摘要:外来植物入侵对全球生态系统构成严重威胁,尽管入侵植物的管理与控制取得了一定进展,但要恢复被入侵生态系统、完成管控目标仍存在问题。本文对入侵植物管控成效的制约因素进行了综述,发现入侵植物管控过程中存在以下问题:(1)预防为主的管理原则不完全适用入侵管理;(2)入侵植物管理存在缺乏公众支持、生态责任主体单一及多头管理等缺陷;(3)入侵植物管理目标与社会管理目标及生态环境保护目标矛盾;(4)入侵植物控制措施成本高昂,易受环境因子限制,并产生负面生态效应;(5)目标入侵植物控制后可能产生入侵遗留效应并发生继发入侵。此外,全球变化在加速植物入侵的同时会改变人类观念,进一步增加了管控难度。据此,我们认为,未来应加强大时空尺度下入侵植物生态效应、入侵生境恢复、管控决策等方面的研究,以提升入侵植物管控成效,恢复植物入侵生态系统。
关键词:生物入侵; 生境恢复; 全球变化;
Restraining factors on the effectiveness of invasive plants management and control
XU Guang-yao LI Hong-yuan MO Xun-qiang MENG Wei-qing
College of Environmental Science and Engineering,Nankai University School of Geographic and Environmental Sciences,Tianjin Normal University
Abstract:Biological invasion is one of the major causes of global biodiversity loss. Although some progress has been made in its management and control,there are still problems in restoring the invaded ecosystems and completing the management objectives. We reviewed the restraining factors on invasive plant management and control. The following problems are found in the management of invasive plants:( 1) the precautionary principle is not fully applicable to invasion managements;( 2) invasion management is facing the problems of lacking public participation,single ecological responsibility subject,and multiple management;( 3) target conflicts exist in management of invasion management and social management and ecological environmental protection;( 4) measures on invasive plants control are costly,susceptible to environmental factors,and common to produce side effects;( 5) legacy effects and secondary invasion emerged after target plants are controlled. In addition,global change may exacerbate plant invasion and change the concept of human beings,which will further increase the difficulty of control. Hence,to enhance the effectiveness of management and control efforts and restore invaded ecosystems,it is necessary to strengthen the research on ecological effects of invasive species,invaded habitat restoration,and management decision at large spatial and temporal scales.
生物入侵被认为是仅次于栖息地破坏导致生物多样性减少的第二大原因,植物是生态系统中最主要的生产者,入侵植物往往会在生态系统中占据优势地位,进而改变生态系统的运行过程(许光耀等,2018b)。尽管不同入侵植物对入侵生态系统影响的程度和方向有所差异,但植物入侵普遍降低了本地植物的丰度(-43.5%)、多样性(-50.7%)和适合度(-41.7%),入侵植物对生态系统的影响还会随着食物链和营养级传递,降低本地动物的适合度(-16.5%)、丰富度(-17.5%)(Eviner et al.,2012)。在全球化的推动下,入侵植物的数量不断增加,Kleunen等(2015)对全球外来入侵植物进行了量化统计,共发现13168种归化维管植物,保守估计其中10%会成为入侵植物。
当前,对植物入侵的多样性与格局、入侵机制及生态学效应、入侵控制与管理等方面的研究已取得重要进展(鞠瑞亭等,2012),但入侵植物的研究最终都要面向实际应用。植物入侵是事后概念,入侵植物的认定是基于其造成的负面影响,但很多具有入侵潜力的外来植物,在成功入侵前存在较长的潜伏期,导致入侵植物早期识别、早期控制异常困难(Simberloff,2014),使得入侵植物的防控成为入侵生态系统恢复的最后手段。入侵植物的控制经验来源于农业系统的杂草控制,但在更复杂的环境(尤其是自然环境和城市环境)中,光照、地形等环境因素以及文化、经济等社会因素往往制约控制成效(Kettenring et al.,2011)。由于全球气候变化、土地利用方式改变等原因,入侵植物在造成消极影响的同时,往往还提供积极的生态服务功能(Tassin et al.,2015),由于利益或目标冲突导致利益相关者或不同管理部门的管理决策冲突(Mason et al.,2017)。景观规模上的入侵植物管控需要极大的财政投入,但控制副作用、气候变化、遗留效应、继发入侵等因素注定了难以保证入侵控制效果,这进一步增加了管控难度。以全球生物多样性热点地区之一的南非弗洛勒尔角(Cape Floristic Region,CFR)为例,植物入侵使CFR地表径流降低30%以上,上千种本地植物受到威胁,为保护生物多样性,南非政府进行了为期超过20年的入侵植物控制工作,然而,由于小种群监测困难及控制成本太高,入侵植物仍未根除(van Wilgen et al.,2016)。本文综述了影响植物入侵管控成效的问题,以期相关部门予以防范或权衡,为提高入侵植物管控成效、促进植物入侵区域生态恢复提供参考。
1 入侵植物管理过程中的限制
1.1 预防为主的管理原则难以实行
植物检疫是防止外来植物入侵的第一道防线,而检疫名单是检疫的基础。从生态学角度看,除非使用白名单制度(只有入侵性非常低的物种才能引入),否则不可能完全阻止外来植物入侵,但出于经济社会发展的需要,所有国家使用的都是黑名单制度(拦截入侵物种)。即使这样,由于认定标准不统一、动态信息更新不及时等原因导致入侵植物名单很难迅速更新。
1.1.1 入侵植物认定标准不统一
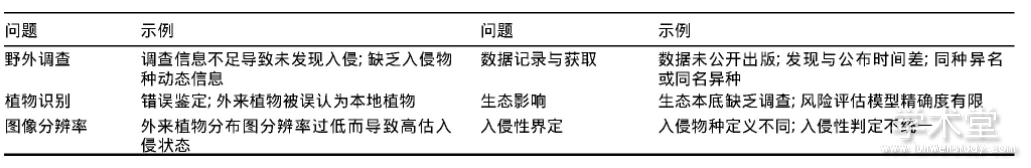
外来入侵物种名单是分析、评估、预防、控制入侵植物的基础,然而,不同学者对入侵概念的认识不同,导致入侵植物的认定标准不明确(Courchamp et al.,2017),因而无法形成统一的名单,这为入侵植物管理造成障碍。植物入侵性通常基于以下5个维度来判断:局部丰度(local abundance)、地理分布幅(geographic range size)、环境分布幅(environmental range size)、扩散速率(spread rate)、负面效应或影响(negative effect or impact)(Catford et al.,2016),定量判断依据的缺乏导致不同研究人员可能将同一地区表现相同的外来物种归类于不同的入侵状态。此外,以下问题使植物入侵的界定更加复杂:入侵植物的引入时间阈值如何界定,几百甚至上千年前传入的植物算不算外来植物?入侵植物应该基于何种边界界定,是地理边界(纬度带)还是政治边界(国界)?若基于政治边界,是否应该包括全球变化导致的、反常的、大范围扩散的本地植物(Nackley et al.,2017)?造成多大的生态、经济或社会损失的外来植物才算入侵植物?
1.1.2 入侵植物动态信息更新不及时
生物入侵是事后概念,只有外来植物造成很大的负面影响时,才能认定其为入侵植物,因而入侵植物名单的更新必然存在滞后性(表1)。由于群落动态性,需要定期调查以保持准确的外来植物信息,但外来植物种类繁多,全球仅菊科(Compositae)、禾本科(Poaceae)和豆科(Leguminosae)植物中,归化物种就近4000种(Py2ek et al.,2017),有限的研究资源不可能分配到每个物种、每个地区。入侵植物被引入后,可能扩散到沼泽、山地等交通不便的地区,并长期处于小种群阶段,入侵植物的实际分布动态很难精准获得(Hyndman et al.,2015)。此外,某些入侵植物与本地同属种或近缘种极其相似,即使专业的植物学家也很难判断,本底调查资料的匮乏和植物分类学的不确定使这种问题更复杂(Hirsch et al.,2017)。
表1 入侵生物名单更新中存在的问题
1.2 入侵植物管理制度存在缺陷
由于自身的生态需求及入侵过程的复杂性,公众很难理解或支持入侵管理。入侵植物渐进式生态影响及多途径扩散方式,使生态损害的责任主体很难确认,管理权责的混乱进一步降低了入侵植物的管理成效。
1.2.1 公众参与制度难落实
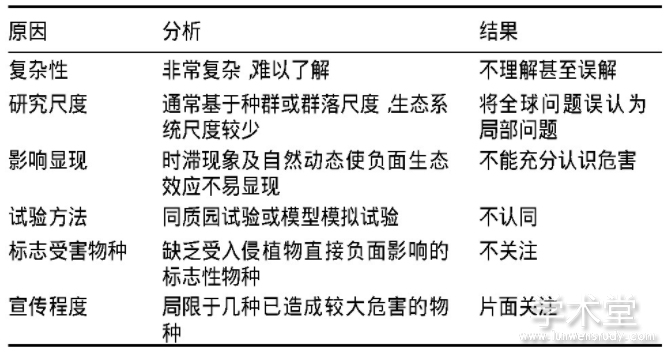
除新西兰和澳大利亚等少数国家外,各国公众很少重视或关心植物入侵。没有生态学背景的公众会简单地认为外来植物进入生态系统不会产生问题,而根除某种植物以保护生物多样性却是荒谬的(表2)(Courchamp et al.,2017)。研究和控制入侵植物的根本原因必然是其造成生态和经济损失,但要从生态系统的自然变化中区分外来植物的渐进式影响非常困难,事实上,即使是对入侵性最强的植物造成的最方便直观估计的经济影响,人们也知之甚少。与其他环境问题相比(如全球变暖与北极熊,过度捕捞与鲸鱼),很少有研究报道入侵植物对珍惜、濒危保护动物的影响,这无疑降低了舆论及公众对入侵植物的关注(Touza et al.,2014)。
表2 入侵植物管理缺乏公众参与的原因分析
1.2.2 生态损害责任主体难确定
植物入侵的责任主体往往只有也只能是国家(政府),90%以上的入侵植物是人为引入或带入的,其中大多数是作为牧草、饲料、蔬菜、观赏植物等被有意引进或是随有意引进的原木、苗木、花钵、土壤而被无意传入(Simberloff,2014),这些植物的引入者大多是园林、农业等政府部门。入侵植物从有意引入到爆发,涉及引入、使用、管理等多个单位或个人,交通、旅游、气候变化、土地利用等因素都会促进入侵植物扩散(MacDougall et al.,2005),这导致生态破坏的责任主体很难确定。从本地植物自然演替及其他人为干扰的过程中区分入侵植物渐进式的生态影响,无疑增加了生态破坏的定损难度。入侵植物形成灾难性后果之前,长期保持无害性,加之其几十乃至上百年的时滞效应(Caplat et al.,2014),事后追责几无可能。
1.2.3 多头管理导致职责不清
管理职责混乱进一步加剧了入侵植物管理的困境,例如,美国维拉帕湾,潮间带的入侵互花米草(Spartina alterniflora)由联邦、州、地方政府以及土地所有者共同负责,各方很难达成统一的管理决定。即便加州海岸环保局强行通过米草控制项目(ISP),十多年努力后却发现濒危秧鸡(Rallusobsoletus obsoletus)已将米草作为栖息地,控制工作不得不中止(Strong et al.,2016)。分割的部门管理方式与生态系统的整体性矛盾,不同部门的目标冲突使入侵植物管理更加混乱,例如,在法国,尽管被列为高度入侵树木,但一些政府机构仍在积极推广种植刺槐(Robinia pseudoacacia)(Dickie et al.,2014)。更有趣的是,世界农林中心(ICRAF)在非洲同时鼓励和反对种植木豆树属(Prosopis spp.)植物(Low,2012)。
1.3 与其他管理目标并非完全一致
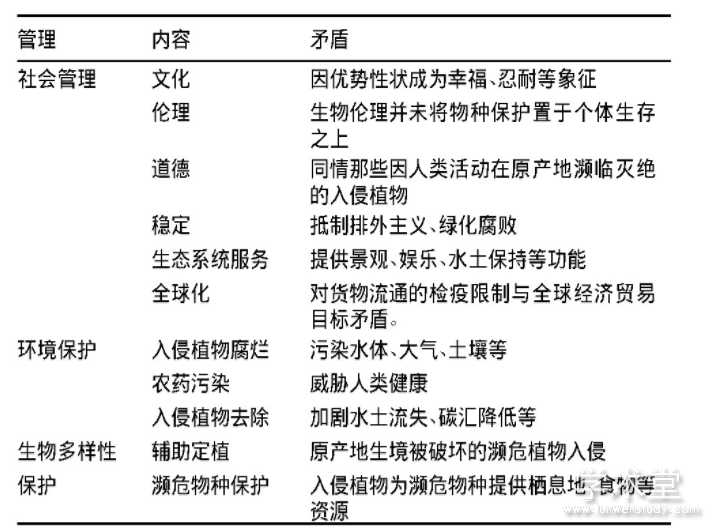
植物入侵并不限于负面影响,在许多自然环境被改变的景观中,入侵植物对生态系统及其服务功能同样具有积极影响,但入侵植物管控往往忽视其积极生态效应(如经济价值、美观价值等),只是试图在尽可能大的面积上消灭或控制入侵植物,导致入侵管理与社会、环境保护等管理目标冲突(表3)。
1.3.1 与社会管理目标矛盾
城市中的外来植物作为入侵生物,成为周边生境的入侵繁殖体来源,同时也作为园林物种,为城市输出景观、娱乐等生态服务功能(McLean et al.,2017),出于对“绿化腐败”的忧虑及自身生态需求,民众往往不会赞成去除城市中的入侵植物(许小过等,2017)。很多城市甚至将入侵植物作为地区标志,例如,南非比勒陀利亚有一个蓝花楹市(the Jacaranda city),阿根廷松海市以入侵松树(Pinus spp.)命名(Dickie et al.,2014)。此外,引入时间较长的外来植物,可能由于其美丽、寿命和耐性等优势性状,渗入当地文化中,例如,藏民将波斯菊(Cosmos bipinnata)视为“格桑花”(幸福花)(周浙昆,2017)。出于补偿或同情心理,许多在原产地濒临灭绝而在入侵地大量存在的外来植物,如米氏野牡丹(Miconia calvescens)、辐射松(Pinus radiata)等,往往被看成物种自发的“辅助移植”,人们在感情上很难支持根除这种植物(Caplat et al.,2014)。这些因素导致入侵植物管理被很多人(包括生态学者)认为是“环境纳粹主义(environmental Nazism)”或“排外主义(xenophobia)”而加以抵制。
表3 入侵植物管理目标与其他管理目标的矛盾
1.3.2 与生态环境保护目标矛盾
许多自然条件被改变的景观中,本地物种往往能从入侵植物中获利(Tassin et al.,2015),在亚热带澳大利亚,入侵植物香樟(Cinnamomum camphora)延长了粉顶果鸠(Ptilinopus regina)食物资源的供应季节(Neilan et al.,2006),在这种情况下,往往需要权衡入侵植物管理与生物多样性保护(Vitule et al.,2012)。此外,入侵管理过程中会产生土壤、水体等污染,将植物入侵的生态问题转变为环境问题。张辉等(2011)调查发现,有20%~70%的除草剂会残留于土壤中,从而使土壤、地下水受到污染;de Winton等(2013)研究发现,物理控制中大约有一半的成本是用来运输和处理收获的入侵植物,为降低费用,收割的入侵植物随意堆放,其腐烂可能产生大气、土壤、水体污染。这个问题已引起相关部门的重视,出于对环境安全的考虑,很多国家限制使用草甘膦等农药控制入侵水生植物(Hussner et al.,2017),但在入侵植物迅速扩散时,化学控制是唯一可行的选择。此外,很多国家最初引入入侵树种是利用其较宽的生态位、较快的生长速度,阻止土壤流失或增加碳汇等(Proche爧et al.,2012),短期内入侵植物管理会直接降低这些生态功能。
2 入侵植物控制过程中的局限
入侵植物的物理、化学及生物控制措施均存在局限,前期调查、环境因子限制、投入费用等都会影响控制成效。入侵植物的管控目标是恢复入侵区域生态系统功能,由于控制副作用、入侵植物遗留效应及继发入侵等原因,即使成功抑制甚至根除入侵植物,管控目标仍很难实现。
2.1 高昂成本导致区域控制不均衡
入侵植物控制总成本可分为发现成本、定界成本、控制成本,3种成本相互影响,且受入侵程度、控制目标和控制时间的影响(Hester et al.,2013)。入侵植物控制成本高昂,采用收割机处理入侵水生植物的成本大约是每公顷2000~4000美元,为了达到较高的去除效率,假设每年清除3次,控制费用将达到每公顷6000~12000美元(de Winton et al.,2013)。此外,根除入侵植物成本远高于抑制扩散所需成本,van Wilgen等(2016)统计表明,南非在过去20年里控制入侵植物的总成本约为5.64亿南非币,只有大幅度增加资金(1.70~26.08亿南非币),才能根除剩余的(4%)入侵植物。经济欠发达地区必然难以维持高昂的控制成本,即便是经济发达的地区,有限的控制成本也要优先用于自然保护区、国家公园等高价值生态系统,区域管理的不均衡给入侵植物留下再次扩散的桥头堡,进而影响入侵植物控制的总体效率(Kerr et al.,2016)。
2.2 环境因子限制控制成效
物理控制易受地形限制,大型机械很难进入山地或林地,即便是水生生境,河道、水位、河底地形等也会影响机械使用。化学除草剂也易受到环境因子的影响,降雨、潮汐等都会降低其浓度,挥发性农药在植物体表面的滞留时间也会受到光照强度、风速的影响(Kerr et al.,2016)。生物控制剂作为生命体,只有在生境筛选中存活下来并发展至一定的种群规模,才能控制入侵植物,且会持续受到气候以及天敌等环境因子的影响。Mahoney等(2017)研究发现,露西苔莺(Oreothlypis luciae)和黄苔莺(Setophaga petechia)会捕食控制入侵柽柳的昆虫,使该昆虫种群增长受到限制。即使生物控制剂能够适应这些条件,由于环境变化,某些区域的治理效果仍不理想,例如,用四重叶甲(Chrysolina quadrigemina)控制入侵贯叶连翘(Hypericum perfortum),森林大火会极大地限制双金叶甲存活和扩散,导致火灾发生区域的治理效率降低(Swope et al.,2012)。
2.3 控制措施易引发负面生态效应
人为干扰是促进外来植物扩散的最主要外在因素,而大型控制机械对生境干扰严重,物理作业会改变原生地貌,导致水体浑浊、土壤压实等问题(Dickens et al.,2016)。入侵植物,尤其是水生植物,很多都能进行克隆繁殖,且其繁殖体具有很强的生存能力,切割、收获、运输很可能促进入侵植物的扩散和传播(Souza-Alonso et al.,2017)。生物控制剂引入前,通常采用输出种群发育法(centrifugal phylogeny method)进行宿主特异性检测,以降低非目标影响,但自然界的潜在宿主不可能完全检测,引入的生物控制剂有可能成为新的入侵物种(Hussner et al.,2017)。
由于控制精度有限,入侵管理往往会对本地生物产生直接影响,例如,Ortega等(2011)使用阔叶除草剂来控制入侵矢车菊属植物Centaurea stoebe时,本地阔叶植物同样受到抑制。残留化学除草剂还会毒害传粉昆虫(齐月等,2016),由于协同进化作用,传粉昆虫种群降低显然对本地植物影响更大。此外,肉食动物能降低入侵植物因天敌释放获得的优势,间接支持了本地植物(Smith et al.,2015),然而入侵控制产生的生态陷阱,会降低本地肉食动物的丰度。例如,Carter等(2017)研究发现,铜斑蛇(Agkistrodon contortrix)更倾向栖息于入侵植物控制生境,但由于管理干扰,这些地方的铜斑蛇死亡风险更高。
2.4 控制不能消除入侵遗留效应
入侵植物控制往往只是简单地在尽可能大的面积上消灭或控制入侵植物,但由于遗留效应,生态系统并不会随入侵植物的控制而恢复(Corbin et al.,2012)。首先,入侵植物与本地植物基因流和杂交影响本地植物遗传多样性,Bleeker等(2007)发现,德国有17种本地植物因与入侵植物杂交而遭受远交衰退(outbreeding depression),8种植物基因库受基因渐渗(gene introgression)影响。其次,入侵植物中的“生态系统工程师”(ecosystem engineer)改变入侵生境水文地质状态,例如高密度的入侵芦苇(Phragmites australis)能通过降低水流速度和泥沙运输量改变河道(Kessler et al.,2011),这种被改变的生境显然无法随着入侵植物去除而恢复。第三,入侵植物能通过根系分泌物改变土壤根际微生物群落,破坏原来的微生物-植物共生关系,还能改变土壤pH值、有机质、营养成分等(全国明等,2016),即使去除入侵植物,改变的土壤理化性质和微生物环境更有利于外来植物(Magnoli et al.,2013)。最后,一个植物从地上植被中消失,它在种子库(seed bank)中的休眠种子仍使其在一段时间内存续,入侵植物能够改变种子库组成,削弱种子库对群落恢复的“保险效应”(Gioria et al.,2012)。
2.5 控制区域易发生继发入侵
入侵植物控制后往往会产生继发入侵,Pearson等(2016)对入侵植物控制进行了统计,发现超过29%控制工作促进继发入侵。当前,多物种入侵已成植物入侵的常态,这为继发入侵奠定基础,群落中入侵植物之间以及入侵植物与本地植物之间通常表现出种间竞争关系(Kuebbing et al.,2015),抑制优势物种必然会释放本地植物及其他外来植物,而外来植物又能通过种源效应、天敌释放等原因,在与本地植物竞争中获得优势(许光耀等,2018a),例如,蒙大拿州采用特异性宿主生物控制方法治理入侵乳浆大戟(Euphorbia esula),Butler等(2010)长期监测后发现,治理后控制区的优势种群仍是外来植物。入侵植物遗留效应也会促进继发入侵,本地植物遭受的遗传多样性降低、共生关系破坏等损失,短期内并不会随着目标入侵植物去除而恢复,相比本地植物,其他外来植物受到的影响显然要小得多。
3 全球变化对植物入侵管控的影响
入侵植物不单是驱动群落变化的“司机”也是搭乘全球效应而快速扩散的“旅客”(Waller et al.,2016),在气候及土地覆盖变化的条件下,入侵植物通过补偿生态损失或增加系统冗余,维持生态服务功能。
3.1 全球变化促进植物入侵,增加管控难度
生物入侵会加剧土着生物多样性丧失、生态系统崩溃及生境均质化等全球变化其他因子的进程(邓自发等,2010),另一方面人类活动正在通过富营养化、全球气候变化等迅速改变自然景观,这些变化使入侵植物不单是驱动群落变化的“司机”也是搭乘全球效应而快速扩散的“旅客”(Waller et al.,2016)。有目的的性状筛选和多次引入增大了入侵植物的物种库来源,引入优势性状的可能性更大(Buckley et al.,2016),对全球变化的响应较本地植物更快,Lee等(2016)研究发现,海平面升高、海滨生境盐度增加有助于互花米草入侵;Mozdzer等(2016)研究发现,CO2浓度上升将增加入侵芦苇的根深,并通过生态位分离促进定殖和扩散;Zhang等(2016)发现,CO2浓度升高、氮沉降能够提高入侵薇甘菊(Mikania micrantha)和飞机草(Chromolaena odorata)光合速率和光合氮利用效率。
3.2 全球变化改变人类观念,降低管控积极性
入侵生境修复涉及生态、环境、社会、经济等多个方面,全球变化可能使人类对入侵植物的观念发生改变,进而降低入侵植物管控的必要性和积极性。入侵植物一方面导致生态系统服务功能丧失(Vaz,2017),另一方面,入侵植物作为生态系统的组成部分,提高了生态系统对气候或土地覆盖变化的可塑性和适应性(Pyke et al.,2008)。作为生态系统的基础,本地植物的多样性、丰度、适合度等受全球快速变化的直接影响,并导致食物网自下而上的崩溃。在气候及土地覆盖变化的条件下,入侵植物可以通过补偿生态损失或增加系统冗余,维持碳储存、营养循环等生态功能(Mascaro et al.,2012),例如,在马约特,入侵杧果(Mangifera indica)、蒲桃属(Syzygium)物种,有助于维系本地褐狐猴(Eulemur fulvus)在破碎生境中生存(Tonnabel et al.,2011)。
4 展望
从应用层面来看,无论我们对生物入侵的研究如何深入,最终的研究成果都要面向实际应用,并成为管理行动或政府决策的依据。基于当前入侵植物管理存在的问题,今后需加强以下方面的研究:
(1)加强入侵植物生态效应研究,探寻入侵植物对标志性物种(珍惜、濒危、保护动植物等)的影响,提高政府重视和公众支持力度。在全球变化背景下,将不同尺度融入入侵植物生态效应研究,丰富人们对入侵植物–入侵生境–全球变化关系的理解,为加强入侵植物管理提供公众基础。
(2)加强入侵区域植被恢复研究。即使去除目标入侵植物,由于入侵遗留效应,本地群落重建往往受到限制,因此,应在控制试验中添加本地植物,并将其恢复评估作为入侵控制的一部分,以筛选能迅速占据空余生态位的本地物种或其组合,并确定控制方式对本地物种的负面影响。
(3)进行大时空尺度管控研究。入侵植物控制试验中,研究时间过短则无法考虑气候年度差异,会高估控制效果;研究样地大小不同,得出的结论可能存在差异甚至完全相反。实际上,考虑到地形、经济等因素,小尺度控制试验的成功技术很难应用于景观尺度(如人工除草、焚烧等),因而,应在全球变化背景下,加强大时空尺度入侵植物控制研究,为入侵植物管理提供科学依据。
(4)加强入侵植物管控决策研究。植物入侵并不限于负面影响,必须认识到入侵植物本身也是生态系统组成成分,入侵植物管理必须统筹考虑入侵物种对本地自然/社会生态系统的积极和消极影响,并将公众文化、价值观念、管控成本等纳入管理,尽可能提前发现并权衡入侵管理与生态管理、社会管理的冲突,统筹管控成本与生态损失,早期控制与长期控制,精准控制与范围控制,重点控制与全面控制等,通过成本管理筛选最佳控制方案,以尽可能地提高入侵植物管控效率。
参考文献
[]邓自发,欧阳琰,谢晓玲,等.2010.全球变化主要过程对海滨生态系统生物入侵的影响.生物多样性,6(6):605-614.
[]鞠瑞亭,李慧,石正人,等.2012.近十年中国生物入侵研究进展.生物多样性,5(20):581-611.
[]齐月,李俊生,闫冰,等.2016.化学除草剂对农田生态系统野生植物多样性的影响.生物多样性,24(2):228-236.
[]全国明,毛丹鹃,章家恩,等.2016.五爪金龙、南美蟛蜞菊入侵对土壤化学和微生物学性质的影响.植物营养与肥料学报,22(2):437-449.
[]许光耀,李洪远,孟伟庆,等.2018a.“Secondary Invasion”释义辨析及植物继发入侵.生态学杂志,37(12):3780-3786.
[]许光耀,李洪远,莫训强,等.2018b.入侵植物生态效应及其影响因素研究进展.安全与环境学报,1(18):375-380.
[]许小过,聂超仁.2014.浅析园林绿化工程建设中的腐败问题及对策.现代园艺,(7):144-145.
[]张辉,唐国强.2011.除草剂产生危害的原因及其防控措施.现代农业科技,(17):174-175.
[]周浙昆.2017.格桑花是什么.民主与科学,(3):38-41.
[]Bleeker WA,Schmitz UB,Ristow M.2007.Interspecific hybridisation between alien and native plant species in Germany and its consequences for native biodiversity.Biological Conservation,37:248-254.
[]Buckley YM,Catford J.2016.Does the biogeographic origin of species matter?Ecological effects of native and non-native species and the use of origin to guide management.Journal of Ecology,104:4-17.
[]Butler JL,Wacker SD.2010.Lack of native vegetation recovery following biological control of leafy spurge.Rangeland Ecology and Management,63:553-563.
[]Caplat P,Hui C,Maxwell BD,et al.2014.Cross-scale management strategies for optimal control of trees invading from source plantations.Biological Invasions,16:677-690.
[]Carter ET,Ravesi MJ,Eads BC,et al.2017.Invasive plant management creates ecological traps for snakes.Biological Invasions,19:443-453.
[]Catford JA,Baumgartner JB,Vesk PA,et al.2016.Disentangling the four demographic dimensions of species invasiveness.Journal of Ecology,104:1745-1758.
[]Corbin JD,D’Antonio CM.2012.Gone but not forgotten?Invasive plants’legacies on community and ecosystem properties.Invasive Plant Science and Management,5:117-124.
[]Courchamp F,Fournier A,Bellard C,et al.2017.Invasion Biology:Specific problems and possible solutions.Trends in Ecology&Evolution,1:13-22.
[]deWinton M,Edwards T,Wells R,et al.2013.Review of best management practices for aquatic vegetation control in stormwater ponds,wetlands,and lakes.[EB/OL].[2019-2-18].https://researchcommons.waikato.ac.nz/handle/10289/7999.
[]Dickens SJM,Mangla S,Preston KL,et al.2016.Embracing variability:Environmental dependence and plant community context in ecological restoration.Restoration Ecology,24:119-127.
[]Dickie IA,Bennett BM,Burrows LE,et al.2014.Conflicting values:Ecosystem services and invasive tree management.Biological Invasions,16:705-719.
[]Eviner VT,Garbach K,Baty JH,et al.2012.Measuring the effects of invasive plants on ecosystem services:Challenges and prospects.Invasive Plant Science and Management,5:125-136.
[]Gioria M,Pyek P,MoravcováL.2012.Soil seed banks in plant invasions:Promoting species invasiveness and longterm impact on plant community dynamics.Preslia,84:327-350.
[]Hester SM,Cacho OJ,Dane Panetta F,et al.2013.Economic aspects of post-border weed risk management.Diversity and Distributions,19:580-589.
[]Hirsch H,Gallien L,Impson FAC,et al.2017.Unresolved native range taxonomy complicates inferences in invasion ecology:Acacia dealbata Link as an example.Biological Invasions,19:1715-1722.
[]Hussner A,Stiers I,Verhofstad MJJM,et al.2017.Management and control methods of invasive alien freshwater aquatic plants:A review.Aquatic Botany,136:112-137.
[]Hyndman RJ,Mesgaran MB,Cousens RD.2015.Statistical issues with using herbarium data for the estimation of invasion lag-phases.Biololical Invasions,17:3371-3381.
[]Kerr DW,Hogle IB,Ort BS,et al.2016.A review of 15 years of Spartina management in the San Francisco Estuary.Biological Invasions,18:2247-2266.
[]Kessler AC,Merchant JW,Allen CR,et al.2011.Impacts of invasive plants on Sandhill Crane(Grus canadensis)roosting habitat.Invasive Plant Science and Management,4:369-377.
[]Kettenring KM,Adams CR.2011.Lessons learned from invasive plant control experiments:A systematic review and metaanalysis.Journal of Applied Ecology,48:970-979.
[]Kleunen MV,Dawson W,Essl F,et al.2015.Global exchange and accumulation of non-native plants.Nature,525:100-107.
[]Kuebbing SE,Nuez MA.2015.Negative,neutral,and positive interactions among nonnative plants:Patterns,processes,and management implications.Global Change Biology,21:926-934.
[]Lee AK,Ayres DR,Pakenham-Walsh MR,et al.2016.Responses to salinity of Spartina hybrids formed in San Francisco Bay,California(S.alterniflora×foliosa and S.densiflora×foliosa).Biological Invasions,18:2207-2219
[]Low T.2012.Australian acacias:Weeds or useful trees.Biological Invasions,14:2217-2227.
[]MacDougall AS,Turkington R.2005.Are invasive species the drivers or passengers of change in degraded ecosystems.Ecology,86:42-55.
[]Magnoli SM,Kleinhesselink AR,Cushman JH.2013.Responses to invasion and invader removal differ between native and exotic plant groups in a coastal dune.Oecologia,173:1521-1530.
[]Mahoney SM,Theimer TC,Johnson MJ,et al.2017.Similar dietary but different numerical responses to nonnative tamarisk(Tamarix spp.)by two native warblers.Biological Invasions,19:1935-1950.
[]Mascaro J,Hughes RF,Schnitzer SA.2012.Novel forests maintain ecosystem processes after the decline of native tree species.Ecological Monographs,82:221-238.
[]Mason NWH,Palmer DJ,Vetrova V,et al.2017.Accentuating the positive while eliminating the negative of alien tree invasions:A multiple ecosystem services approach to prioritising control efforts.Biological Invasions,19:1181-1195.
[]McLean P,Gallien L,Wilson JRU,et al.2017.Small urban centres as launching sites for plant invasions in natural areas:Insights from South Africa.Biological Invasions,19:3541-3555.
[]Mozdzer TJ,Langley JA,Mueller P,et al.2016.Deep rooting and global change facilitate spread of invasive grass.Biological Invasions,18:2619-2631.
[]Nackley LL,West AG,Skowno AL,et al.2017.The nebulous ecology of native invasions.Trends in Ecology&Evolution,32:814-824.
[]Neilan W,Catterall CP,Kanowski J,et al.2006.Do frugivorous birds assist rainforest succession in weed dominated oldfield regrowth of subtropical Australia.Biological Conservation,3:393-407.
[]Otega YK,Pearson DE.2011.Long-term effects of weed control with picloram along a gradient of spotted knapweed invasion.Rangeland Ecology and Management,64:67-77.
[]Pearson DE,Ortega YK,Runyon JB,et al.2016.Secondary invasion:The bane of weed management.Biological Conservation,197:8-17.
[]Proche,Wilson JRU,Richardson DM,et al.2012.Native and naturalized range size in Pinus:Relative importance of biogeography,introduction effort and species traits.Global Ecology and Biogeography,21:513-523.
[]Pyke CR,Thomas R,Porter RD,et al.2008.Current practices and future opportunities for policy on climate change and invasive species.Conservation Biology,22:585-592.
[]Pyek P,Pergl J,Essl F,et al.2017.Naturalized alien flora of the world:Species diversity,taxonomic and phylogenetic patterns,geographic distribution and global hotspots of plant invasion.Preslia,89:203-274.
[]Simberloff D.2014.Biological invasions:What’s worth fighting and what can be won.Ecological Engineering,65:112-121.
[]Smith LM,Schmitz OJ.2015.Invasive plants may promote predatormediated feedback that inhibits further invasion.Ecology and Evolution,5:2411-2419.
[]Souza-Alonso P,Gonzlez L.2017.Don’t leave me behind:Viability of vegetative propagules of the clonal invasive Carpobrotus edulis and implications for plant management.Biological Invasions,19:2171-2183.
[]Strong DR,Ayres DA.2016.Control and consequences of Spartina spp.invasions with focus upon San Francisco Bay.Biological Invasions,18:2237-2246.
[]Swope SM,Satterthwaite WH.2012.Variable effects of a generalist parasitoid on a biocontrol seed predator and its target weed.Ecological Applications,22:20-34.
[]Tassin J,Kull CA.2015.Facing the broader dimensions of biological invasions.Land Use Policy,42:165-169.
[]Tonnabel J,Tattersall I,Simmen B,et al.2011.Decline and demographic changes in the population of the near threatened brown lemur eulemur fulvus on mayotte.Oryx,45:608-614.
[]Touza J,Pérez-alonso A,Chas-amil ML,et al.2014.Explaining the rank order of invasive plants by stakeholder groups.Ecological Economics,105:330-341.
[]van Wilgen BW,Fill JM,Baard J,et al.2016.Historical costs and projected future scenarios for the management of invasive alien plants in protected areas in the Cape Floristic Region.Biological Conservation,200:168-177.
[]Vaz AS,Kueffer C,Kull CA,et al.2017.Integrating ecosystem services and disservices:Insights from plant invasions.Ecosystem Services,23:94-107.
[]Vitule JRS,Freire CA,Vazquez DP,et al.2012.Revisiting the potential conservation value of non-native species.Conservation Biology,26:1153-1155.
[]Waller DM,Mudrak EL,Amatangelo KL,et al.2016.Do associations between native and invasive plants provide signals of invasive impacts.Biological Invasions,18:3465-3480.
[]Zhang L,Chen X,Wen D.2016.Interactive effects of rising CO2and elevated nitrogen and phosphorus on nitrogen allocation in invasive weeds Mikania micrantha and Chromolaena odorata.Biological Invasions,18:1391-1407.