��ѧ����


����NAC��NAM��ATAF �� CUC��ת¼�����Ǹߵ�ֲ�����е�һ�������϶��ת¼���ӣ��� N �˶�����һ��Լ 150 ��������л��ı������У���ֲ���������������ٽ��ɡ�в��Ӧ���Լ�Ʒ�ʸ��������з�������Ҫ���ã��۷�ϼ �ȣ�2008�����ѳ�Ϊ��ǰֲ������ܼ�������������о��е��ȵ㣨Zheng et al.��2009������һ�� NAC ���� NAM �� Souer ���� 1996 ��Ӱ�ǣţ�п�¡���ģ�Soueret al.��1996������� 1997 �� Aida �ȣ�1997�������Ͻ��з����� NAM��CUC2 �� ATAF1/2 �� 3 �ֻ���CUC2 ���������� NAM����ֲ��ķ�����أ�ATAF1/2 �Ĺ�����ֲ���澳Ӧ����ء�
����Ŀǰ�о�������NAC ת¼������ˮ������������ 151 ����Ա���̲ݣ�Rushton et al.��2008���ʹ�Le et al.��2011���о��� 152 �������Ͻ����� 117 �����Ϲ��� �ȣ�2012������ѩ�ֵȣ�2010������ˮ���� SNAC1 �ǿ�����ת¼���ӣ�Hu et al.��2006�������ڷ������澳в�ȣ��ɺ������գ��� SNAC1�ı��������߿����ԣ����䵼������ͨ�� NaCl в��ɸѡ������ת����ϵ��������2011���о������ж��� NAC �������ܸ���в��ʱ�ı������ GhNAC1 ������ά������������NAC ת¼�����ڸ�����֯�о��б���� GhNAC12 �����ܵ���в�ȵ��յ���GhNAC20 �����ܵ��¡����κ� PEG �����յ���GhNAC16 ��Ҫ�Ը��δ������С����� ZmNAC1 ���Ա����¡�PEG�����κ�ABA �յ�����չ�� �ȣ�2009����ϸ���ߵ㲡����Ⱦ������ CaNAC1 �������յ������ڷǼ���������Ⱦ�뿹���źŷ��� SA �� ET ���������� CaNAC1 ���յ�����Ҳ��ǿ�ң�Oh et al.��2005����
�������й�������������ʩ�����еĵ��������������������������أ������� �ȣ�2010���������������ջ�Ҳ��Ϊ���أ�ʷ�컪 �ȣ�2004���ź��� �ȣ�2013�������ӣ�Solanum melongena L.��2n = 2x =24������ʩ�������Ҫ�߲ˣ�Ŀǰ���������ͺ��ԣ���־�� �ȣ�2000���ι��� �ȣ�2007�����棬2008���������ԣ���ѩϼ �ȣ�2010��2011������һЩ����������Ҫ���о����������������������ָ���Ӱ�죬���й����ӿ����Ի���������ֵ��о����١����ߴ������з�������� NAC1 ת¼����ȫ�����������澳в����ʱ�ձ���ģʽ������������������Ϊ���ӿ����Է������ֵ춨������
����1�� �����뷽��
����1.1 ���ϼ��䴦��
����ѡ�ñ���ũѧԺ�߲����ֿ�����ѡ������Ʒϵ ETC01���� 2012 �� 7 ����Ѯ���������ϴ�������糤�� 2 ~ 3 Ƭ��Ҷʱժȡ��������ҶƬ����Ϻ�ÿ�� 1 g������ֽ�������ɷݣ�Һ���ٶ�������–80 ����䴢�汸�á�
����2013 �� 6 ����Ѯ���������Ӳ�����ֱ�� 15 cm ��Ӫ�����ڣ�Ӫ������Ϊ��̿�U��ʯ = 1�U1������ȣ���Ӫ���������ڱ���ũѧԺũҵ������֯����ʵ�����������������糤�� 2 ~ 3 Ƭ��Ҷʱ���в�ͬ���ش�������1������Ũ�� 50 mg · L-1GA ��Һ��ÿ��Ӫ�����м��� 200 mL����2������ 4 �洦����ÿ��Ӫ�����м��� 200 mL ����ˮ������Ӫ�������� 4 ����������У���3������Ũ��Ϊ 20 g · L-1��NaCl ��Һ��ÿ��Ӫ�����м��� 200 mL��ÿ�����ش��� 3 ��Ӫ������ÿ��Ӫ���� 20 ~ 25 �����磩���� 0 ~ 12 h �ڶ�ʱȡ�������ĸ�������Ҷ��Ʒ��Һ���ٶ�������–80 ����䴢�汸�á�������������� 200 mL ����ˮ�� 1 ��Ӫ������20 ~ 25 �����磩��Ϊ���� 0 h �Ķ��ա�
����1.2 SmNAC1 ���п�¡
���� UNIQ-10 ��ʽ Trizol �� RNA ��ȡ�Լ��У��Ϻ��������﹤�̼�����������˾����ȡ������ RNA���� DNase��TaKaRa��ȥ���� RNA �л����� DNA ����Ⱦ���� RW1�������£�ȥ����RNA �еĵ�����Ⱦ������� Thermo ��˾�� NANODROP 2000 �����Ũ�Ȳ����� 1%����֬��������Ӿ����� RNA �������ԡ�����ȡ���� RNA Ϊģ��ϳ� cDNA��cDNA �ϳ���ϵ�������Ϊ 20 μL����Ӧ����Ϊ���� RNA �� 10 μmol · L-1Oligo��dT��18 ���Һ�� 70 ��ˮԡ 10 min��Ȼ����� ReverseTranscriptase M-MLV ��ת¼ø��200 U · μL-1����dNTP��10 mmol · L-1����RNase Inhibitor��40 U · μL-1���������˫��ˮ������ 42 �汣�� 60 min������ 70 ����� 15 min��������–20 �������Ϊ����������ʵʱӫ�ⶨ�� PCR��Real-time quantitative PCR��RT-qPCR����ģ�屸�á�
������ NCBI ������������ GenBank ���ѹ��������� NAC��AY714222�����������Ͳ� NAC1��AY245879�������� NAC��AY573802�������� NAC1��NP_001234482.1����Ƽ沢�������� SmNAC1�ı����������� S1��CAYCCDACKGAYGAAGA������ A1��TTCTTSTTGTADATTCGACA��
�������� Clontech �� SMARTerTMRACE cDNA Amplification Kit �ֱ��¡ SmNAC1 �� 3′�˺� 5′�����У���Primer Primier 5.0�����������3′S��GCTGGAAAAGCACCAAGAGGAAT��5′A��GGTTCCTGCTGCTCGGTTCGGC��UPM ͨ������ Long��5′-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3′��Short��5′-CTAATACGACTCACTATAGGGC-3′���� RACE �����Լ����ṩ��94 ��Ԥ���� 4 min��94 �� 30 s��72 �� 3 min��5 ��ѭ����94 �� 30 s��70 �� 30 s��72 �� 3 min��5��ѭ����94 �� 30 s��68 �� 30 s��72 �� 3 min��25 ��ѭ����72 ������ 10 min��4 �汣�档
�������� UNIQ-10 ��ʽ DNA �������Լ��У��Ϻ�������������������գ�������Ӿ�����ñ���ȫʽ�����\����˾ pEASY-T3 Cloning Kit ���ӣ�ת���� Trans1-T1 ����̬ϸ�������װ�ɸѡ������Һ PCR �����������ͱ�������Զ־���\���������ι�˾���������� DNAMAN ����ƴ�����л������ SmNAC1 ȫ�� cDNA ���С�����ƴ�ӳ����������� Primer Premier 5.0 �������������QS��AAAATAAACAGCGAAGAGAGA��QA��CTACACCAAAAAACCAAGAAA ���� SmNAC1 ȫ����ͬ��������������ա����ӡ�ת����ɸѡ���Ͳ���
����1.3 ʵʱӫ�ⶨ�� PCR
�������� SYBR Green ӫ��Ⱦ�Ϸ����� CFX96��Bio-Rad��ʵʱӫ�ⶨ�� PCR �Ƕ� SmNAC1 �ڲ�ͬ�����µ�ʱ�ձ�����м�������Ficko & Cernelc��2005��������Ŀ����� SmNAC1 ����������SmNAC1-S��CTTGAGGCTTGATGAATGG ���������� SmNAC1-A��GTGGTAATTGTGTGGGTAAA��
���������ڲλ������٣�2011��Smβ-ACTIN��GenBank ��¼�ţ�GU984779�������� Smβ-S��GTCGGAATGGGACAGAAGG��Smβ-A��CAGTCAGGAGAACAGGGTG�����ձ����﹫˾�Լ���Real Master Mix��SYBR Green��PCR ����������У�����ת¼���õ� cDNA �ֱ�����ݶ�ϡ�ͣ�1��1�U10��1�U100��1�U1 000��1�U100 000������ӫ�ⶨ�� PCR ��Ӧ��������Ա����ߡ����� cDNA�ʹ��������� 3 ���ظ���
����RT-qPCR �����ķ�Ӧ��ϵΪ 20 μL������ 10 μL Real Master Mix ���Һ��2 μL cDNA��0.5 μLSmNAC1-F��10 μmol · L-1����0.5 μL SmNAC1-R��10 μmol · L-1����7 μL ddH2O����Ӧ����Ϊ��94 ��Ԥ���� 2 min��94 ����� 20 s��53 ���˻� 25 s��72 ������ 20 s��ÿ��ѭ���� 3 ������ӫ��ɼ���40 ��ѭ�����ڲλ���� PCR �����ķ�Ӧ��ϵΪ 20 μL��10 μL Real Master Mix ���Һ��2 μL cDNA��0.5 μL Smβ-S��10 μmol · L-1����0.5 μL Smβ-A��10 μmol · L-1����7 μL ddH2O���䷴Ӧ������ SmNAC1�ķ�Ӧ������ͬ��
����2�� ��������
����2.1 SmNAC1 ���еĻ��
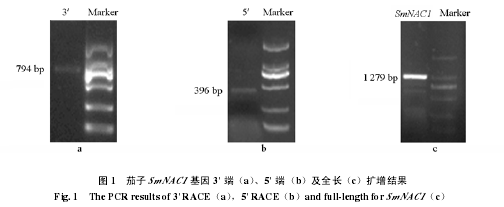
��������������ֱ���л��ա���¡����������õ��� 426 bp ���м����У�Blast ���߶ԱȺ����֤ʵ�� NAC ������壬���� NAC ���屣�ؽṹ������������������Ƶ�������������ջ�� 794 bp ���� 3′�� cDNA Ƭ�����У�ͼ 1��a������ 396 bp �� 5′�� cDNA Ƭ�Σ�ͼ 1��b������������ QS �� QA ���������������� SmNAC1 ȫ������ͼ 1��c������������ʾ cDNA ȫ������Ϊ1 279 bp��NCBI ���߷���ͬԴ�Ժ������û���Ϊ SmNAC1����¼�ţ�KF668813����
����
�����û�����1������302������������������Ķ���������ͼ2����ʼ������ATGλ��124 bp����ֹ�� TAG λ�� 1 032 bp �������� DNAMAN �Ʋⰱ������������������ NAC �������н��ж������бȶԣ�ͼ 3�������� SmNAC1 ���� NAC ���干���ص㣬�� N �˶�����һ�α��صİ��������У�Լ 150 ��������л������������ A��B��C��D��E �� 5 ���ǽṹ��
����

����2.2 SmNAC1 �������з������ṹԤ��
����Protparam ���߷�����ʾ������ SmNAC1 ������ 20 �ֻ����İ�������ɣ������ʽΪC1570H2376N430O461S12�����к�����ߵİ������� Lys��7.6%����Pro��7.6%����������͵İ������� Cys��1.3%�����������ᣨAsp + Glu������Ϊ 38 �����������ᣨArg + Lys������Ϊ 40 ����Ԥ����Է�����ԼΪ 35.04 kD�����۵ȵ��Ϊ 8.18����ƽ����ˮ�ԣ�GRAVY��Ϊ–0.858�����ȶ�ָ�� 37.02���Ʋ��������ȶ����ס�
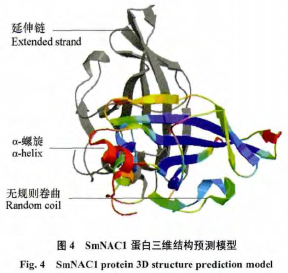
�������� NPSA �е� MLRC ����Ԥ��������ṹ��SmNAC1 ���װ����� 63 �� α–������Alpha helix����ռ 20.86%��41 ����������Extendedstrand����ռ 13.58%��198 �����������Randomcoil����ռ 65.56%������ TMHMM Server v.2.0 Ԥ��˵�����Ĥ �� �� �� PSORT WWW Server �� PSORTPrediction ����Ԥ��ϸ����λ��ϸ�����С��ڵ����ʽṹ���ݿ�����������ͬԴģ�ͣ�������վExpasy �� Swiss Model ����ͬԴ��ģ���Ʋ⵰����ά�ṹģ����ͼ 4��
����
����2.3 SmNAC1 ���������� NAC ��ͬԴ�Լ�ϵͳ����������
������ SmNAC1 ��������ͨ�� NCBI �� BLAST ���߷���ͬԴ�ԣ�������� SmNAC1 �뷬�� NAC1��NP_001234482.1������������CAC42087.1�������� NAC2��ADL60119.1������AAW48094.1�����̲ݣ�ADQ08688.1����ͬԴ�Էֱ�Ϊ 90%��90%��89%��84%�� 82%��
������һ��ͨ�� DNAMAN ������ SmNAC1 �� GenBank ���ݿ����Ѿ���¼�� 14 �� NAC ���������й���ϵͳ��������ͼ 5�����ɼ����� SmNAC1 �뷬�ѡ����������̲���Ե��ϵ����������Ͻ� ATAF2�ڽ���������ͬ����Դ���밫ǣţ�����Ͻ� CUC2��CUC1 ��Ե��ϵ��Զ���Ʋ����� SmNAC1 ͬ������ ATAF2 ��ֲ���澳Ӧ����صĹ��ܡ�
����
����2.4 SmNAC1 �ڲ�ͬ�����µ�ʱ�ձ������
����ͨ�� RT-qPCR ���ڲλ������Smβ-S��Smβ-A���� SmNAC1 �������������SmNAC1-S��SmNAC1-A�����м������۵�����ͼ��ʾ�ڲλ���� SmNAC ��ֻ�� 1 ���壬Tmֵ�ֱ�Ϊ 82 ���76 �棬�����ڲλ���� SmNAC1 �������ﶼ���и߶������ԣ�������������ʾ�ڲλ���� PCR����Ч�ʺ����ϵ���ֱ�Ϊ 90.8%�� 0.997��SmNAC1 ������Ч�ʺ����ϵ���ֱ�Ϊ 112.5%�� 0.997������������нϸߵ��ظ��Ժ�����Ч�ʡ����ӫ�ⶨ�� PCR ��������������Ӿ������Ŀ��Ƭ�Σ�����������塣
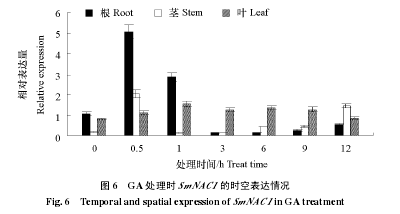
������ 50 mg · L-1�� GA �����������磬ʵʱӫ�ⶨ�� PCR ����������Ҷ�� SmNAC1 ����Ա�������ͼ 6���ɼ�������ʱ��ı仯 SmNAC1 �ڸ�������Ҷ�о��б��ͬʱ�ڶ�ʱ���ڱ������������ﵽ���ֵ�������� 0.5 h���ڸ��;��еı������� 0 h ʱ��ȣ���Լ�ֱ����� 4 ���� 8 ��������½����ϵ�ˮƽ���������� 0 h ʱ���������������ָ�����ҶƬ���ڴ��� 1 h ʱ�ﵽ��ߣ��� 0 h ʱ���ԼΪ�� 2 ���������������ʱ��仯�������ƽ�С��Լ�� 0.8 ~ 1.5 �䲨����
����
����4 �����в���£�SmNAC1 �ڸ�������Ҷ�ж��б��ͼ 7����0.5 h ʱ���е���Ա������½�����ͣ�����Ҷ�ж��� 0 h ���ߣ������϶���ʱ����ӳ������ߣ�˵���ڲ�ͬ�����е���в�ȴ�����SmNAC1 �ı��ﶼ��һ�����յ����ã����ڸ���ҶƬ�У�в�� 9 h ʱ����Ա������ﵽ���ֵ���ھ��д��� 6 h ʱ������ˮƽ��ߡ�
����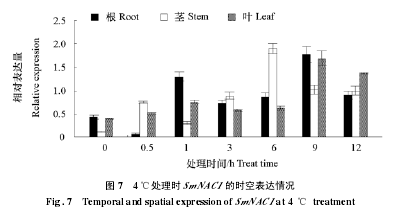
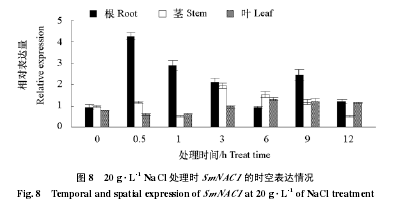
������ͼ 8 ��֪���� 20 g · L-1�� NaCl в���£����Ӹ�������Ҷ�� SmNAC1 ���б�����еı������仯�ϴ���Ҷ�в�����ƽ�����ڸ��д��� 1 h ʱ SmNAC1 ����Ա������� 0 h �� 0.95 �ﵽ��ֵ 4.26���ɼ� SmNAC1 �ڸ��ж�ʱ��Ѹ�ٵ��յ���������ʱ���ӳ�����Ա��������ͣ��ھ��д��� 1 h ʱ��Ա������������ߣ�6 h ʱ�ﵽ���ֵ 1.95����ͣ���ҶƬ�� 9 h ʱ�����ֵ 1.32���������� 0.59 ~ 1.32 ��Χ������
��������������ɼ��������Ӹ�������Ҷ�� SmNAC1 ���ܱ��˵���˻������ĵ�����һ�������Ӹ�������Ҷ�������ձ���ڵ�ת¼���ӣ���Դ GA �ͷ�����в�ȴ�����������������仯�����������������ƣ������϶� SmNAC1 ��һ�����յ������ת¼���ӣ�������Щ��ͬ������ SmNAC1�����ˮƽ��ͬ�����еı���ˮƽ������Ը��ھ���ҶƬ�У���ʾ�ڸ������Ʊ����ҶƬ�б���ˮƽ����ȶ����ɼ���ת¼�����ڲ�ͬ���ٷ��������������������һ�����졣
����
����3�� ����
����NAC ��Ϊֲ�������һ��ת¼���ӣ���������ֲ�������������̵ĵ��ء����ص��أ�������ֲ�������������в��Ӧ����Ҳ������Ҫ��������ع��ܡ����о��д������п�¡��ȫ�� 1 279 bp�� SmNAC1 �����뺬�� 302 ������������������Ķ����� NAC ���徭�䱣��������A��B��C��D��E �ǽṹ���ܸ����� DNA ����������ϣ�Ernst et al.��2004��������Ԥ����Է�����ԼΪ 35.04 kD�����۵ȵ��Ϊ 8.18���Ʋ���Ϊ�ȶ����ס��ۺϰ���������ͬԴ�Է�����ϵͳ������������SmNAC1 �뷬�ѡ����������̲���Ե��ϵ����������Ͻ� ATAF2 �ڽ���������ͬ����Դ�����ܾ��� ATAF2 ��ֲ���澳Ӧ����صĹ��ܡ�
�������о���ʾ�����Ͻ��� NAC1 ����鵼�������������е��������źţ�����ǿ����������Ũ�ȡ����ԭ���ķ������ʸ߶�����أ������������������ l kb ��Χ�ں��� 3 ����ù�ط�ӦԪ����GAresponse complex���������ܳ�ù�����õ�������Xie et al.��2000�����̲� NAC1 Ҳ�ڸ��⡢���ԭ�������Ա�����ѻ���2005����֤�˸û���ı������������յ����ҽ�һ�������������غͳ�ù�ض���ı�����յ�ЧӦ�����о��м������ʩ GA �����Ӳ�ͬ���ٵ� SmNAC1 ����Ա�������ͬ����֤�˳�ù���ܹ��յ����� SmNAC1 �ı�������������̲ݵ�ͬԴ�Ժܸߣ�ͬ���ڸ����յ�����ˮƽ�ϸߣ����Dz���ֻ�ڸ��������Ա������Ҷ�о��б�����ھ����յ�����仯������������ƣ�����Ҷ�б������ά���ȶ�ˮƽ���б��������Ͻ��д��ھ��з�������ø���Ե� SIAT5 ���ף��ܽ� NAC1 ���ػ������� NAC1 ����Ѹ�ٽ��⣬�о���ʾ SINAT5 ���������ͨ�����ػ���NAC1 ���Ŀ��ٽ��⣨Xie et al.��2002�������������ɱ��о��� SmNAC1 �ڸ��;��б�����Ѹ���������������ָ���ԭ��ˮƽ����Ҫԭ��
�����������ȣ�2012��������ˮ�� OsNAC ��������ڸ��Ρ��ɺ�������в�Ⱥ�ı���ˮƽ������45 ����в����Ӧ OsNAC ������ 33 ���ϵ����12 ���µ����44 ������в����Ӧ OsNAC ������31 ���ϵ����13 ���µ������������ SNAC1 ת����ˮ�����������ͬʱ��ǿ������������ʵ����� 22% ~ 34%���������� OsNAC6/SNAC2��OsNAC5��ONAC045 �� OsNAC10 ��������������ˮ���Ըɺ������ε������������о����� 4 ����¼� 20 g · L-1NaCl в���������磬��ⲻͬ���٣���������Ҷ���� SmNAC1 ���б����Ȼ��ʾ�ڸ��е����Ʊ�������Կ��� SmNAC1 ����������һ���ձ���ڵĿ��յ������ת¼���ӣ��ܵ����¡����εķ�����в�Ⱥ���ʾ�ϵ����
����SmNAC1 ���ܲ��������ӶԵ��º���в�ȵ����ܵ��ڹ��̣�����Ԥ��������� SmNAC1 ����������ӶԵ��º��ε������ԣ�Ϊ�������ӷ�����в���Ŵ������о��춨��һ�������ۻ�����

���ӣ�SolanummelongenaL.�������ѿ�������ֲ��һ���1�����ң�С֦Ϊ��ɫ��ҶƬһ��Ϊ���λ��߳�Բ״�����Σ�����һ����6�������ϣ�ҶƬ��ɴﵽ15�������ϣ������ӵ�Ʒ�ּ����ƶ��������ӻ������һ��Ϊ��ɫС����ϡ��״�������ӹ�ʵ�ɹ���ʳ...
���ø߿������ߵ���ľ�����Ӽӵ���������
������������ʡ���������Ѹ������,���ӵ��������Ҳ����������,�������ٶ�Զ����ڻƹϺͷ���;����Ҫԭ������ʡ�紺���µ�,��ɵ����ӻ�ή���������Բ�������,�����ӻ�ή��Ϊ��������,ֲ�귢�����ʵ���Ա�...
֣��1����֣�����߲��о���ѡ��������һ���ӽ���Ʒ�֣���ʵ��Բ�Σ������ʵΪ��ɫ����Ƥ�⻬��������Ʒ��״�ѡ���ʵƽ��������0.46kg,����ɴ�1.3kg.ÿ667m2����4000һ5000kg,�߲��ɴ�6000kg���ϡ��ʺ��紺¶�ؼ����ﱣ������ֲ��֣��1�����紺��...