被试一般情况( 年龄、学历、性别) 无显着性差异,具有可比性。
1. 4. 3 SNP 位点连接酶特异检测反应( LDR) 针对 SNP 位点设计识别两种不同碱基的左端探针( 长度相差 3 bp) ,以及带羟基荧光素( FAM) 的右端共用探针,终产物长度差别即为左端探针的长度差别。LDR 探针序列为右端公用探针 P-TGTAC-CATTTGTACTTGAGCCAGCATTTTTT-FAM, 左 端 探 针 TCTTTTTTTTTACCAAATTCCTCACTATCACAGGC,连接产物为 64 /C,左 端 探 针 TT TTTTTTTTTTTTACCAAATTCCTCACTAT-CACAGGT,连接产物为 67 / T.连接体系: PCR 产物 3 μl、10 ×Taq DNA 连接缓冲液 1 μl、Taq DNA ligase 5 U,特定 LDR 探针各 0. 1 pmol,去离子水补足体积至 10 μl.连接反应参数: 94℃变性30 s、60℃退火并连接3 min,循环20 次。反应结束后,4℃保存待用。取 1 μl 连接反应产物,加 10 μl 上样缓冲液( 已混入 Marker) ,95℃ 变性 3 min,立即冰水浴。连接产物经 ABI3730XL 测序仪扫描,根据目的峰与 Marker 的位置差距来读取结果。
1. 5 统计学方法 导出行为学数据( 反应时、正确率、漏报率、错报率) 进行单因素方差分析。本研究主要对 EEG 结果中线的五个电极位点进行统计分析。以刺激出现后200 ~300 ms 时间窗内最大负波峰值及其出现时间为 N2 幅值和潜伏期、300 ~500 ms 时间窗内最大正波峰值及出现时间为 P3 的幅值和潜伏期。为了强调 Nogo 效应和进行组间比较,研究使用相减技术从 Nogo 成分中减去 Go 成分,分别获得 N2 和 P3 的差异波( N2d 和 P3d) ,以刺激出现后 200 ~ 300 ms、300 ~ 500 ms 时间窗内 N2d 和 P3d 的面积作为 Nogo 效应的指标.分别对 N2、P3 峰值及差异波出现的潜伏期和峰值波幅进行混合重复测量数据方差分析。重复测量方差分析的 P 值采用 Greenhouse-Geisser 法校正。所有数据均采用 SPSS19. 0 软件进行分析。
2 结 果
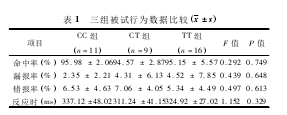
2. 1 一般情况及行为学数据 各组被试一般情况中年龄〔CC组( 19.50 ± 0. 67) 岁; CT 组( 19. 44 ± 1. 13) 岁; TT 组( 19. 71 ±0. 92) 岁; F = 0. 311,P = 0. 734 〕、学历 ( 均为大学本科) 、性别( 均为女性) 无显着性差异。三组被试的命中率、漏报率( Go 任务) 和错报率( Nogo 任务) 差异均没有统计学意义,反应时差异也没有统计学差异。见表 1.
2. 2 三组被试在不同刺激条件下产生的 N2、P3 成分峰潜伏期比较 各组间 Go 和 Nogo 刺激产生的 N2 成分组间及反应类型间差异均不显着〔F( 2,33)= 1. 057,P = 0. 359,η2= 0. 060; F( 1,33)=0. 158,P = 0. 693,η2= 0. 005〕; 各组间 Go 和 Nogo 刺激产生的 P3成分潜伏期组间差异不显着〔F( 2,33)= 2. 523,P = 0. 096,η2=0. 133〕,但反应类型间差异显着〔F( 1,33)= 4. 778,P = 0. 036,η2=0. 126〕,事后检验显示 Nogo 效应峰潜伏期( 390. 18 ± 5. 35) ms 较Go 效应峰潜伏期( 379. 11 ± 6. 68) ms 慢。见表 2,表 3.


2. 3 波幅 三组被试在不同刺激条件下产生的 N2 成分波幅,在反应类型、脑区及电极位置方面均存在明显的主效应〔F( 1,33)= 63. 932,P = 0. 000,η2= 0. 660; F( 2,66)= 9. 129,P =0. 000,η2= 0. 217 和 F( 4,132)= 25. 961,P = 0. 000,η2= 0. 440〕。
脑区和分组间存在交互效应〔F( 4,66)= 3. 217,P = 0. 019,η2=0. 163〕,简单效应分析表明同一脑区三组被试间波幅差异不显着,但同一组被试三个脑区间波幅差异显着,以中线区波幅最负。见表 5.
三组被试在不同刺激条件下产生的 P3 成分波幅比较,组间比较差异显着〔F( 2,132)= 3. 722,P = 0. 035,η2= 0. 184〕,成对比较 CT 组 波 幅 〔( 11. 29 ± 1. 27) μV〕比 CC 组 〔( 7. 07 ±1. 15) μV〕及 TT 组〔( 7. 50 ± 0. 95) μV〕波幅大。在反应类型、脑区和电极位置均存在明显的主效应〔F( 1,33)= 6. 428,P =0. 016,η2= 0. 163; F( 2,66)= 7. 227,P = 0. 003,η2= 0. 180 和F( 4,104)= 5. 641,P = 0. 012,η2= 0. 146〕。成对比较表明 Nogo 条件下的 P3 波幅大于 Go 条件下的波幅,中线位置的电极波幅最大,左右脑区无差异,最大波幅分布在额中央区。见表 4,表 5.
中线部位五个电极的 N2d、P3d 面积组间差异均不显着〔N2d: F( 2,33)= 1. 377,P = 0. 266,η2= 0. 077; P3d: F( 2,33)=0. 327,P = 0. 723,η2= 0. 019〕,电极位 置差异显着〔F( 4,132)=18. 722,P = 0. 000,η2= 0. 362〕,成对比较显示额区和额中央区差异波面积最大,与其他 3 个位点间差异显着。见表 6.
3 讨 论
本研究从行为数据上看,三组被试的 Go/Nogo 任务的行为反应无明显差异。Go/Nogo 任务十分简单,这可能是三组被试的任务表现无显着性差异的原因。从 ERP 数据来看,被试在Nogo 试次的 N2 及 P3 波波幅增大,成功地诱发出了 Nogo 效应;Nogo-P3 最大波幅分布在额中央位点,出现前部化效应。虽然N2 和 P3 差异波的差异不显着,但 CT 组被试 Go 和 Nogo 试次的 P3 波幅较 CC 组和 TT 组被试均显着增大,显示三组被试在认知信息加工处理过程中具有不同的脑电活动特征。
执行功能是指在完成复杂的认知任务时,对各种认知过程进行协调,以保证认知系统以灵活、优化的方式实行特定目标的一般性控制机制; 其本质就是对其他认知过程进行控制和调节.执行功能的主要成分包括抑制控制,转换和刷新等,其中抑制控制是执行功能的核心成分.个体愤怒特质与攻击行为关系密切,研究表明高特质愤怒个体在敌意情境下会表现出更多的行为攻击倾向,而国内外研究均表明攻击行为等与执行功能尤其是抑制控制功能呈负相关。ERPs 具有较高的时间分辨率和一定的空间分辨率。大量研究表明,Go/Nogo 任务可以用来检测抑制控制功能。在 Go/Nogo 任务中被试需要对靶刺激做出反应( Go) ,而抑制对非靶刺激的反应冲动( No-go) .
该 任 务 产 生 两 个 主 要 脑 电 成 分,第 一 个 是 波 峰 在200 ~ 300 ms潜伏期内的 Nogo-N2 成分; 第二个成分是波峰在300 ~ 500 ms潜伏期内的 Nogo-P3 成分。这两个 ERP 成分都被认为与反应抑制活动有关。Nieuwenhuis 等报告 N2 波幅在低频刺激下增大且定位于前扣带回,并据此认为 Nogo-N2 代表了信息输入及加工过程中干扰信息的监测。Smith 等发现被试对 Go 刺激反应越快,诱发出来的 Nogo-P3 就越大。Kok等则报道在 Stop-Signal 任务中,正确的 Nogo 试次明显比错误的 Nogo 试次诱发出来的 Nogo-P3 大。
因此,前中央部需要抑制反应的试次诱发出来的 Nogo-P3 增大被认为是反映了抑制过程.尽管两者的心理过程没有最终确定,但可以认为 Nogo-N2 和 Nogo-P3 可能分别代表了与抑制控制有关的两个加工过程: 冲突监测和反应抑制。虽然本研究中三组被试的 N2d 和P3d 均未发现不同,即未发现三组被试的抑制控制能力存在显着差异,但三组被试在 P3 波波幅的显着差异,表明不同 MAO A基因多态性对大脑信息加工和处理过程存在影响,CT 多态性被试在加工处理信息过程中产生了更显着的电活动,动员了更多的认知资源。

单胺氧化酶(MAO)是一种降解生物肽的酶[1],分为两种亚型,单胺氧化酶A(MAOA)和单胺氧化酶B(MAOB)[2].MAO-A对底物血清素(5-HT)、去甲肾上腺素(NE)、多巴胺(DA)和抑制剂氯吉兰具有高度亲和性[3];单胺氧化酶B是一种线粒体黄素蛋白酶,其作用主要...