ϸ������ѧ����

����ժ Ҫ�������ν���ϸ��(AS)��������ϵͳ(CNS)������Ҫ�Ľ���ϸ�����ͣ��������Ͳ���״̬�¾������Ÿ��Ӷ���Ҫ�ĵ������á�AS��CNS�еĸ��ֵ��ڹ���������ڵ�������������ء�������Ҫ��ASԴ��������о���չ����������
�����ؼ��ʣ������ͽ���ϸ��; ������; ��ϵͳ����;
�������ν���ϸ����astrocytes,AS����������ϵͳ��central nervous system,CNS������ḻ�Ľ���ϸ�����ͣ������ṩӪ�����ʡ�����ϸ��������Ũ�ȡ�Э����Ԫ��л�ȶ�����֧�ֹ���[1,2]��AS��CNS�IJ������ˣ�ȱѪȱ������֢�����ˡ������ȣ��������У���������Ӧ������������һϵ�в��������ı䡣��������ϸ�����ڵ�һ��Ĥ���ݣ���ϸ��ͨѶ������������չ�������־�������ȷ������DZ�ڹ��ܡ��о�֤ʵ��AS��CNS�����Ͳ���״̬�з��ӵĹ��ܶ������������������[1,3,4]�����Ľ�Χ��ASԴ������ķ��֡�����빦�ܼ�����CNS�����е�Ӧ��չ��������
����1�� ASԴ������ĸ���
����1.1�� ASԴ������ķ��ּ���ȡ����
������ʮ���ͳ����о����Ǿͷ�����AS��ǿ�����DZ��[5]����������һȺֱ��Ϊ40��100 nm��֬��˫���Ӳ�ṹ����[6,7]�����ɼ����������͵�ϸ�����ڣ�������Ԫ��С����ϸ������ͻ����ϸ����AS����ϸ��[8]����2007�꿪ʼ��½�����о�С�鷢��AS����Һ�д���������[9,10]��Taylor��[9]����AS���������ú���Ӧ���¿��ͷ������壬���γɹ���������ϸ����Ĥ�����γɰ���С�ݣ�С�ݽ�һ����չ�γɶ������壬��һ���Ĵ̼��¶���������ϸ����Ĥ�ںϲ��ͷŵ����⣬Ȼ����������ϸ����Ĥ�ںϲ��������á�
����Ŀǰ�����Ͻ��ҳ���ķ�������ȡ������������Ʒ�е�ASԴ�����塣һ��ͨ���ռ�ϸ�����������ȡ�����壬��Ҫ��ȡ�����г������ķ������˷����ݶ��ܶ����ķ������ߴ��鷨���Լ�����ȡ����[1,10]��Ŀǰ���ó������ķ�����ԭ��AS�������������Ƥ���Ʊ���Ϊ������AS����������������������ϸ����Ƭ������������ٶȻ�ó����������ó����ڻ���Һ��ϴ�ӡ��������ٴγ�������ȡ���������ɳ����õ������塣��һ���������ߴ��鷨��������֤��������[10]�����⣬����Ҫͨ����̬��ɢ����ʾ��Ⱥ����羵�۲���״��Western blotting֤ʵ������ʽϸ�������������ȷ�����֤���õ���������ASԴ������[8,11]��
����1.2�� ASԴ���������ɼ�����
����ASԴ��������һ��“������”��ǻ�ڳ�������ϸ�����ס����ᡢ֬�ʡ����������ӣ�����ϸ�����ӡ��������ӡ�ø�����ӣ�[10,12]���ɽ�����ϸ������Ϣͨ����Я���ĵ����ʡ�mtDNA��miRNA�ȴ��ݵ��ܱߵ���Ԫ�ͽ���ϸ����ʵ����ϸ��֮�����Ϣ���������ʽ���[10,13]������ʵ�鷢�֣������������ˮƽ����ʱ��ASԴ�������ͷ�ͻ������-1�����ܶ���ͻ����������Ԫ�Ĵ���л�����Ӱ��[14]��Ҳ�б�����AS�������������������˷�������ת�˵��ף�excitatory amino-acid transporters,EAAT)-1��EAAT-2�����൰����ά�ֹȰ�����̬�Ļ����ɷ֣�����ά��������̬[15]�������о�������ASԴ�������������ݿ˵���70��AS�������ǽͽ�ø�Ĵ���[9,16]����Щ�����Ƿ����ת�Ƶ���Ԫ�У��Ӷ�����Ԫ�Ĺ��ܲ���Ӱ�죿�������о����֣��ڼ�ή���Բ���Ӳ��֢��amyotrophic lateral sclerosis,ALS���У�AS��ͨ���������ͷ��ж����ӽ鵼�˶���Ԫ����[17]������ϸ���ܵ�һϵ����ȱ������Ѫ�Ǻ�ȱѪ��Ӧ������ʱ��AS���ͷź��з��²����õ��ף��õ�����һ�ֿɱ�����ϸ����������Ӧ����ȱ����ȱѪ�͵�Ѫ���ֺ��ĵ�����[18]���������壬��Щ���������ת�Ƶ���Ԫ�ڣ�����������[19]������������Щ�о����������ASԴ�������ȷ���Զ���Ԫ�Ĺ��ܲ���һ����Ӱ�졣
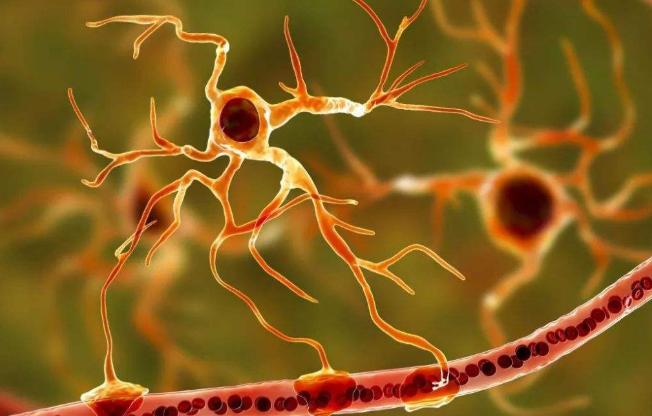
����1.3�� ASԴ��������CNS�����IJ������������е�����
������AS�ܵ���֢�̼�ʱ����ͨ���ͷ�������Ӱ��������Ԫ����ϸ���������ƶ������Ʋ��̷�չ��ALS���˶���Ԫ�������������ͣ���һ�����Խ�������ϵͳ���Լ���������Ϊ�ϡ����˶���Ԫ��AS�ڼ�����չ�з����Źؼ�����[20]�������ڼ����Գ��������绯ø��copper-zinc superoxide dismutase,SOD)-1ͻ����ALS��С��ģ�����о����֣�����ͻ����SOD1��AS��ͨ��������������Ч�ؽ�ͻ����SOD1ת�Ƶ�������Ԫ����ѡ�����յ��˶���Ԫ����[17,21]������ĸϸ�����dz��������ԭ���Զ����������������Ժ���Ϯ��������Ҫ����[22]������֤�ݱ����������������ļ���ʸ�ϸ�����ڵ���������������չ������أ���ǿ����������Ϯ�Լ����Գ̶�[22,23,24]���б����ƣ�������ϸ�����ڵ������壬���Ըı�ֲ�����ʹ�����������棬�����ڽ���AS�ٽ������������������ܽ�ASת��Ϊ��������[11]���ܵ�����Ӱ���AS��ͨ������������ʵ��miR-19a�Ĺ���ת�ƣ���������������������ͬԴ������ø��phosphatase and tensin homolog,PTEN���ı���ٽ�������ϸ��������[1,25]������ɭ����Parkinson’s disease,PD���dz������������Լ�������·��С�������ѡ���ԵĶ�Ͱ���Ԫ����Ϊ��������·��С����Ҫ��α-ͻ���˵��ͷ��ػ�������ɡ���Դ�������������Ԫϸ����AS֮��ת��α-ͻ���˵����أ��Ӷ��ٽ�PD�Ĵ�����AS��α-ͻ���˵��ij�����������֢������ͨ��ASԴ�������һ����������������ϸ������Ԫ[26]�����¼�����չ��
����2�� ASԴ��������ٴ�Ӧ���о�
����2.1�� ASԴ���������ΪCNS������ϵ�DZ�������־��
����ASԴ�����巴Ӧ������AS�IJ�������״̬�������ڶ�����Һ������Ѫ����Һ���Լ�Һ�м�[27]����֬��˫���Ӳ��Ĥ�ṹ��Ҳ�ɱ������������絰���ʡ���������ܽ���[28]������о������Ʋ�ASԴ��������ܺ�����ϵͳ���ڲ����ı�Ĺؼ���Ϣ��������Ϊһ���µ������־����������Hélène Ipas��[29]���о�����ĸϸ����ʱ���֣�����ASԴ�������к��е�2�ָ�ˮƽmicroRNA�����ܽ�����Ӱ����������½���microRNAs��һ��ḻ����Դ��С���ӷDZ���RNA��ͨ�����ưл���ı��������ظ�������ѧ����[30]���������ѭ���������е�microRNAs�������ʺ�������ϵ������־���Ϊ������Ĥ�����������ܽ��⣬��Щ���������д��ݵ�RNA��������Ҳ�㹻�ȶ�[28,29]��������ƣ�������֢��˫������ϰ����ྫ���У�microRNAsҲ����DZ�ڵ������־��֮һ����ѹ��Ӧ���£�AS���ڵ��������д���һЩmicroRNAs���ϵ����µ�[4]����Щ�没�鲨����������Ӧ�仯��microRNAs��������ASԴ�������У��Ϳ�����Ϊ��Щ�����������־�
���������ĺ�Ĭ����Alzheimer’s disease,AD����һ�ֽ����Ե��������Լ�������Ҫ����Ϊ��֪���������½��ͼ�������ɥʧ�����о����֣�AS���ڵĸ�����������������Ӿ���AD�����֪ȱ��[31]����������һ��Ĥ���������ʰ������������֬������ɣ����ڶ��ֲ������������з������ã�����ϸ��Ӧ����Ӧ����֢��ϸ��������ͬʱ�������Ŵ���ϸ�����ӵ����߶��ϵ�[31,32,33]����ˣ�������������ASԴ�����������AD������ϵ�����֤�ݡ�
����2.2�� ASԴ���������ΪCNS������ȫ�������ֶ�
��������ϸ���ܵ���������ʱ��AS���ͷŵ��������Я�������������õĺ��ᡢ�����ʣ�����������������Ԫ����ϸ������ASԴ���������Ч��Ѫ�����ϣ������ASԴ�������Ʊ���ҩ����ܰ�����ڵIJ���������ϸ�����и�Ԥ���Ӷ����ƻ���ת���̵ķ�չ��ʵ�־�ҽ�ơ�Kathrin Guitart��[19]��������������AS�������ᣨoxygen-glucose deprivation,OGD����ģ��ʱ���֣�AS�ͷŵ����������õ���ˮƽ���ߡ��ý��������������֯�ܵ�ȱѪȱ���̼�ʱ��AS�ͷź����õ��������壬��Щ�������ת�Ƶ���Ԫ�����������������á�Mingyang Deng��[34]��ͨ����H9������̥��ϸ���ֻ���������Ԫ����̥��ϸ��������ϸ����AS����������OGDģ�����о���ͬ����ϸ�����͵����������Ԫ�ı������á��ȽϷ��֣�ASԴ�������ƺ���������������ֳ����㷺������ЧӦ�����⣬��ALS���ߺ�ALS���ݶ���ģ���пɹ۲쵽�Ȱ���ת�˵���EAAT2������ѡ���Խ��ͣ��ٽ����˶���Ԫ���Ե��˷ܶ��ԡ�����������͵�G93A SOD1ͻ����ALSС��ģ���з��֣�������ĩ�ڵ�С������֯��miR-124aѡ���Խ��ͣ���ASԴ������鵼����Դ��miR-124a��������G93A SOD1ͻ����ALSС����GLT1���ı���[17,35]����һ�����ʾЯ��miR-124a��ASԴ��������ܾ��п�������GLT1��ALS�������Ե�DZ����
����3�� ������չ��
����ASԴ��������������������ѧ�е�����������Ϊһ�����˵��о�����Ŀǰ���е��о����������ASԴ����������ϵͳ�в�����������Ҫ�Ĺ��̡��������������£�ASԴ��������γɺ��ͷſ�ͨ��������ء��Ƶ�����ˮƽ���е��أ�����������Я���ĵ����ʡ��������ά��������̬����Ҫ���ʡ����������£���֢�̼�AS���ӻ������������ͷţ��ƶ�������CNS������չ����ASԴ������������ˮƽҲ���没�鷢չ���������ͣ�����Ϊ���μ�������Ҫ����Ԥ�⼲����չ�仯���������Ȼ����ASԴ�������ר���о���Ȼ���٣������о��ɹ���ͣ���ڶ���ʵ��ĽΡ���ˣ����ǻ����ȫ�桢��������о����ڼ�����Ϸ��棬��̽���������Ի������ﵰ��ASԴ����������Ϊ����ѧ������θ���������������������ˮƽ�ĸߵ;��жϼ�����չ���Լ����ȷ���������Լ��ȶ�������ҪŬ����Ŀ�ꣻ�����Ʒ��棬��̽�������������������õ�ASԴ��������ʵ�ֶ������ơ����⣬ASԴ��������Ϊ���������������ɴ�Ѫ�ܱڡ�ͨ��Ѫ�����ϣ���Ч���ⵥ�˾���ϸ��ϵͳ�����ɣ�ͬʱ�������ﰲȫ�ԡ��ȶ��ԡ����ð����Եȶ����ŵ㣬��ν�ASԴ���������γ�����ҩ������塢������ASԴ���������ҩ������ν�һ������ASԴ����������ǿ����Ч����ʵ�־����ƣ���δ�����о�����
���������
����[1]Lafourcade C,Ramirez JP,Luarte A,et al.Mi RNAs in Astrocyte-Derived Exosomes as Possible Mediators of Neuronal Plasticity[J].J Exp Neurosci,2016,10:1-9.
����[2]Goetzl EJ,Mustapic M,Kapogiannis D,et al.Cargo proteins of plasma astrocyte-derived exosomes in Alzheimer’s disease[J].FASEB J,2016,30:3853-3859.
����[3]Pegtel DM,Peferoen L,Amor S.Extracellular vesicles as modulators of cell-to-cell communication in the healthy and diseased brain[J].Philos Trans R Soc Lond B Biol Sci,2014,369:1-9.
����[4]Luarte A,Cisternas P,Caviedes A,et al.Astrocytes at the Hub of the Stress Response:Potential Modulation of Neurogenesis by mi RNAs in Astrocyte-Derived Exosomes[J].Stem Cells Int,2017,2017:1-13.
����[5]Verkhratsky A,Matteoli M,Parpura V,et al.Astrocytes as secretory cells of the central nervous system:idiosyncrasies of vesicular secretion[J].EMBO J,2016,35:239-257.
����[6]Zhang G,Yang P.A novel cell-cell communication mechanism in the nervous system:exosomes[J].J Neuro Res,2018,96:45-52.
����[7]����,������,���ӄ�.��������������ϵͳ��Ⱦ[J].�л�ҽѧ��־,2017,97:2638-2640.
����[8]Willis CM,Ménoret A,Jellison ER,et al.A Refined Bead-Free Method to Identify Astrocytic Exosomes in Primary Glial Cultures and Blood Plasma[J].Front Neurosci,2017,11:335-346.
����[9]Taylor AR,Robinson MB,Gifondorwa DJ,et al.Regulation of heat shock protein 70 release in astrocytes:Role of signaling kinases[J].Dev Neurobio,2007,67:1815-1829.
����[10]Guescini M,Genedani S,Stocchi V,et al.Astrocytes and Glioblastoma cells release exosomes carrying mt DNA[J].J Neural Transm,2010,117:1-4.
����[11]Oushy S,Hellwinkel JE,Wang M,et al.Glioblastoma multiformederived extracellular vesicles drive normal astrocytes towards a tumourenhancing phenotype[J].Philos Trans R Soc Lond B Biol Sci,2018,373:20160477.
����[12]Whiteside TL.Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment[J].Semin Immunol,2018,35:69-79.
����[13]Kojima R,Bojar D,Rizzi G,et al.Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment[J].Nat Commun,2018,9:316-317.
����[14]Wang S,Cesca F,Loers G,et al.Synapsin I Is an OligomannoseCarrying Glycoprotein,Acts As an Oligomannose-Binding Lectin,and Promotes Neurite Outgrowth and Neuronal Survival When Released via Glia-Derived Exosomes[J].J Neurosci,2011,31:7275-7290.
����[15]Gosselin RD,Meylan P,Decosterd I.Extracellular microvesicles from astrocytes contain functional glutamate transporters:regulation by protein kinase C and cell activation[J].Front Cell Neurosci,2013,7:251.
����[16]Sandoval M,Luarte A,Herrera-Molina R,et al.The glycolytic enzyme aldolase C is up-regulated in rat forebrain microsomes and in the cerebrospinal fl uid after repetitive fl uoxetine treatment[J].Brain Res,2013,1520:1-14.
����[17]Basso M,Pozzi S,Tortarolo M,et al.Mutant Copper-Zinc Superoxide Dismutase(SOD1)Induces Protein Secretion Pathway Alterations and Exosome Release in Astrocytes[J].J Biol Chem,2013,288:15699-15711.
����[18]Onodera T,Sakudo A,Tsubone H,et al.Review of studies that have used knockout mice to assess normal function of prion protein under immunological or pathophysiological stress[J].Microbio Immuno,2014,58:361-374.
����[19]Guitart K,Loers G,Buck F,et al.Improvement of neuronal cell survival by astrocyte-derived exosomes under hypoxic and ischemic conditions depends on prion protein[J].Glia,2016,64:896-910.
����[20]Jovicic A,Gitler AD.Distinct repertoires of micro RNAs present in mouse astrocytes compared to astrocyte-secreted exosomes[J].PLo S One,2017,12:e171418.
����[21]Tang L,Ma Y,Liu XL,et al.Better survival in female SOD1-mutant patients with ALS:a study of SOD1-related natural history[J].Transl Neurodegener,2019,8:2.
����[22]Figueroa J,Phillips LM,Shahar T,et al.Exosomes from GliomaAssociated Mesenchymal Stem Cells Increase the Tumorigenicity of Glioma Stem-like Cells via Transfer of mi R-1587[J].Cancer Res,2017,77:5808-5819.
����[23]Godlewski J,Ferrer-Luna R,Rooj AK,et al.Micro RNA Signatures and Molecular Subtypes of Glioblastoma:The Role of Extracellular Transfer[J].Stem Cell Reports,2017,8:1497-1505.
����[24]Ricklefs F,Mineo M,Rooj AK,et al.Extracellular Vesicles from High-Grade Glioma Exchange Diverse Pro-oncogenic Signals That Maintain Intratumoral Heterogeneity[J].Cancer Res,2016,76:2876-2881.
����[25]Zhang L,Zhang S,Yao J,et al.Microenvironment-induced PTEN loss by exosomal micro RNA primes brain metastasis outgrowth[J].Nature,2015,527:100-104.
����[26]Chistiakov DA,Chistiakov AA.alpha-Synuclein-carrying extracellular vesicles in Parkinson's disease:deadly transmitters[J].Acta Neurol Belg,2017,117:43-51.
����[27]Zhang HG,Grizzle WE.Exosomes:a novel pathway of local and distant intercellular communication that facilitates the growth and metastasis of neoplastic lesions[J].Am J Pathol,2014,184:28-41.
����[28]Zhang Y,Cai H,Chen S,et al.Exosomal transfer of mi R-124 inhibits normal fibroblasts to cancer-associated fibroblasts transition by targeting sphingosinekinase 1 in ovarian cancer[J].J Cell Biochem,2019,120:13187-13201
����[29]Ipas H,Guttin A,Issartel JP.Exosomal Micro RNAs in Tumoral U87MG Versus Normal Astrocyte Cells[J].Microrna,2015,4:131-145.
����[30]Chen B,Xia Z,Deng Y,et al.Emerging micro RNA biomarkers for colorectal cancer diagnosis and prognosis[J].Open biology,2019,9:180212.
����[31]Dinkins MB,Enasko J,Hernandez C,et al.Neutral Sphingomyelinase-2 Deficiency Ameliorates Alzheimer's Disease Pathology and Improves Cognition in the 5XFAD Mouse[J].J Neurosci,2016,36:8653-8667.
����[32]Gomez-Munoz A,Presa N,Gomez-Larrauri A,et al.Control of inflammatory responses by ceramide,sphingosine 1-phosphate and ceramide 1-phosphate[J].Prog Lipid Res,2016,61:51-62.
����[33]Shamseddine AA,Airola MV,Hannun YA.Roles and regulation of neutral sphingomyelinase-2 in cellular and pathological processes[J].Adv Biol Regul,2015,57:24-41.
����[34]Deng M,Xiao H,Peng H,et al.Preservation of neuronal functions by exosomes derived from different human neural cell types under ischemic conditions[J].Eur J Neurosci,2018,47:150-157.
����[35]Morel L,Regan M,Higashimori H,et al.Neuronal exosomal mi RNA-dependent translational regulation of astroglial glutamate transporter GLT1[J].J Biol Chem,2013,288:7105-7116

��֯��������ҽѧ�и�ϸ���������Ӧ���о�
��������һ�������������Է�����֬�ࡢ���ᡢ�����ʵȵ�Ĥ�����ݣ�ֱ����СԼΪ30��100 nm������֪����С�İ������ݡ���������ɾ���������͵�ϸ�����ڲ�����������ϸ������Ƥϸ���ȣ���ɴ�����ѪҺ���Լ�Һ����֭����Һ�С�...

ϸ��������(EV)��ϸ���ھ�Ϣ��Ӧ��״̬���ͷŵĸ��־���֬��˫��Ĥ�ṹ�������ܳ�,���ݵ�ֱ������ʮ���������ס��������л�ϸ�������ͷ�EV,���Ǵ����ڸ�������������,��ѪҺ����Һ����Һ�Լ�ϸ������ȡ�...
��������������ϸ����л��������Ҫ�����á����������ϸ���й㷺���ڵ�ϸ���ڽ���;��������С���к��е����ʡ������ϸ�����������Բ�ԭ��ȣ�������ᱻת������ø�岢����ø������ˮ��ø���⡣...

�������о��м���ʸ�ϸ�������������������
��ϸ�������Ƹβ�����о�����ʾ�������˵�Ӧ��ǰ������������Ҹ��¡�����ֻ������ߵ��ڵ����ԣ��ڶ���ģ�ͺ��ٴ��������Ѿ�֤ʵ�˸�ϸ�����и��Ƹι��ܵ����ã���δ���β����Ƶ��·���...