���﹤������

����ժ Ҫ�������Ż��̼���������ѧ�ͷ�������ѧ�����ķ��ٷ�չ,�������������ࡢ������ʽ���Ͻ������Ż�������ת��������ֲ����Ϊ���ﷴӦ��,������ת����ֲ�������ѳ�Ϊ�о��ȵ�֮һ��ת����ֲ��������Ʊ���Ҫͨ��Ŀ�꿹ԭ��ȷ��������ֲ���ѡ��ֲ���������Ĺ�����ת�������P���Ȳ�����ɡ�Ŀǰ,ת����ֲ�����簴�չ��ܿɴ����Ϊϸ�����硢�������硢���������硢�������缰���������,���ѳɹ���ֲ����½�������˿�ԭ����һϵ��������ٴ������,�ڻ����о����������Ե�����Ӧ��,�����鷶Χ�ڷֱ����˵ֿ���ԭ������μ����桢���С�Ԥ�������������á�ͨ���Ż������ӡ�ѡȡ�߱���������ֲ�������Դ����Ҷ�������ó�ת����ֲ��Ի�����ֶ�,�������ֲ�ת����ֲ������Ӧ�÷���IJ��㡣������Ҫ������ת����ֲ�����紴�ƵĻ������̡��о���չ��,����Ϊ���ת����ֲ������Ŀ�����Ӧ���ṩ�ο���˼·��
�����ؼ��ʣ�������; ת������; ��ԭ�������; ����Ӧ��;
����Abstract����With the rapid development of genetic engineering, immunology and molecular biology technologies, the types of vaccines are increasing, and the methods are being continuously established and optimized. Using transgenic plants as bioreactors to produce transgenic plant vaccines has become one of hot topics today. The preparation of transgenic plant vaccines mainly comprises of determining the target antigen, selecting the recipient plant, constructing the plant expression vector, transforming, expressing, detecting and so on. Nowadays, genetically modified plant vaccines can be broadly classified into bacterial vaccines, viral vaccines, parasitic vaccines, contraceptive vaccines and diabetes vaccines by function, the corresponding antigens have been successfully expressed in plants. After a series of biological or clinical trials, obvious immune responses were produced in the bodies, which played a role in resisting pathogenic microorganisms, preventing and controlling parasites, contraception, and preventing and treating diabetes respectively during the experiment. The shortcomings of transgenic plant vaccines will be continuously overcome by optimizing the promoter, selecting high expression receptor plants, using exogenous protein chloroplast expression, knocking out resistance genes in transgenic plant and other new methods. This paper mainly reviews the basic procedures and research progress of transgenic plant vaccines, so as to provide references and ideas for the development and application of transgenic plant vaccine in the future.
����Keyword����genetic engineering; transgenic technology; antigen gene expression; immune response
����ת����ֲ�����������÷�������ѧ����̼�������ԭ�������ͨ������ֲ��������嵼������ֲ��,����ֲ���ȫ����ʹ�������ڱ�������������Եĵ�����,�õ���ʹ�����������ԭ�ԵĻ�����������,����ͨ��ע���ʳ�ú�Ŀ�꿹ԭ��ת����ֲ�ﵰ��,��������ϵͳ��������Ӧ��,�Ӷ����������ԵĿ�������[1]��1987���38����֯����Э��(The Tissue Cutural Association)����ᵽ��ת����ֲ����о�,Curtiss��[2]��1990�����Ȼ�ú������������(Streptococcus mutants)���濹ԭA(spaA)��ת�����̲ݡ�
����ת����ֲ��������ֲ����Ϊ���ﷴӦ��,������ֲ���ڶԿ�ԭ���б���,ͬʱ�Ե�����ȷ�ļӹ�����,����ڴ�ͳ����,���������ɱ��͡��������ڶ̡��ɹ��ʸ�,���γɹ�ģ�����������ơ�����ת����ֲ��������о���������,���ѱ�����ת����ֲ����������ȡ���˺ܴ��չ����ֹĿǰ,����ת����ֲ�������Ŀ�꿹ԭ(����1)����ϸ����[����Ҷ���B�ǵ�λ(Choleratoxin B subunit,CTB)���˾������ȳ�����B�ǵ�λ(Escherichia coli heat-labile toxin B subunit,LTB)����˸˾���(Mycobacterium tuberculosis)�����������˾�ϸ��������ص���(Helicobacter pylori Cytotoxin associated protein)��]��������[�������ײ���(Hepatitis B virus)�������߲���(Foot and Mouth Disease Virus,FMDV)����״����(Rotavirus,RV)��ŵ�ֿ˲���(Norwalk Virus,NV)��]����������[��űԭ��(Plasmodium)����Ѫ��(Schistosoma)����Ƭ����(Fasciola hepatica)��]��������[������(Zona Pellucida,ZP)]��������[���ȵ���ԭ(Proinsulin,PROIN)��GAD65��ԭ��λ��],����ֲ��ӽ�Ϊ��ģʽֲ���̲ݡ����Ͻ�,����չΪ��������Խϸߵķ��ѡ�ݫ�ġ��ײ�(Brassica pekineniss)�������ȶ������ס��������ȡ�
��������������Tacket��[3]��ת��LTB���������������ٴ�����:14��־Ը����,11��ʳ��ת����������,3��ʳ�÷�ת����������,����11��־Ը������10����ѪҺ�к��п��塣���,Kapusta��[4]Ҳ�����˺��Ҹβ������濹ԭ(hepatits B surface an tigne, HBsAg)ݫ�ĵ��ٴ�����,�����ʾ,3��־Ը����2�����ڽ���2������,���ܺ������ڵĿ���ζ���߿ɴ�100 IU·L-1���������\���IJ��Ϸ�չ���Ż�,ת����ֲ��������ܿ�������������Ӧ�õ���;��,������ľ��ü����Ч�潫������ƶ��������ơ���ԭ���Ρ��������߷���ķ�չ��
����1�� ת����ֲ�����紴�ƵĻ�������
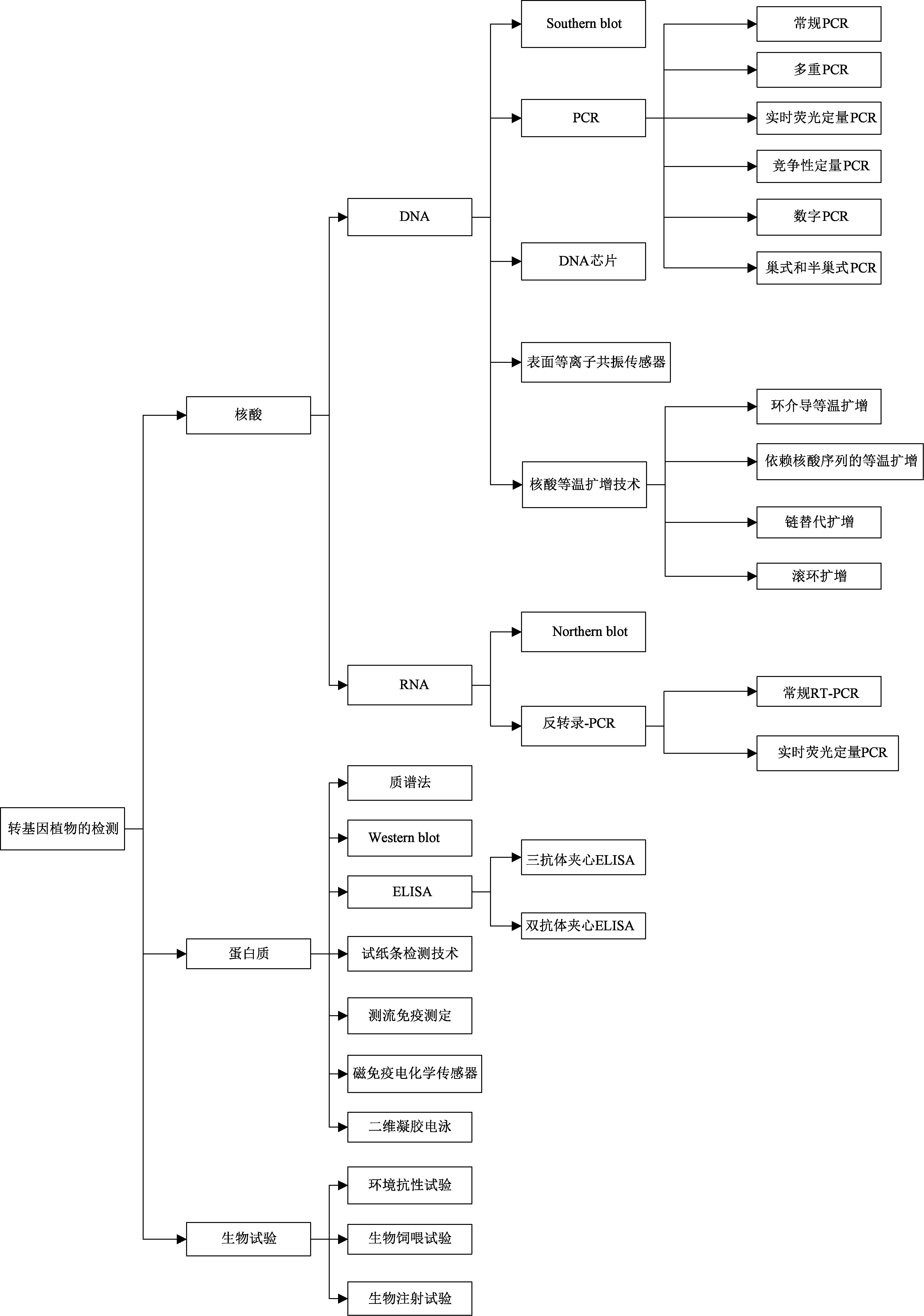
��������ת����ֲ������Ļ�ȡ;��������:1)�ȶ����Ŵ�����:ͨ����ԭ�Ľṹ����ֲ���������,�ٽ���һϵ�з�������Դ����������ֲ�������,����ܹ��ȶ��Ŵ���Ŀ��ֲ��;2)˲ʱ�Ը�Ч����:˲ʱ����ϵͳ����ԴDNA������ֲ������鲢����������,�����ڵ���ϸ��һ��ʱ�����б��ֲ���н�����ϵͳ�ķ����������Ҷ��������缤��������ǹ����������Ⱦ����ũ�˾��鵼���ȡ�����õIJ�����Ⱦ��Ϊ��(ͼ1)[5],��Ŀ�꿹ԭ����Ϊ�����ֲ�ﲡ�������ںϱ���,��Ⱦֲ���ֲ��ϸ����,ʹ֮˲ʱ���������Ա����Ŀ�군���з��뼰������ȷ�������ȷ��,������������,��ȡת����ֲ�����硣
����ͼ1 ת����ֲ������Ļ�����������[5]
![ͼ1 ת����ֲ������Ļ�����������[5]](http://www.xueshut.com/uploads/allimg/201208/36-20120Q152129C.jpg)
����Fig.1 Basic operational procedures for transgenic plant vaccines[5]
����1.1 ��Ŀ�Ŀ�ԭ��ѡ��
����ת����ֲ���������������ǵ�λ����,���н�����������ԭ�ԵĿ�ԭ�ṹ����,���������Ŵ���Ϣ��ͨ��ʹ����������,����ʹ�����յ���������Ӧ��Ӷ��ֿ���ԭ������μ����桢���С�Ԥ��������������[6]����ѡ��ԭʱ,����Ҫѡȡ��ԭ����������ԵĿ�ԭDNA,һ��Ϊ��ԭ���������ǵ��Ļ��������ױ���IJ���,��ѡȡ�������еĺ��ĵ���Ϊ��Ҫ�����Կ�ԭ��������[7]��Ŀǰ,�ѳɹ���ϸ����������������Ȳ��ֽṹ������Ϊ��Դ����������ֲ�ﲢ�õ���Ч�ر���,�������õ�����ԭ�ԡ�
����1.2�� ����ֲ���ѡ��
����ת��������ֲ���ѡ����ڿ�ԭ�������ı��������ڷ������Ӧ��ǰ������ʮ����Ҫ�����塣����ֲ��Ӧ�߱����Ŵ��������������ߡ�����������֪������������ֲ����㷺�����ڴ��漰��������˿ڷ����ص�[8]����ѡ������ֲ��Ĺ�����,Ҫ���ݸ���ֲ��������벻��,����������������Լ����߶�����ѡ����ʵ�����ֲ��̲���Ϊ�����ģʽֲ�����ﷴӦ��,������ֱ��ʳ�õ�ȱ��,�����ڴ��ģ�ƹ�����������ת����������������,�ܹ����õ�ת����ֲ������Խ��Խ��,Ŀǰ�����ڱ���ת���������ֲ���н�ˡ�â��(Mangilera indica)��ݫ�ġ����ˡ����ѡ����ܲ���ƻ������������ޣ��һϵ��ũ�����(����1)��
����1.3 �������������
��������Ĺ�����ͨ��ø�е��ֶν�Ŀ�Ļ�����ѡȡ��������������,��֤���ᴿ��ת��������̬ϸ��,�Ӷ�����ֲ�����Դ���ױ������ֲ������������ҪĿ���ǽ�Ŀ�Ļ���������θ���,ʹ��ת������ֲ�������������,�����ȶ��Ŵ������������Gateway��pBIϵ�С�pCAMBIAϵ�С�pRTLϵ�С�pPZPϵ�е�(����2),��˲ʱ����ϵͳͨ��ѡ�û�Ҭ�˻�Ҷ����(Cauliflower mosaic virus,CMV)���̲ݻ�Ҷ����(Tobacco mosaic virus,TMV)����Ϊ���塣
����1.4 ������ı�����ת��
����1.4.1�� �ȶ��Ա���ϵͳ
�����ȶ�����ϵͳ�ǽ���Դ��ԭ��������������������,��ͨ��һϵ�з���������ԭ������ֲ��ϸ����,ʹ����ֲ����������ȶ�������,��һ�������½�������Դ�����ֲ��ϸ���յ������µ�ֲ��,�������������б���Ŀ�꿹ԭ����,������״�����Ӵ�,��Ϊ���������Ʒϵ[9]��Ŀǰ�㷺���õĻ������Ϸ�ʽ������:һ�����ϵ��˻�����;�������ϵ�Ҷ��������顣�ñ���ϵͳ����ͨ��ʹ�����ӡ��龥������Ҷ��֯���������鿹ԭ���ȶ�����ϵͳ���ŵ����������ȶ������贿������ȫ�Ժ�,��ͬʱ�������Դ������б���,�ҽ���ô���ת������ϵ,��ͬʱҲ����Ŀ�군�ױ������͡�����Ч�ʵ͡���Դ������ʧ�ֲ�������ʹ������ڳ�������������ɵ�λ��ЧӦ�ȱ�[10]��
����1.4.2 ��˲ʱ�Ա���ϵͳ
����˲ʱ����ϵͳ�ǽ��������翹ԭ�����ػ������в��벡����������,����ֲ�ﲡ����ֲ��ĸ�Ⱦ,�Բ�����Ϊ���彫���鲡��������ֲ��ҶƬ��,ʹ�ÿ�ԭ�����没����ֲ�����ڽ��и��ơ�ת¼�������װ��,��ת����ֲ���ϸ���в���������鵰����[11]������ϵ������ʱ�϶̡����ײ����������ٶȿ졢�������ߡ�������Ⱦɫ���ϵ�λ��ЧӦ���ŵ�,��ȱ���ǹ������ϴ����ȶ��Ŵ��Լ���Դ������ʧ�ȡ�
����1.5�� ת����ֲ��ļ��
������������������Ҫ,ת���ɹ���ֲ��Ҫ���м��,����֤�����������翹ԭ������ʵ��,һ���Ϊ������:һ����Ե����Ŀ�꿹ԭ����ļ��;���������Դ�����������Ŀ�군�ļ��;���ǽ�����������,�������Ŀ�군���Ƿ�����������(ͼ2)��
�������ں���ļ���ΪDNA��RNA�ļ�⡣Ŀǰ���õļ��DNA�ļ����оۺ�ø��ʽ��Ӧ(polymerase chain reaction, PCR),��������PCR[12]�������Զ���PCR��ʵʱӫ�ⶨ��PCR������PCR������PCR[13]����ʽ�Ͱ볲ʽPCR[14]��,Southern blot,���������������[15],�������鵼���������������������еĵ����������������������������,�����DNAоƬ����������ӹ�����[16]�ȷ�����
�������г��õĵ����ʼ�⼼����Western blot[17]��ELISA���������߲ⶨ[18]�������ߵ绯ѧ����������ά������Ӿ[19]������������оƬ�ȡ�����֮��,��Ӧ����һϵ����������,���������������顢������ι���顢����ע�����鼰�ٴ�����ȡ�
����2 ������ֲ��������о���״
����ת����ֲ��������о�������ֲ��ɷ�Ϊ�̲ݡ����Ͻ桢���ѡ���ޣ��ƻ������������,�����ܴ��¿ɷ�Ϊ����,�ֱ�Ϊϸ�����硢�������硢�������硢��������ͼ��������硣Ŀǰ,��ʮ��ת����ֲ�������ѽ��붯������,���������õ�����ԭ�ԡ�һЩת����������ֲ������,��HbsAg��LTB��ŵ�ֿ˲�����,Ҳ�ѽ����ٴ�����,����Ԥ��������������ϸ���Ը�к,���������в�������Ӧ��,��ʾ�����õ�����Ч����
����2.1 ��ϸ������
����2.1.1�� ���ҳ�����B�ǵ�λ
����CTB���볦��ճĤϸ�����������Ե�����֬��϶�����к��������Arakawa��[20]���û�������������,������ȶ��Ŵ���ֲ��,С����ע���,�丹к�ۻ�����60%;���Kim��[21]Ҳ�ɹ����û���ͨ��ũ�˾�ת����������ֲ��;Jiang��[22]����ũ�˾��鵼����CTB����ת�뷬��ֲ��,��ýϸߴ��ȿ����Ե���,�����ԭ����ķ�����ιС��һ��ʱ���,��С���Ѫ����Ĥ�м����˻��Ҷ��ؿ��塣����CTB���ں��ܲ�[23]�ȿ�ʳֲ���гɹ�����,��Soh��[24]��Ϊˮ�����ʺ�Ӧ���ڿڷ�����,�һ���˹���������CTB�����ϸߵ�ת����ֲ��,ˮ��������Ŀ�군�ױ������ڿ������ܵ����пɴﵽ21.3 ng·μg-1��CTB���о���Խ���,ת����ϵҲ��Խ�Ϊ����,�ֶ���������ԭ�����ںϱ��
����ͼ2 ת����ֲ��ij����ⷽ��
����Fig.2 Conventional detection methods for transgenic plants
����2.1.2�� �˾������ȳ�����B�ǵ�λ
�����˾������ȳ�����(heat-labile toxin,LT)����1��A�ǵ�λ(LTA)��5��B�ǵ�λ(LTB)���,����B�ǵ�λ���������ϸ�������GM1-����֬���������Խ�ϵ�������LTB������ǿ�ҵ�����ԭ,Ҳ��һ��ǿ�������Ĥ����������LTB����������Haq��[25]��1995�����̲ݺ��������еõ�����,ͨ�����Ƿ����ߵ�С������˳�ճĤ��LTB������,����ι��С������ת�����������龥Ҳ����ڷ����ߡ�����Mason��[26]��Lauterslager��[27]Ҳ�ֱ���ũ�˾�ת������LTB������������,����С�������ճĤ���߷�Ӧ��Ŀǰ���Ե�LTB���ﵰ����������[28]��ֲ���еõ�����,��Soh��[24]��ˮ����(Glb)�����ӵĿ����»�ø��߱������Դ����,Ӧ���ڿڷ�������Ԥ����к����������ϰ��������ˮ����ʳ,�����¿��ܵ�����Դ���ױ���ʧ��,�Ӷ�Ӱ������Ч��,����߿�ֱ��ʳ��ֲ��ı���Ч�ʽ������ڸ��������������ٴ��ϵķ�չ��
����2.1.3 �����������˾�
���������ݸ˾�����һ�������Ρ�����������������Ҫ��ʮ�ֿ��̵�ϸ��,��ȷ��Ϊ������θ�ͻ����������Ҫ�²����ӡ����������˾�ϸ��������ص���(cytotoxin-associated gene A, CagA)�ѳɹ�ת���̲ݺͷ���[29]��Zhang��[30]����ũ�˾�ת�������ת����øB�ǵ�λ(urease B, UreB)���ܲ�ֲ��,ת���������ĺ��ܲ����б����UreB����Ϊ25 μg· g-1,������Ч�յ���С������߷�Ӧ,����,UreB��ԭ�������Ͻ�[31]���̲ݺ�ˮ����ֲ���н����˱����Yang��[32]ͨ������������������������������Oleosin�Ķ�Ԫ����,��ø߲�UreB����ת������ֲ��;Kalbina��[33]Ҳ��2010��ɹ�����3�ֲ�ͬ���������˾�TonB��������,����������첢����,ֲ���еı��ﵰ����Ũ��ռ�ܿ����Ե����ʵ�����0.05%��������,�����ݸ˾�����ػ�����о���Ҫ������ϸ��������ص���(CagA)������ø�����ǵ�λ(Urease)��,�������о��߶Ա��������3����Դ��������Ч�����бȽϡ����ǵ�����ת����ֲ�������δ����չ,�о���Ӧ��һ��̽���䲻ͬ��ԭ����������������ԡ�
����2.1.4�� ����ϸ������
������˷�֦�˾��������˲��IJ�ԭ��,���ַ�ȫ��������,�Էν��������Ŀǰ���н�˸˾�ESAT-6�ڷ�ľ��[13]�Լ���˸˾�MPT64�ں��ܲ�[34]�зֱ�����˱��Módolo��[35]��2018�����˽�˸˾��ǵ���APAת�������Ͻ�ֲ��;Hajibehzad��[36]��־���������˾�IpaD���̲��л��˲ʱ����,�̲�ҶƬ��ë״���о���IpaD,���̲�ë״���п������ܵ�����߱�����Ϊ0.94 ng·μg-1��Tregoning��[37]��2003�꽫���˷綾��CƬ��(tetanus C segment, TetC)ͨ��Ҷ����ת���̲�,�̲�Ҷ�����л��۵�TetCռ������ϸ���ܵ���10%,�����������TetC����ʹС�������ճĤ����Ӧ�𡣴������о��������,ϸ��������о������ڳ�����,�����о���Χ������,���ϸ��������ģʽֲ��Ϊ��,��δ�ܽ����ٴ��о�������,�պ���������ٴ�����Ŀ�չ��Ϊ�����������ϸ����ԭ���ṩ�µķ���
����2.2 ����������
����2.2.1 �������ײ�������
���������ײ���Я�������ջ����Ӳ����ΰ����������Ҹα��濹ԭ(hepatitis B sarface antigen, HBsAg)�����б���S��M��L��HBc�ȵ���ñ��Mason��[38]���罫HbsAg�����̲���,���ת����ֲ�ꡣ����ʵ��֤��,������ֲ��ҶƬ�ĵ�����ȡ��ע����������ײ�����С���к�,���������߷�Ӧ���ĸ�����HbsAg�൱�����Ҳ�ɹ�����˺��������ķ���[39]���㽶[40]�����ܲ�[41]������[42]���˲�[43]����[44]��ƻ��[45]������[46]���̲�[47]��ֲ�ꡣDeineko��[48]���ֽ�preS2-S��ԭ����ת����������ǻ���ź�������,��������ǿĿ�군����ת������ܲ��еĻ���,��Ϊת����ֲ������������ṩ���µ�˼·��Thanavala��[49]��HBsAg��ԭ����Ϊ8.5±2.1 μg·g-1��ת������������42��־Ը�߽���������,�����Ϊ3��,�����ʾ,ʳ��HBsAg��������־Ը�߾����ֳ��ζ�������ǿ��Ч��,��ʳ�ô���Խ��ζ�Խ�ߡ������ײ����ײ�������,��ʹ��������Ӧ�����,�����˸������ʼ�����������,ת����ֲ�������������ײ���������о���Ϊ�ٴ��������ɸò��������ļ����ṩ�µķ�����
����2.2.2 �������߲�������
����������(foot-and-mouth disease virus,FMDV)���ɿ����߲��������һ�ָ�Ⱦż�㶯��ļ��ԡ����ԡ��߶ȽӴ��Դ�Ⱦ������ṹ������ VP1Ϊ��Ҫ��ԭλ��,�ɲ����кͿ��塢�յ����߷�Ӧ,���ص��о�����[50]��1993��,Usha��[51]���Ƚ�VP1�������������Ҷ���� (Cowpea Mosaic Virus,CPMV)��ǵ���������,�������Ӻϲ��������Ʊ��ij�������������,����ڻ����м����嵰�ס����,���ñ������½�����������Ͻ桢��ޣ���̲ݡ������������ѵ�ֲ���в��ɹ�����,����˾�������ԭ�ԵĿ��嵰�ס�����,Dus Santos��[52]����ũ�˾�ת�����������߲���(FMDV)�¿ǵ���ǰ��P1������ø3C������ޣ,��ת����ֲ����ȡ��3������С��,10 d����С��Ѫ���м�������FMDV���塣FMDV��λ������о�һֱ���ܹ�ע,O��FMDV�ϳ����������ѱ�Ӧ������������,������ȷ�����߱���Ч������Կ�ԭ�ṹ����ϴ��A��FMDV��λ���������Ч���ǹ��ܹ�ע���о�����,��Ϊδ��A��FMDV�ķ����ṩ�µķ�����
����2.2.3�� ��״����
������״�������ں����²�����,�Dz����Ը�к����Ҫ�IJ�ԭ��о�֤ʵ,��״�������濹ԭVP7��VP4�����кͿ�ԭ����,VP6���б�����ԭ����,���нϸߵ�����ԭ�Ժͷ�Ӧԭ��[53,54]���ѱ�������״����VP7�����������ͺ��ܲ���,�������ӳ��ֲ���в����˾��л��ԵĿ�ԭ���ס�Kim��[55]����״����VP6���뷬��ϸ��,������������ϵ��,���������Ƶ��յ�����������VP6 0.73 mg·L-1;��Dong��[56]����ũ�˾�ת���������ת������״����VP6��ޣֲ��,��ޣ��VP6���ױ�����ռ�ܿ����Ե���0.28%,ǿ��С���,С��ĸ�к֢״���Լ��ᡣ��״����������Ӥ�������䶯��θ����к����Ҫ����,���о�����Ͽ�,VP6�����ܾ��и��õ�����Ч���Ҿ��и��㷺��Ӧ��ǰ����
����2.2.4 �����������²�����
������������ȱ�ݲ���(human immunodeficiency virus,abbr: HIV),����1981���״����������ֵ������������ϵͳȱ�ݵ�һ�ֲ���������һ�ָ�Ⱦ��������ϵͳϸ����������(Lentivirus),����ת¼������2002��Zhang��[57]�����˽���������ȱ�ݲ���(HIV-1)P4�������̲ݡ����Karasev��[58]���û���ǹ�����תHIV-1 Tat������,��С�����ʱ��һ�ܵ�3�οڷ�����,����С������Tat����ζ�����,���ڽ���4�ܺ�ﵽ��ֵ������Ŀǰ,HIV-1��gap��gp120�ڷ����еõ�����,Vafaee��[59]��2018����ת����ݫ���л�ýϸߵ����鵰��,�������ߴ�8.942 μg·100 mg-1��
��������ͷ������(Human papilloma virus,HPV)��һ���������ղ����Ƶ���ͷ�����ݲ���,������DNA����,�������������Ե��������ࡣ���ѳɹ���HPV-16 L1ת��������[60]���̲�[61]�ͷ���[62,63]���������绹��ŵ�ֿ˲�������[64]���᳦ֱ����(Carcinoma of colon and rectum)����[65]���ǻ�����ܰ���(Non-hodgkin’s lymphoma,NHL)����[66]�������(Measles virus,MV)����[67]������������������(Zaire ebolavirus)����[68,69]���Ǹ��Ȳ���(Dengue virus,DV)����[70]���˾�ϸ������(Human cytomegalovirus)����[71]�����۹�������[72]����Ȯ������(Rabies virus,RV)[73,74]�ȡ���Щ�²�������Ҫ������˲ʱ����,δ���������������ȶ��Ŵ����﷽��ķ�չ,���ƶ��������༲�����ٴ��ϵ�Ԥ�������ơ�
����2.2.5 �����������²�����
���������²�����ͨ������������ද����������̴�������ʧ,Ϊ��Ч��ֹ���������������Ժ��,���ѳɹ����һ��ת����ֲ�����硣Tien��[75]���������Ը�к����(Porcine Epidemic Diarrhea Virus, PEDV)S1D��ԭͨ��Ҷ����ת���������̲�,ֲ�������S1D������������ԼΪ������0.07%,�ںϵ���S1D-CTB����ˮƽ�ﵽ1.4%;An��[76]����˱�������ֳ�ͺ����ۺ�������(Porcine Reproductive and Respiratory Syndrome Virus,PRRSV)GP4D��GP5D�������Ͻ�ֲ��,��ת�������Ͻ��л��1.66%Ŀ�군��,������������������ڼ������Կ��塣2015��,Mbewana��[77]���̲���˲ʱ�����˼���H5NI�����в���M2e����,����ֲ����������鵰�״�����ע�䵽С����,Westernӡ������֤ʵ�����ߵ�С��Ѫ���д���M2e�����Կ��塣���������������ϢϢ���,�������ද��IJ�ԭ�������й�ע����ʱ��ȡԤ����ʩ���������ڱ�������,����������ʧ,���ɼ�ӵض��˵�����������������Ӱ�졣
����2.3�� ����������
����ű���������Ե����ؼ����没�������没����Σ����ֳҵ,������ʵ����,��ѧҩ��Ĵ���ʹ��,��������������Ѳ�����ҩ��,��ͳ�Ļ�ѧҩ����η����ܵ���������ս,��ת����ֲ�����翪������Ч���Ƽ����没��;����Turpen��[78]������űԭ�濹ԭ�����ػ�������̲ݻ�Ҷ����,�Թ���ֲ�ﲡ������,Ȼ��תȾ�̲�, ��ҶƬ�л�ȡ��0.4��1.2 mg·g-1 ʪ�����鵰�ס���ϼ��[79]��űԭ����λ��ԭ������CTB�����ںϹ���ֲ���������,���ת�����̲�,��PCR����֤ʵ���ϳɹ���Ŀǰ,���й��γ�SAG1����[80]����Ƭ������Ҫ�����Կ�ԭ����FH3���ձ�Ѫ����������[81]����ת�뵽���ѡ��ϻ���ޣ���Ͳ�ֲ����,���ɹ�����˱��ԭ��ת��ֲ�ꡣ
����2.4 ����������
�������鶯��������������ĸϸ��������ϸ�����ڲ���������ĸϸ�����Ŵ�ǰ�ܾ������һ�����,ZP�����ھ��ѽ�ϼ����Ӵ������������ŷdz���Ҫ������,Ӱ���ܾ����̵ijɰܡ�Fitchen��[82]��С��ZP3���Ŀ�ԭ�����ز���TMV���¿ǵ�����,תȾֲ�ﲢ��ø�ˮƽ���ںϵ���,����Щ��ȡ������������Ϊ��ԭ���г�θ������,ʹ��С��Ѫ�忹��ʶ�������ڵ�ZP3��λ,�ҷ��������ۼ��п�ZP3����,�Ӷ��ﵽ�˱��е�Ч��������Ŀǰ,ת�����������ı�����Խ���,��ת����������翪���ɹ�,����ر��������Ⱥ��
����2.5�� ��������
�����������������һ����������������������,Ŀǰ��Ҫͨ��ע���ȵ��������Ƹ�Ѫ�ǡ�����[83]���ȵ���ԭ���ȵ���B��(insulin B chain,INSB)����������,���ת����ֲ��;��������[84]���û���ǹ��,���������������һ��������ԭ—��P277��CTB�ںϻ���ת��С��,���ת�����������硣Ruhlman��[85]ͨ��Ҷ����ת������CTB�����ȵ���ԭ(Pins)�����̲�,�����Զ��յ��ȵ���ʣ�������βϸ�����,CTB-Pins������С����ȵ��в����ȵ��ص�βϸ����������,���нϵ͵�ѪҺ����Һ������ˮƽ;Kwon��[86]���̲���ʹCTB�밬������(exenatide)�����ںϱ���,�����ˮƽ�ﵽ��Ҷ����14.3%; Choi��[87]ʹ��˲ʱת����,����ͨ�̲�ҶƬ�м���������-4����ת�������ںϵ��ײ����ߴ�37 μg·g-1,�����ڱ���ķ���̲ݵ�˲ʱת��Ҷ�л��137 μg·g-1�ı�������������Ϊ�����һ�ִ�л�ϰ�����,����������Ч�ؿ���,��Ϊ�������һϵ��������Ӱ��,�����������ڸ����ʳ��ֲ�﷽�������,��Ϊ�����������˵������ṩ�µ��ֶ�,���������������о�����̱�ʽ�����塣
����3 ���ܽ���չ��
��������ڴ�ͳ����ת����ֲ�������м��������,��������һЩ����:1)��ԭ����Ч�ʵ�:��Ҳ��Ŀǰת����ֲ�����������ٵ��������֮һ,��Դ���ױ��ﺬ����,��ʹ�ù�������ʧȥ�������,�������俪����Ӧ�á�2)���ﰲȫ��:ת����ֲ��Ļ�������[11],һ����ת����ֲ��Ŀ���ɸѡ���ܶ�Ӧ�û����������Ԥ���Σ��,��һ���������д��ģ����ֲҲ���ܻ����Χ�����������������һ��Ӱ�졣3)����Ч����:ת����ֲ����Ϊһ�ֿڷ�����ʹ��ʱ,����Դ���ᾭ����ϵͳ����ͬ�̶ȵؽ���,�Ӷ����Ϳ�ԭ��[88]��4)������:����������������������������߳���ԭ�����Եȶ������ۺ����õĽ��,������Ը������������������,�ᵼ������Ч����
���������������,Ŀǰ����ת����ֲ��������Ż��ɲ�ȡ���´�ʩ:1)��ͬ���������ڲ�ͬ��ת����ֲ���б���Ч�ʲ����,�Ҳ�ֲͬ���Ŀ�꿹ԭ�ı�����Ҳ��ͬ,���,��ȡ�Ż������ӻ���ѡ����ֲ��ķ������Ŀ�군�ı�����������,����ԭDNA������Ҷ����������˻�����Ҳ��������߱���Ч�ʡ�2)����Ҷ�����ĸϵ�Ŵ�ʹ��Դ���װ�����Ҷ����Ҳ���Խ����������IJ���Ӱ�졣����,Ҳ�������������Ի������������ӵĿ�����,ʹ�ÿ������ƻ����Բ�������ֲ������û��̼�����ת����ֲ��Ŀ��Ի�������ó���3)����ʹ��ɳ���Ͼ��ͻ�״���Ҹ˾���Ϊ�������[89]�����ð��¶Կڷ�������б��������Ӷ���������������ڵ��ȶ���,�������Ч����4)�ɼ��ٷ��û�ʹ�ô���,�Ӵ��������ȷ�ʽ�Ա������������⡣
������������,��ֲ���������ṩ������۵ĵ�����Դ,������ת����ֲ�↑������������һ�ֽϾ�����Ч��;��,�Ҿ��й����ķ�չǰ������������ֲ��ѧ������ѧ����������ѧ�ȸ���ѧ�������Ѹ�ٷ�չ,��ת����ֲ��ı������������ֲ�������ԭ�Ժ��ٴ����۵������о�,ת����ֲ�������ܹ������Ż�,�õ��㷺Ӧ�ò��츣���ࡣ
��������1 Ŀ�꿹ԭ������ֲ�����ϵͳ���ձ�
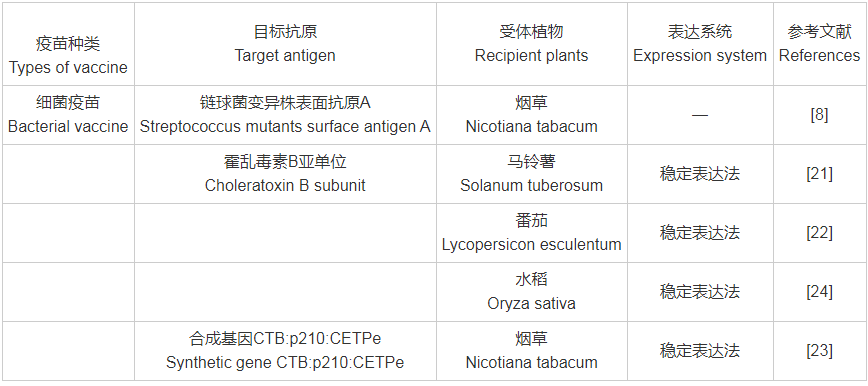
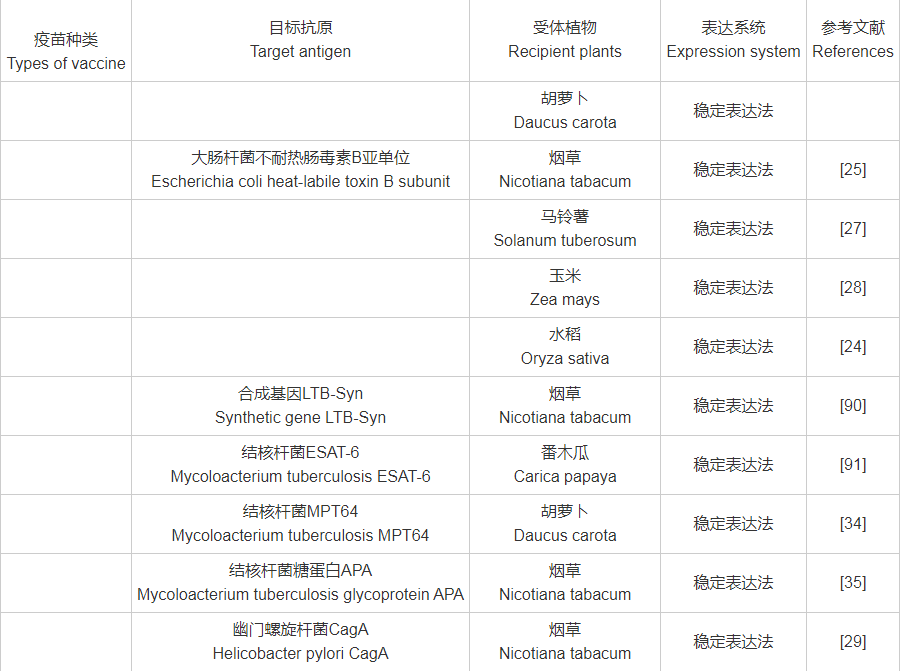
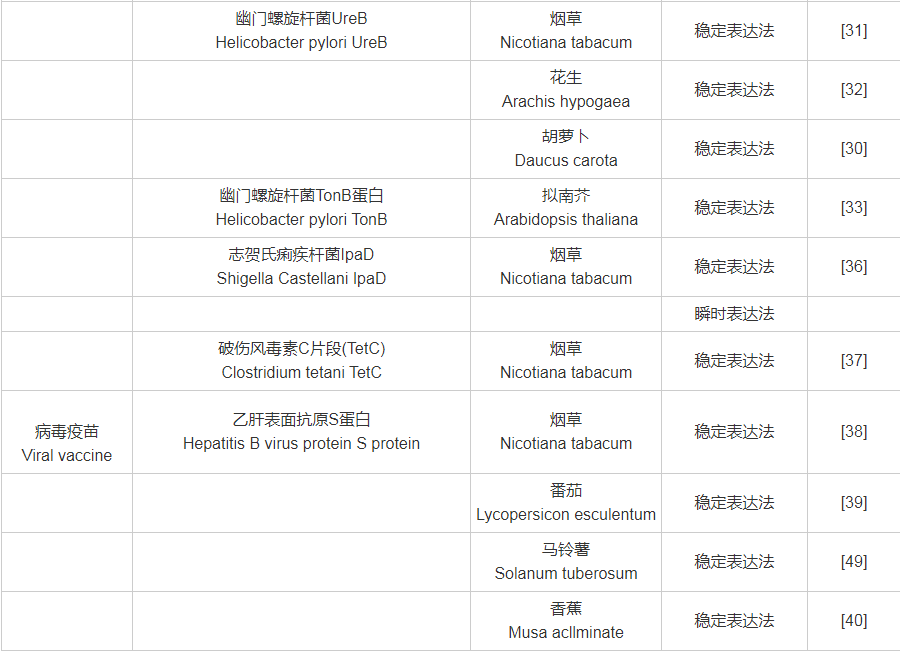
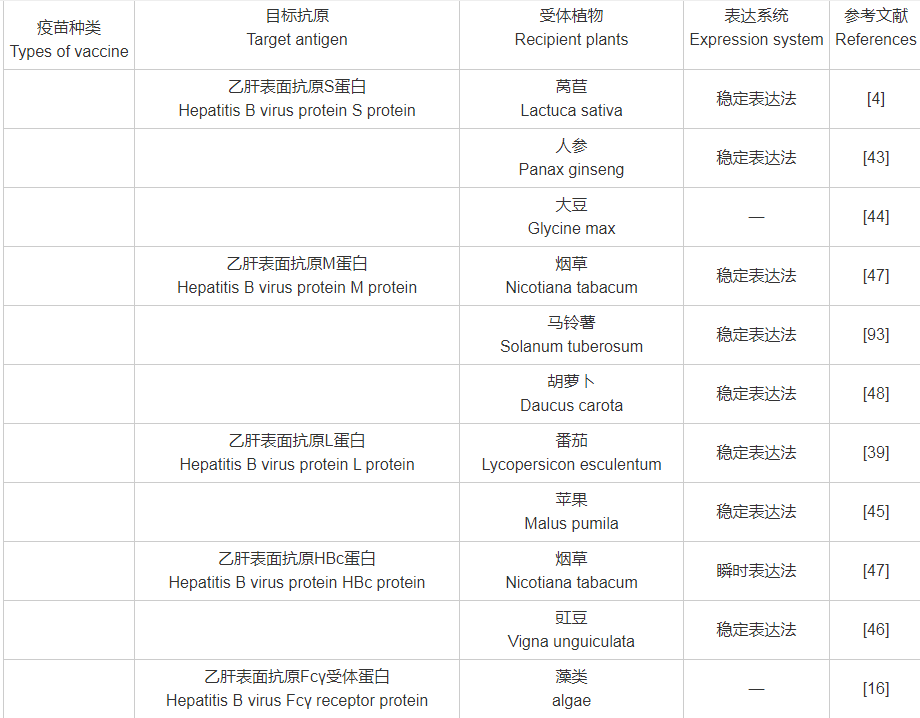
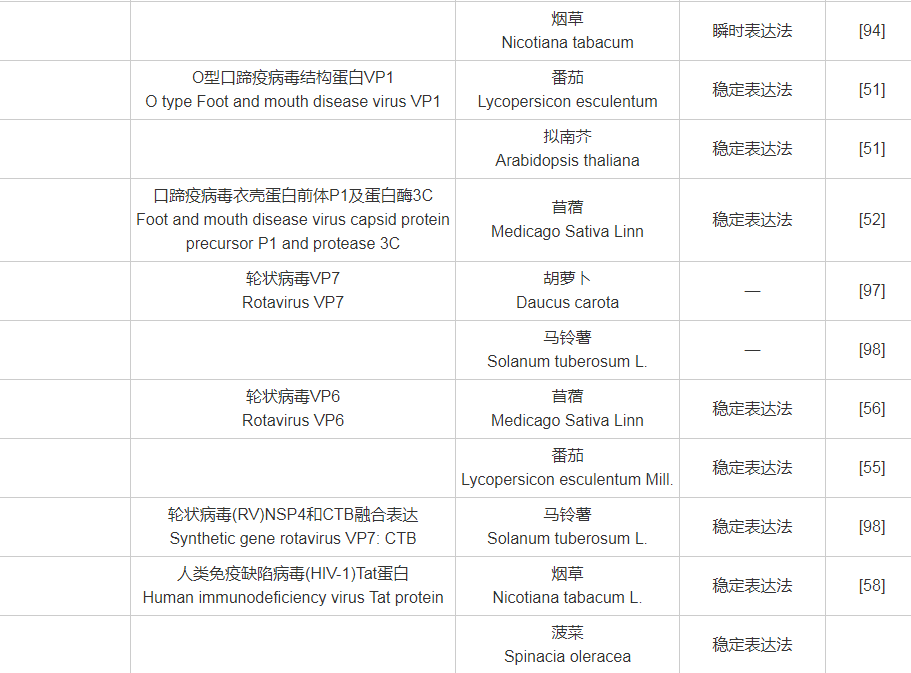
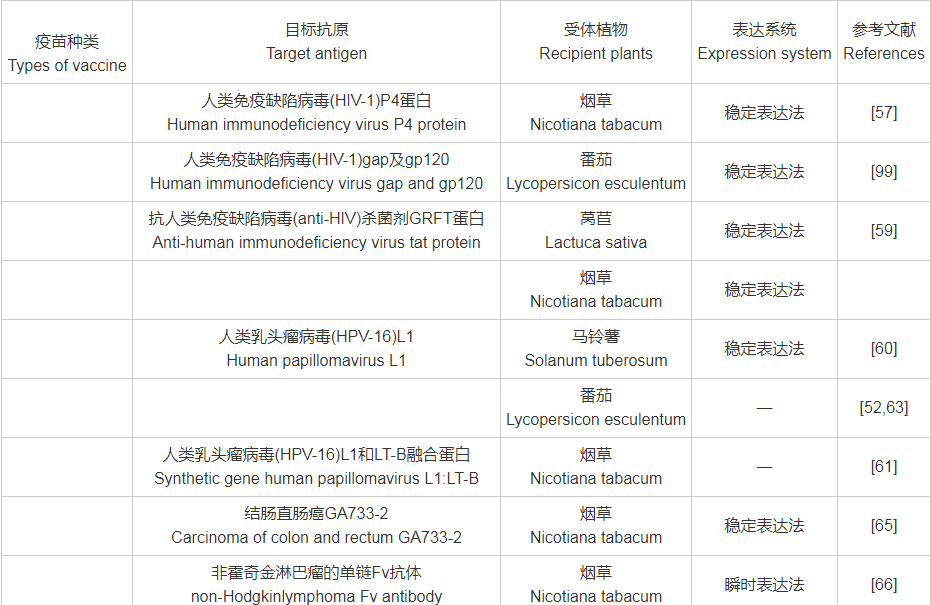
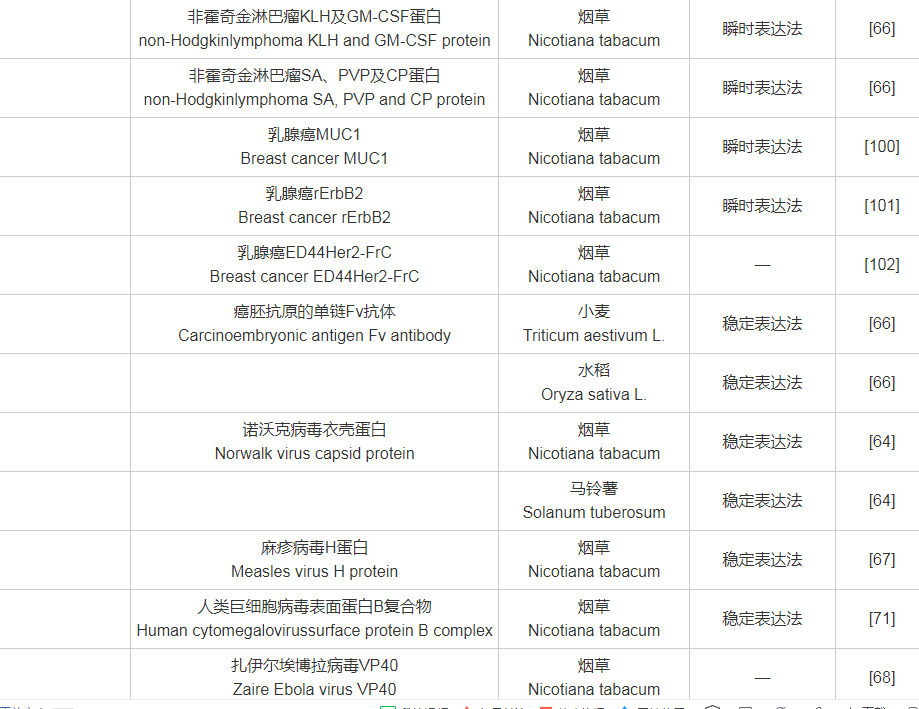
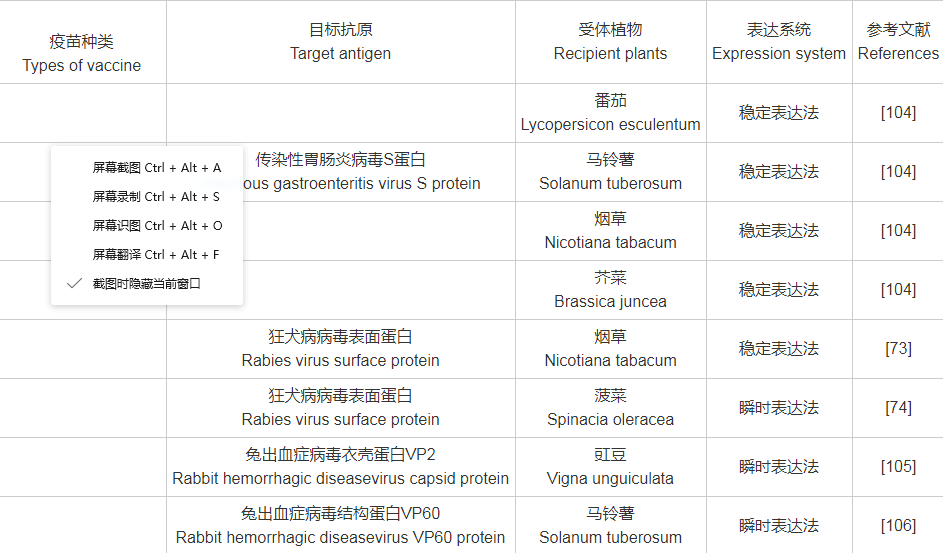
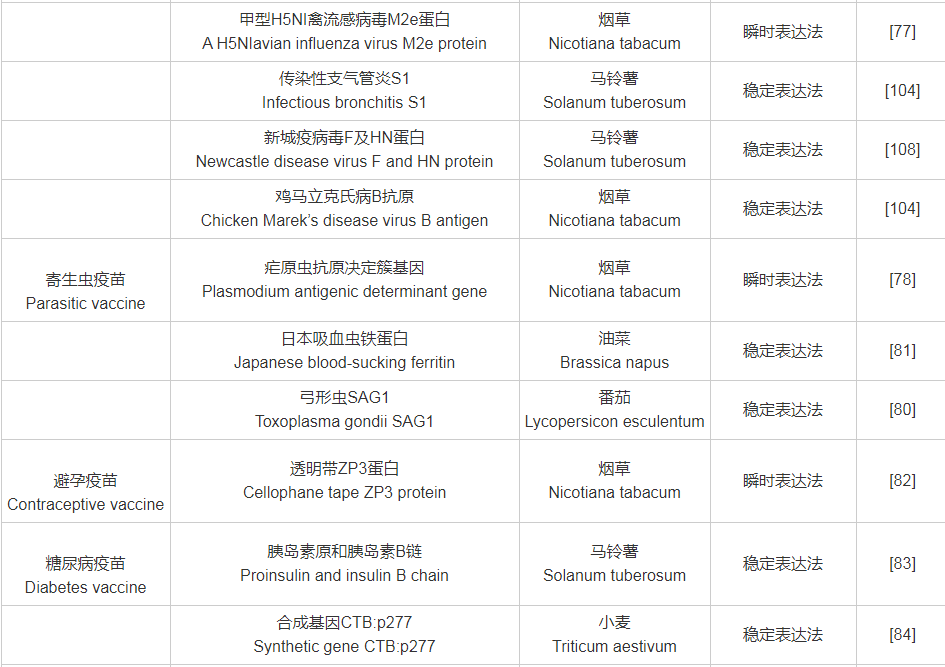
��������1(��)
����2 ���곣��ֲ���������������ֲ��һ����
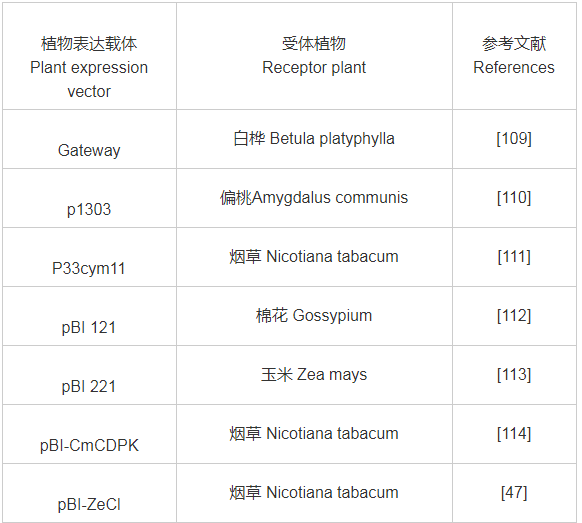

���������
����[1]Mason H S,Warzecha H,Mor T,Arntzen C J.Edible plant vaccines:Applications for prophylactic and therapeutic molecular medicine[J].Trends Molecular Med,2002,8(7):324-329
����[2]Curtiss R I,Cardineau C A.Oral immunization by transgenic plants[J].World Plant Application,1990,90:107-111
����[3]Tacket C O,Mason H S,Losonsky G,Clements J D,Levine M M,Arntzen C J.Immunogenicity in humans of a recombinant bacterial antigen delivered in a transgenic potato[J].Nature Medicine,1998,4(5):607-609
����[4]Kapusta J,Modelska A,Figlerowicz M.A plant-derived edible vaccine against hepatitis B.virus[J].The FASEB Journal,1999,13(13):1796-1799
����[5]Mason H S,Arntzen C J.Transgenic plants as vaccine production systems[J].Trends in Biotechnology,1995,13(9):388-392
����[6]�����־�£�����ʡ��ת����ֲ�������о���չ[J]�����\��ͨ����2010(4):28-32
����[7]Tae-Geum K,Moon-Sik Y.Current trends inedible vaccine development using transgenic plants[J].Biotechnology and Bioprocess Engineering,2012,15(1):61-65
����[8]��ΰ���������������ۣ��Ӿ���ת����ֲ��ڷ������о���չ[J]������ҽѧ��չ��2005,26(6):6-8
����[9]��������ת����ֲ�������о���״[J]�����ɹ���ҽҩ��2014,33(5):117-119
����[10]Wong-Arce A,González-Ortega O,Rosales-Mendoza R.Plant-made vaccines in the fight against cancer[J].Trends in Biotechnology,2017,35(3):241-256
����[11]Kumar G,Karthik L,Rao K V B.Plant vaccines:An overview[J].Microbial Bioprospecting for Sustainable Development,2018,35(3):249-263
����[12]κ�������ƣ�����ϼ��������ë����������Դ�����ȶ��Լ������������Ӱ��[J]����ũѧ����2009,23(6):1054-1059
����[13]Fu W,Zhu P Y,Wang C G,Huang K L,Du Z X,Tian W Y,Wang Q,Wang H Y,Xu W T,Zhu S F.A highly sensitive and specific method for the screening detection of genetically modified organisms based on based on digital PCR without pretreatment[J].Scientific Reports,2015,5(1):1-10
����[14] Huang Z,Dry I,Webster D,Strugnell R,Wesselingh S.Plantderived measles virus hemagglutinin protein induces neutralizing antibodies in mice[J].Vaccine,2001,19(15/16):2163-2171
����[15]Ҷ�Z����ۣ�����������������ⷽ�����о���չ[J]���Ƽ�ͨ����2019,35(3):215-218
����[16]Vanier G,Stelter S,Vanier J,Hempel F,Maier U G,Lerouge P,Ma J L,Bardor M.Alga-made anti-Hepatitis B antibody binds to human Fcγreceptors[J].Biotechnology Journal,2017,13(4):e1700496
����[17]�ǵ�ϼ��תBt˫�ۻ�������Ŀ����Լ��Ŵ��ȶ����о�[D]������:�й�ũҵ��ѧ��2014:23
����[18]Holst-Jensen A,Ronning S B,Lovseth A,Berdal K G.PCRtechnology for screening and quantification of genetically modified organisms (GMOs)[J].Analytical and Bioanalytical Chemistry,2003,375(8):985-993
����[19]�ų�������������ܣ�Ѧ�£����� ̄���������ξ�ǻԣ�תHBs Ag����ӣ�ҷ���������ҶƬ�ܵ���˫���Ӿ��ϵ�Ľ���[J]����ũѧ����2012,26(8):1111-1117
����[20]Arakawa T,Chong D K X,Langridge W H R.Efficacy of a food plant based oral choleratoxin B subunit vaceine[J].Nature Biotechnology,1998(16):292-297
����[21]Kim T G,Gruber A,Langridge W H R.HIV-1 gp120 V3 cholera toxin B subunit fusion gene expression in transgenic potato[J].Protein Expression and Purification,2004,37:196-20
����[22]Jiang X L,He Z M,Peng Z Q,Qi Y,Chen Q,Yu S Y.Choleratoxin B protein in transgenic tomato fruit induces systemic immune response in mice[J].Transgenic Research,2007,16(2):169-175
����[23]Govea-Alonso D O,Tello-Olea M A,Beltrán-López J,MonrealEscalante E,Salazar-Gonzalez J A,Banuelos Hernandez B,RosalesMendoza S.Assessment of carrot callus as biofactories of an atherosclerosis oral Vaccine prototype[J].Molecular Biotechnology,2017,59(11/12):482-489
����[24]Soh H S,Chung H Y,Lee H H,Ajjappala H,Jang K,Park J H,Sim J S,Lee G Y,Lee H J,Han Y H,Lim J W,Choi I,Chung I S,Hahn B S.Expression and functional validation of heat-labile enterotoxin B (LTB) and cholera toxin B (CTB) subunits in transgenic rice (Oryza sativa)[J].Springer Plus,2015,4 (148):148-162
����[25]Haq T A,Masson H S,Clements J D,Arntzen C.Immunization with a recombinant bacrerial antigen produced in transgenic plants[J].Science,1995,268(5211):714-716
����[26]Mason H S,Haq T A,Clements J D,Arntzen C J.Edible vaccine protects mice against Escherichia coil heat-labile enterotoxin (LT):Potatoes espressing asynthetid LT-B gene[J].Vaccine,1998,16(13):1336-1343
����[27]Lauterslager T G M,Florack D E A,van der Wal T J,Molthoff J W,Langeveld J P M,Bosch D,Boersma W J A,Hilgers L A T.Oral immunization of naive and primed animals with transgenic potato tubers espressing LT-B[J].Vaccine,2001,19(17/19):3661-3670
����[28]Chikwamba R K,Cunnick J,Hathaway D,Mc Murray J,Mason H,Wang K.A functional antigen in a pratical crop:LT-B producing maize protests mice against Escherichia coli heat labile enterotoxin(LT) and cholera toxin(CT)[J].Transgenic Research,2002,11(5):479-493
����[29]����ˮ����ƽ��������Ƽ����ϣ������ݸ˾�Cag A������CTB�ںϻ���Ŀ�¡����ֲ���������Ĺ���[J]���Ƽ�ͨ����2004,20(3):189-193
����[30]Zhang H X,Liu M,Li Y J,Zhao Y H,He H,Yang G D,Zheng CC.Oral immunogenicity and protective efficacy in mice of a carrot derived vaccine candidate expressing Ure B subunit against Helicobacter pylori[J].Protein Express and Purification,2010,69(2):127-131
����[31]Gu Q,Han N,Liu J Y,Zhu M Y.Expression of Helicobacter pylori urease subunit B gene in transgenic rice[J].Biotechnology Letters,2006,28(20):1661-1666
����[32]Yang C Y,Chen S Y,Duan G C.Transgenic peanut (Arachis hypogaea L.) expressing the urease subunit B gene of Helicobacter pylori[J].Current Microbiology,2011,63(4):387-391
����[33]Kalbina I,Engstrand L,Andersson S,Strid A.Expression of Helicobacter pylori Ton B protein in transgenic Arabidopsis thaliana:Toward production of vaccine antigens in plants[J].Helicobacter,2010,15(5):430-437
����[34]���轨���ߵϰ�����������������������ת������ܲ�����ν������[J]��ֲ��ѧ����2001,43(2):132-137
����[35]Módolo D G,Horn C S,Soares J S M.,Yunes J A,Lima L M,de Sousa S M,Menossi M.Transgenic Nicotiana tabacum seeds expressing the Mycobacterium tuberculosis Alanine-and Proline-rich antigen[J].AMB Express,2018,12(8):178-188
����[36]Hajibehzad S S,Mehrizi F A,Honari H,Alizadeh H.Hijacking tobacco hairy roots and leaves in order to produce Ipa D antigen by means of different signal peptides[J].Journal of Crop Science and Biotechnology,2017,20(5):359-368
����[37]Tregoning J S,Nixon P,Kuroda H,Svab Z,Clare S,Bowe F,Fairweather N,Ytterberg J,van Wijk K J,Dougan G,Maliga P.Expression of tetanus toxin Fragment C in tobacco chloroplasts[J].Nucleic Acids Research,2003,31(4):1174-1179
����[38]Mason H S,Lam M K,Arntzen C J.Expression of Hepatitis B surface antigen in transgenic plants[J].Proceedings of the National Academy of Sciences of the United States of America,1992,89 (24):11745-11749
����[39]Salyaev R K,Rekoslavskaya N I,Stolbikov A S,Hammond R W,Shchelkunov S N.Synthesis of hepatitis B virus surface antigen in tomato plants transgenic for the pre S2-S gene[J].Doklady Biochemistry and Biophysics,2007,416(1):290-293
����[40]Sunil-Kumar G B,Ganapathi T R,Revathi C J,Srinivas L,Bopat VA.Expression of hepatitis B surface antigen in transgenic banana plants[J].Planta,2005,222(3):484-493
����[41]����Ӣ�����������������Ƶ�ׯ���ֻ�ƽ����ϼ�����ƽ���Ҹβ������濹ԭ�����ں��ܲ��еĿ�¡������[J]������ѧ����ѧ��չ��2002,30(1):1-4
����[42]�º��ң��ž������㣬�ź�������Ӣ��֣���������ۣ��Ҹβ������濹ԭ�����ڻ����е��Ŵ�ת��������ԭ�Լ��[J]�����\��ͨѶ��2002,13(4):245-250
����[43]�����ף�ʢ�����������������꣬�����������������������������ײ������濹ԭ�������˲�ϸ���еĸ�Ч����[J]���й�������Ʒѧ��־��2008,21(2):119-122,130
����[44]Smith M L,Keegan M E,Mason H S,Shuler M L.Factors important in the extraction,stability and in vitro assembly of the hepatitis Bsurface antigen derived from recombinant plant systems[J].Biotechnology Progress,2002,18(3):538-550
����[45]¦����������ҦȪ�飬�ܰ����������������պɣ����ͣ��Ҹα��濹ԭ��(S1S2S)������ת����ƻ���б���[J]������ѧ����2005,22(6):5-9
����[46]Mechtcheriakova I A,Eldarov M A,Nicholson L,Shanks M,Skryabin K G,Lomonossoff G P.The use of viral vectors to produce hepatitis B virus core particles in plants[J].Journal of Virology Methods,2006,131(1):10-15
����[47]Huang Z,Santi L,Le Pore K,Kilbourne J,Arntzen C J,Mason H S.Rapid,high-level production of hepatitis B core antigen in plant leaf and its immunogenicity in mice[J].Vaccine,2006,24(14):2506-2513
����[48]Deineko E V,Zagorskaya A A,Pozdnyakov S G,Filipenko E A,Permyakova N V,Sidorchuk Y V,Uvarovac E A,Pozdnyakovac L D,Shumnya V K,Vlasovb V V,Hammond R V,Shchelkunov S N.Comparative analysis of HBV M-antigen production in leaves of inpidual transgenic carrot plants[J].Doklady Biochemistry and Biophysics,2009,425(1):76-79
����[49]Thanavala Y,Mahoney M,Pal S,Scott A,Richter L,Natarajan N,Goodwin P,Arntzen C J,Mason H S.Immunogenicity in humans of an edible vaccine for Hepatitis B[J].Proceedings of the National Academy of Sciences of the United States of America,2005,102(9):3378-3382
����[50]��ޱ�����ڣ����㣬����������ΰ���½�ϲ�������������壮O�Ϳ����߲����ṹ������VP1�Ŀ�¡��ԭ�˱���[J]������ũҵ��ѧ��2008,36(35):15386-15388
����[51]Usha R,Rohll J B,Spall V E,Shanks M,Maule A J,Johnson J E,Lomonossoff G P.Expression of an animal virus antigenic site on the surface of a plant virus particle[J].Virology,1993,197(1):366-374
����[52]Dus Santos M J,Carrillo C,Ardila F,R�cos R D,Franzone P,Piccone M E,Wigdorovitz A,Borca M V.Development of transgenic alfalfa plants containing the foot and mouth disease virus structural polyprotein gene pland its utilization as an experimental immunogen[J].Vaccine,2005,23(15):1838-1843
����[53]���棬�����Jing H,Glass R I,Gentsch J R�����緶Χ��A����״����G1�Ͷ���VP7����VP4����߱����ķ���[J]���л�ʵ����ٴ�����ѧ��־��1997(1):3-7
����[54]��˾������ƽ���ź�����Ӧ�ۻۣ���������ҫ�ޣ�A����״�����ṹ����vp6�ڼҲϸ�״��������ϵͳ�еı���[J]����ҵ��ѧ��2012,38(6):1024-1028
����[55]Kim C H,Kim K I,Hong S H,Lee Y H,Chung I S.Improved production of recombinant rotavirus VP6 in sodium butyrate supplemented suspension culture of transgenic tomato (Lycopersion esculentum Mill.) cells[J].Biotechnology Letters,2001,7 (13):1061-1066
����[56]Dong J L,Liang B G,Jin Y S,Zhang W J,Wang T.Oral immunization with p Bs VP6-transgenic alfalfa protects mice against rotavirus infection[J].Virology,2005,339(2):153-163
����[57]Zhang G G,Rodrigues L,Rovinski B,White K A.Production of HIV-1 p24 protein in transgenic tobacco plants[J].Molecular Biotechnology,2002,20(2):131-136
����[58]Karasev A V,Foulke S,Wellens C,Rich A,Shon K J,Zwierzynski I,Hone D,Koprowski H,Reitz M.Plant based HIV-1 vaccine candidate:Tat protein produced in spinach[J].Vaccine,2005,23(15):1875-1880
����[59]Vafaee Y,Alizadeh H.Heterologous production of recombinant antiHIV microbicide griffithsin in transgenic lettuce and tobacco lines[J].Plant Cell Tissue and Organ Culture,2018,135(1):85-97
����[60]Warzecha H,Mason H S,Lane C,Tryggvesson A,Rybicki E,Williamson A L,Clements J D,Rose R C.Oral immunogenicity of human papillomavirus-like particles expressed in potato[J].Journal of Virology,2003,77(16):8702-8711
����[61]Liu H L,Li X K,Lei T,Li W S,Lusheng S,Jin Z.Transgenic tobacco expressed HPV16-L1 and LT-B combined immunization induces strong mucosal and systemic immune responses in mice[J].Human Vaccine Immunotherapeutics,2013,9(1):83-89
����[62]Monroy-García A,Go’mez-Lim M A,Weiss-Steider B,Herna’ndezMontes J,Huerta-Yepez S,Rangel-Santiago J F,Santiago-Osorio E,Mora Garc�c’a M de L.Immunization with an HPV-16 L1-based chimeric virus-like particle containing HPV-16 E6 and E7 epitopes elicits long-lasting prophylactic and therapeutic efficacy in an HPV-16 tumor mice model[J].Archives of Virology,2014,159(2):291-305
����[63]Salyaey R K,Rekoslavskaya N I,Stolbikov A S.The new plant expression system for the development of vaccines against papillomaviruses[J].Dokl Biochem Biophys,2019,484(1):52-54
����[64]Cortes-Penfield N W,Ramani S,Estes M K,Atmar R L.Prospects and challenges in the development of a norovirus vaccine[J].Clinical Therapeutics,2017,39(8):1537-1549
����[65]Lu Z,Lee K J,Shao Y,Lee J H,So Y K,Choo Y K,Oh D B,Hwang K A,Oh S H,Han Y S,Ko K.Expression of GA733-Fc fusion protein as a vaccine candidate for colorectal cancer in transgenic plants[J].Journal of Biomedicine and Biotechnology,2012,2012:1-11
����[66]Jobsri J,Allen A,Rajagopal D,Shipton M,Shipton M,Kanyuka K,Lomonossoff G P,Ottensmeier C,Diebold S S,Stevenson F K,Savelyeva N.Plant virus particles carrying tumour antigen activate TLR7 and induce high levels of protective antibody[J].PLo S One,2015,10(2):e0118096
����[67]Huang Z,Dry I,Webster D.Plant-derived measles virus hemagglutinin protein induces neutralizing antibodies in mice[J].Vaccine,2001,19(15/16):2163-2171
����[68]Monreal-Escalante E,Ramos-Vega A A,Salazar-Gonza’lez J A,Banuelos-Herna’ndez B,Angulo C,Rosales-Mendoza S.Expression of the VP40 antigen from the Zaire ebolavirus in tobacco plants[J].Planta,2017,246(1):123-132
����[69]Ríos-Huerta R,Monreal-Escalante E,Govea-Alonso D O,Angulo C,Rosales-Mendoza S.Expression of an immunogenic LTB-based chimeric protein targeting Zaire ebolavirus epitopes from GP1 in plant cells[J].Plant Cell Report,2017,36(2):355-365
����[70]Gottschamel J,Lossl A,Ruf S,Wang Y L,Skaugen M,Bock R,Clarke J L.Production of dengue virus envelope protein domain��-based antigens in tobacco chloroplasts using inducible and constitutive expression systems[J].Plant Molecular Biology,2016,91 (4/5):497-512
����[71]Tackaberry E S,Dudani A K,Prior F,Tocchi M,Sardana R,Altosaar I,Ganz P R.Development of biopharmaceuticals in plant expression systems:Cloning,expression and immunological reactivity of human cytomegalovirus glycoprotein B (UL55) in seeds of transgenic tobacco[J].Vaccine,1999,17(23/24):3020-3029
����[72]Entesari M,Wakasa Y,Zanjani B M,Takaiwa F.Change in subcellular localization of overexpressed vaccine peptide in rice endosperm cell that is caused by suppression of endogenous seed storage proteins[J].Plant Cell Tissue and Organ Culture,2018,133(2):275-287
����[73]Tiwari S,Mishra D K,Roy S,Singh A,Singh P K,Tuli R.High level expression of a functionally active cholera toxin B:Rabies glycoprotein fusion protein in tobacco seeds[J].Plant Cell Reports,2009,28(12):1827-1836
����[74]Hu R,Zhang S,Fooks A R,Yuan H J,Liu Y,Li H T,Tu C C,Xia X Z,Xiao Y Q.Prevention of rabies virus infection in dogs by a recombinant canine adenovirus type-2 encoding the rabies virus glycoprotein[J].Microbes Infection,2006,8(4):1090-1097
����[75]Tien N Q D,Huy N X,Kim M Y.Improved expression of porcine epidemic diarrhea antigen by fusion with cholera toxin B subunit and chloroplast transformation in Nicotiana tabacum[J].Plant Cell Tissue and Organ Culture,2019,137(2):213-223
����[76]An C H,Nazki S,Park S C,Jeong Y J,Lee J H,Park S J,Khatun A,Kim W,Park Y,Jeong J C,Kim C Y.Plant synthetic GP4 and GP5 proteins from porcine reproductive and respiratory syndrome virus elicit immune responses in pigs[J].Planta,2018,247 (4):973-985
����[77]Mbewana S,Mortimer E,Pêra F F P G,Hitzeroth I I,Rybicki E P.Production of h5n1 influenza virus matrix protein 2 ectodomain protein bodies in tobacco plants and in insect cells as a candidate universal influenza vaccine[J].Frontiers in Bioengineering and Biotechnology,2015,12(3):1-8
����[78]Turpen T H,Reinl S J,Charoenvit Y,Hoffman S L,Fallarme V,Grill L K.Malarial epitopes expressed on the surface of recombinant tobacco mosaic virus[J].Nature Biotechnology,1995,13 (1):53-57
����[79]��ϼ���º������������ӻԣ��ܳϣ��������ű���ԭ��λ�����������Ĺ����������̲��еı���[J]�����﹤�̽�չ��1999,19(4):39-44
����[80]�����죬�����⣬¬���٣����֣����ţ��������Իۣ����������γ�SAG1����ض�Ƭ��ֲ���������Ĺ���[J]���й�����������־��2004,20(8):662-665
����[81]Ԭ�Σ��ձ�Ѫ������������Ŀ�¡�������Ͳ��еı���ͻ��Լ��[D]����ɳ:����ũҵ��ѧ��2007:12
����[82]Fitchen J,Beachy R N,Hein M B.Plants virus expressing hybrid coat protein with added murine epitope elicits autoantibody response[J].Vaccine,1995,13(12):1051-1057
����[83]������CTB-PROIN��CTB-INSB�ںϻ��������ڴ˾���ֲ���еı���[D]������:���Ĵ�ѧ��2005:27
����[84]�����壮תAGD2��CTB����С����о�[D]���人:����ũҵ��ѧ��2011:18
����[85]Ruhlman T,Ahangari R,Devine A,Samsam M,Daniell H.Expression of cholera toxin B-proinsulin fusion protein in lettuce and tobacco chloroplasts oral administration protects against development of insulitis in non-obese diabetic mice[J].Plant Biotechnology Journal,2007,5(4):495-510
����[86]Kwon K C,Nityanandam R,New J S,Daniell H.Oral delivery of bioencapsulated exendin-4 expressed in chloroplasts lowers blood glucose level in mice and stimulates insulin secretion in beta-TC6 cells[J].Plant Biotechnology Journal,2012,11(1):77-86
����[87]Choi J,Diao H,Feng Z C,Lau A,Wang R,Jevnikar A M,Ma S W.A fusion protein derived from plants holds promising potential as a new oral therapy for type 2 diabetes[J].Plant Biotechnology Journal,2013,12(4):425-435
����[88]���۷����´��¸�˫����ѩ�ޣ������ۣ�ת����ֲ������������о���״��ǰ��[J]�������ѧ�붯��ҽѧ��2004,21(11):61-63
����[89]Ryan E T,Butterton J R,Smith R N,Carroll A A,Crean T I,Calderwood S B.Protective Immunity against Clostridium dif�ncile toxin a induced by oral immunization with a live,attenuated Vibrio cholerae vector strain[J].Infection And Immunity,1997,65(7):2941-2949
����[90]Arevalo-Villalobos J I,Govea-Alonso D O,Monreal-Escalante E,Zarazua S,Rosales-Mendoza S.LTB-Syn:A recombinant immunogen for the development of plant-made vaccines against synucleinopathies[J].Planta,2017,245(6):1231-1239
����[91]�Ÿ��֣���������ƽ���������Σ���С����ת����ľ����Ϊ����˸�ֲ��ڷ�����ij����о�[J]������ֲ���о���2003,25(2):223-229
����[92]Marcondes J,Hansen E.Transgenic lettuce seedlings carrying hepatitis B virus antigen HBs Ag[J].The Brazilian Journal of Infectious Diseases,2008,12(6):469-471
����[93]Joung Y H,Youm J W,Jeon J H,Lee B C,Ryu C J,Hong H J,Kim H C,Joung H,Kim H S.Expression of the hepatitis B surface Sand pre S2 antigens in tubers of Solanum tuberosum[J].Plant Cell Report,2004,22(12):925-930
����[94]Carrillo C,Wigdorovitz A,Oliveros J C,Zamorano P I,Sadir A M,Gomez N,Salinas J,Escribano J M,Borca M V.Protective immune response to foot and mouth disease virus with VP1 expressed in transgenic plants[J].Journal of Virology,1998,72 (2):1688-1690
����[95]Wigdorovitz A,Carrillo C,Dus Santos M J,Peralta A,Trono K,Gomez M C,Rios R D,Franzone P M,Sadir A M,Escribano J M,Borca M V.Induction of a protective antibody response to foot and mouth disease virus in mice following oral or parenteral immunization with alfalfa transgenic plants expressing the viral structural protein VP1[J].Virology,1999,255(2):347-35
����[96]�ζ��⣬�Ż��ɣ������٣��������������䣬��Ӣ�ۣ������ã������ң������߲���VP1����ת������������龥רһ�Ա���[J]�����\��ͨ����2011(2):107-109,173
����[97]�����ӣ���ϼ�������ã���ۣ���������������״����VP7-U�����ں��ܲ��б��������ԭ�Է���[J]����ҵ��ѧ��2012,29(12):1827-1831
����[98]Wu Y Z,Li J T,Mou Z R,Fei L,Ni B,Geng M,Jia Z C,Zhou W,Zou L Y,Tang Y.Oral immunization with rotavirus VP7 expressed in transgenic potatoes induced high titers of mucosal neutralizing Ig A[J].Virology,2003,313(2):337-342
����[99]�����ۣ�����һ�������������Ƽ�������HIV-1 gag�����gp120����ת�����Ѽ�ת����ֲ������[J]���й����﹤����־��2004,24(2):37-40
����[100]Pinkhasov J,Alvarez M L,Rigano M M,Piensook K,Larios D,Pabst M,Grass J,Mukherjee P,Gendler S J,Walmsley A M,Mason H S.Recombinant plant-expressed tumor-associated MUC1 peptide is immunogenic and capable of breaking tolerance in MUC1.Tg mice[J].Plant Biotechnology Journal,2011,9(9):991-1001
����[101]Matic'S,Quaglino E,Arata L,Riccardo F,Pegoraro M,Vallino M,Cavallo F.The rat Erb B2 tyrosine kinase receptor produced in plants is immunogenic in mice and confers protective immunity against Erb B2+mammary cancer[J].Plant Biotechnology Journal,2016,14 (1):153-159
����[102]Chotprakaikiat W,Allen A,Bui-Minh D,Harden E,Jobsri J,Cavallo F,Gleba Y,Stevenson F K,Ottensmeier C,Klimyuk V,Savelyeva N.A plant-expressed conjugate vaccine breaks CD4+tolerance and induces potent immunity against metastatic Her2+breast cancer[J].Onco Immunology,2016,5(6):e1166323
����[103]Kalbina I,Lagerqvist N,Moiane B,Ahlm C,Andersson S,Strid�@��Falk K I.Arabidopsis thaliana plants expressing Rift Valley fever virus antigens:Mice exhibit systemic immune responses as the result of oral administration of the transgenic plants[J].Protein Expression and Purification,2016,127:61-67
����[104]Yusibov V,Rabindran S.Recent progress in the development of plant derived vaccines[J].Expert Review of Vaccines,2008,7(8):1173-1183
����[105]Dalsgaard K,Uttenthal�@��Jones T D,Xu F,Merryweather A,Hamilton W D O,Langeveld J P M,Boshuizen R S,Kamstrup S,Lomonossoff G P,Porta C,Vela C,Casal J I,Meloen R H,Rodgers P B.Plant-derived vaccine protects target animals against a viral disease[J].Nature Biotechnology,1997,15(3):248-252
����[106]Martín-Alonso J M,Casta1ón S,Alonso P,Parra F,Ordás R.Oral immunization using tuber extracts from transgenic potato plants expressing rabbit hemorrhagic disease virus capsid protein[J].Transgenic Research,2003,12(1):127-130
����[107]Sohn E J,Lee Y,Park N,Park M,Kim N H,Park S,Min K,Gu S,Park Y,Song J,An D,Hwang I.Development of plant-produced E2 protein for use as a green vaccine against classical swine fever virus[J].Journal of Plant Biology,2018,61(4):241-252
����[108]Berinstein A,Vazquez-Rovere C,Asurmendi S,Gómez E,Zanetti F,Zabal O,Tozzini A,Grand D C,Taboga O,Calamante G,Barrios H,Hopp E,Carrillo E.Mucosal and systemic immunization elicited by Newcastle disease virus (NDV) transgenic plants as antigens[J].Vaccine,2005,23(48/49):5583-5589
����[109]�ձ��䵤�����������Bp BEE2������Ŵ�ת���������Է���[J]��ֲ���о���2019,39(2):287-293
����[110]�������������죬����Ƽ�������������ã���������Ac AGL15����Ŀ�¡��ֲ��������幹��[J]������ֲ�����֣�2017,14(10):2665-2670
����[111]Wei Z Y,Zhang Y Y,Wang Y P,Fan M X,Zhong F X,Xu N,Lin F,Xing X C.Production of bioactive recombinant bovine chymosin in tobacco plants[J].International Journal of Molecular Sciences,2016,17(5):1-9
����[112]Li X,Ouyang X F,Zhang Z S,He L,Wang Y,Li Y H,Zhao J,Chen Z,Wang C N,Ding L L,Peng Y,Xiao Y H.Over-expression of the red plant gene R1 enhances anthocyanin production and resistance to bollworm and spider mite in cotton[J].Molecular Genetics and Genomics,2019,294(2):469-478
����[113]Zheng L J,Zhang X G,Zhang H J,Gu Y,Huang X R,Huang H H,Liu H M,Zhang J J,Hu Y F,Li Y P,Yu G W,Liu Y H,Lawson S S,Haung Y B.The miR164-dependent regulatory pathway in developing maize seed[J].Molecular Genetics and Genomics,2019,294(2):501-517
����[114]���Ǿ꣬�����ᣬ�ǽ࣬�������������ޣ�����ǿ�����������������ø�����¡��ֲ��������幹��[J]��ֲ���Ŵ���Դѧ����2019,20(6):1613-1620
����[115]Giap D V,Jung J W,Kim N S.Production of functional recombinant cyclic citrullinated peptide monoclonal antibody in transgenic rice cell suspension culture[J].Transgenic Research,2019,28 (5):177-188
����[116]��������˫����ʥ��ά����ٻ�������ۣ��̲���ĤATPase4����Ŀ�¡���������幹�����������[J]���㽭ũҵѧ����2019,31(2):173-181
����[117]Zhou M Q,Tang W.MicroRNA156 amplifies transcription factorassociated cold stress tolerance in plant cells[J].Molecular Genetics and Genomics,2019,294(2):379-393
����[118]Queiroz L N,Maldaner F R,Mendes E A,Sousa A R,D’Allastta RC,Mendonca G,Mendonca D B S,Aragao F J L.Evaluation of lettuce chloroplast and soybean cotyledon as platforms for production of functional bone morphogenetic protein 2[J].Transgenic Research,2019,28(2):213-224
����[119]Al-Dallee Z T,Al-Hajaj M A.Expression of recombinant human glucocerebrosidase protein in sunflowers[J].Baghdad Science Journal,2019,16(1):18-33
����[120]Niu L,Yang J,Zhang J H,He H L,Xing G J,Zhao Q Q,Guo D Q,Sui L,Zhong X F,Yang X D.Introduction of the harpin Xoocencoding gene hrf2 in soybean enhances resistance against the oomycete pathogen Phytophthora sojae[J].Transgenic Research,2019,28(2):257-266
����[121]Camas Reyes A,LagunaRamírez R,Jofre Garfias A E,CardosoMartínez F,Hernández-Orihuela A L,Molina-Torres J,MartínezAntonio A.E.coli cultures expressing a synthetic sequence of ptz gene(stz) promoted in vitro direct organogenesis in Nicotiana tabacum L.[J].Plant Cell,Tissue and Organ Culture,2019,137(1):87-100
����[122]¬껣�������������֥������ҫ����ٻ�����������߲���Ӧ��WRKY����Ľṹ����Ԥ����ֲ��������幹��[J]������ֲ�����֣�2019,17(10):3253-3258
����[123]Zhang Y J,Gao W Y,Lv Y W,Bai Q Q,Wang Y J.Enhanced melatonin production via aralkylamine N-acetyltransferase overexpression enhances Na Cl resistance in transgenic Chlamydomonas reinhardtii (Volvocales,Chlorophyceae)[J].Phycologia,2019,58(2):154-162
����[124]Zhang L,Rylott E L,Bruce N C,Strand S E.Genetic modifcation of western wheatgrass (Pascopyrum smithii) for the phytoremediation of RDX and TNT[J].Planta,2019,249(4):1007-1015
����[125]Mardanova E S,Blokhina E A,Tsybalova L M,Peyret H,Lomonossoff G P,Ravin N V.Efficient transient expression of recombinant proteins in plants by the novel p Eff vector based on the genome of potato virus X[J].Plant Science,2017,8(247):1-8
����[126]�������������գ��̱�����������ѣ���־����â����������ø����Ŀ�¡����������幹��[J]���ȴ�����ѧ����2019,40(2):308-313
����[127]Ma J W,Fu X Q,Zhang T T,Qian H M,Zhao J Y.Cloning and analyzing of chalcone isomerase gene (Aa CHI) from Artemisia annua[J].Plant Cell,Tissue and Organ Culture,2019,137(1):45-54
����[128]Posgai A L,Wasserfall C H,Kwon K C,Daniell H,Schatz D A,Atkinson M A.Plant-based vaccines for oral delivery of type 1diabetes-related autoantigens:Evaluating oral tolerance mechanisms and disease prevention in NOD mice[J].Scientific Reports,2017,7(1):1-15
����[129]����������������������ؼ����������1(Cb PIN1)�Ŀ�¡��������[J]������ũѧ����2019,34(1):26-32
����[130]Swartwood K,Joyce Van Eck.Development of plant regeneration and Agrobacterium tumefaciens mediated transformation methodology for Physalis pruinosa[J].Plant Cell Tiss Organ Cult,2019,137 (3):465-472
����[131]���������������·�������ݽ��ɣ���¶���˲�BZR1�����¡��ֲ��������幹��[J]��ֲ��������֣�2019,17(23):7744-7749
����[132]Han P L,Dong Y H,Gu K D,Yu J Q,Hu D G,Hao Y J.The apple Ubox E3 ubiquitin ligase Md PUB29 contributes to activate plant immune response to the fungal pathogen Botryosphaeria dothidea[J].Planta,2019,249(4):1177-1188

֬��ø��EC3.1.1.3���ֳ�Ϊ������������ˮ��ø�����Դ�����������������һЩˮ�����������ˮ�⡢���⡢������ת���������������ϳɷ�Ӧ������֮�⣬�����ֳ�����֬ø����Ѫ��֬ø�����̴���ø������ˮ��ø���Եȣ�Tong�ȣ�2007��.֬��ø����...
1ת�������ķ�չ1.1�ҹ�ת����������״�ҹ�ת�������з���Ӧ��������3������ȡ���˻�����չ����1���������ֲ�ҵ���ƴ�����ת������Ʒ�������Ͳ�ҵ��ȡ�þ��Ч��ת����ˮ����תֲ��ø��������ð�ȫ֤�飬�߱���ҵ��������������...