园艺学论文


辣椒属(Capsicum)主要有 5 个栽培种(邹学校,2009),即一年生辣椒(C. annuum)、中华辣椒(C. chinense)、浆果状椒(C. baccatum)、灌木状椒(C. frutescens)和茸毛椒(C. pubescens),其中一年生辣椒是分化最多、栽培最广的 1 个种,而中华辣椒是亚马逊地区栽培最为广泛的 1 个种。
另外,辣椒属还有许多近缘野生种,其中有部分已被利用,如茸毛椒。国内外科技人员在辣椒种质资源分子鉴定方面进行了全面的研究,取得了一些阶段性成果。Lefebvre 等(2001)采用 AFLP 和 RAPD 标记以及结合表型数据对辣椒自交系开展遗传多样性研究。
陈学军等(2007)通过 RAPD 和 ISSR 对辣椒属的 5 个栽培种 13 份资源进行了研究,能将一年生辣椒与其他 4 个栽培种区分开来。Sanatombi 等(2010)采用 RAPD 的标记对印度曼尼普尔邦 7 个当地品种开展了种质资源鉴别工作,结果表明 7 个品种分别属于一年生辣椒、中华辣椒和灌木状椒。
李永平等(2011)利用 RAPD 分子标记技术对辣椒种质资源遗传多样性进行分析,结果将供试的 90份种质分为 5 大类群。陈学军等(2012)采用 SRAP 和 SSR 标记对中国灌木辣椒开展了遗传多样性研究。孟金贵等(2012)应用 ISSR 标记开展了涮辣与辣椒属 5 个栽培种亲缘关系的研究。上述研究为辣椒遗传育种及产业化发展奠定了基础。
在核基因组研究中,利用核糖体 DNA 内转录间隔区(Internal Transcribed Spacer,rDNAITS 区)序列对药用植物菲兰(Lee et al.,2006)、红山茶(田敏 等,2008)、石斛(栗丹 等,2012)、桑(陈仁芳 等,2010)、扁桃(邱蓉 等,2012)等进行鉴定和系统进化分析。ITS 区在进化过程中受到的选择压力非常小,且进化速率较快,另外 ITS 区序列具有保守性,不仅适合于低分类群的系统发育分析,而且可以反映种间与种内水平的变化,亦被用于种内各居群内的差异性研究(丁小余 等,2002;德英 等,2012)。近年来,ITS 作为常用分子标记已被用于辣椒属植物的系统分类(蒋向辉 等,2010;Purkayastha et al.,2012),同时也发现了部分种类的 ITS 存在多态性。Ryzhova 等(2002)对 1 个浆果状椒、2 个中华辣椒和 1 个一年生辣椒,蒋向辉等(2010)对 1 个湖南当地农家品种‘浏阳朝天椒’,Purkayastha 等(2012)对 6 份辣椒素极高的辣椒材料‘Bhut Jolokia’进行了 rDNA-ITS 序列分析,由于研究材料类型与数量不足,代表面较窄,研究结论具有一定局限性。
本研究中通过多年的调查,收集了国内外不同来源的一年生辣椒和中华辣椒共 17 份,并从GenBank 下载辣椒属的主要栽培种浆果状椒(Bohs & Olmstead,2001)、茸毛椒、灌木状椒(Purkayasthaet al.,2012)和野生种 C. eximium(Whitson & Manos,2005)、野生种 C. lycianthoides(Smith & Baum,2006)的 ITS 序列,结合本研究材料的 ITS 序列,分析 ITS 长度、(G + C)含量及其变异、遗传分歧和同源性,探讨辣椒资源的系统位置与进化关系。
1、 材料与方法
1.1 材料及 DNA 提取
试验于 2012 年在广东省蔬菜新技术重点实验室完成。试验材料由广东省农业科学院蔬菜研究所辣椒育种与生物技术团队提供(表 1),8 份材料由国外引进,其中一年生辣椒 14 份,中华辣椒 3份。采摘苗期鲜叶片,用第 7 或 8 片真叶提取基因组 DNA,提取方法参照 CTAB 法(王关林和方宏筠,2002),然后将提取的 DNA 放入–20 ℃备用。
1.2 PCR 扩增及测序
聚合酶链式反应(PCR)体系为 20 μL,包括 10× PCR buffer 2.5 μL,25 mmol · L-1MgCl21.2 μL,模板 DNA(40 ng · μL-1)1.0 μL,10 μmol · L-1上下游引物各 1.0 μL,Taq 酶(5 U · μL-1)0.2 μL,2.0mmol · L-1dNTP 2.0 μL,加 ddH2O 补足 20 μL。参照 White 等(1990)的方法设计 ITS 区扩增所用通用引物,ITS4(5′-TCCTCCGCTTATTGATATGC-3′)和 ITS5(5′-GGAAGTAAAAGTCGTAACAAGG-3′)。ITS-PCR 反应程序为:95 ℃预变性 3 min,94 ℃变性 30 s,52 ℃退火 30 s,72 ℃复性40 s,共 35 个循环后,72 ℃延伸 7 min,4 ℃保存。
PCR 产物纯化回收:北京鼎国 NEP013-1 DNA 片段快速纯化/回收试剂盒;连接:pEASY-T1Simple Cloning Kit CT111-01 试剂盒。回收纯化后的产物连接于 pEASY-T1 Simple Cloning Vector 上,随后加连接产物于 Trans 1-T1 感受态细胞中,37 ℃静置培养过夜。挑取单克隆 PCR 方法鉴定阳性克隆。Invitrogen 公司测序。为确保所测序列的准确性,测序过程中使用扩增引物进行双向测序。
1.3 数据分析
测序原始峰图使用 Contig Express 软件拼接,而后用 BioEdit 手工校对,用 Clustal X 对所得序列进行排列。DNAStar 软件计算 ITS 序列的遗传分歧及同源性百分比差异(Kimura,1980;Tamuraet al.,2011)。ITS 序列分析采用 MEGA 5.05 软件,以茄科番茄属(Lycopersicon)栽培种‘上海 906’作为外类群植物,采用邻位相接法(Neighbor-Joining,NJ)构建系统树。系统树的每个分支的统计学显著性分析以自展法(bootstrap)进行检验,重复次数为 1 000 次。
2、 结果与分析
2.1 ITS 序列长度、(G + C)含量及变异
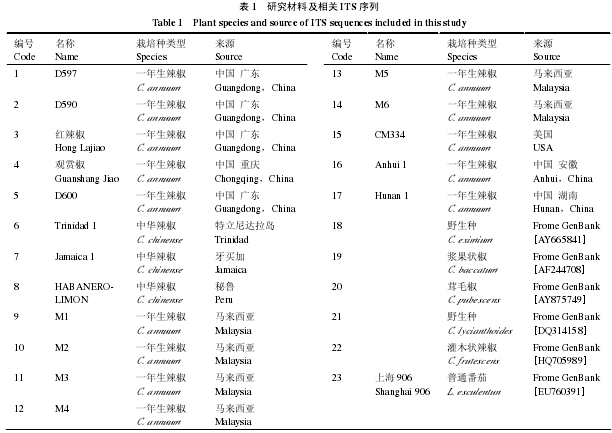
根据 GenBank 中已报道的辣椒 ITS序列(Accession No. AY665841),确定 17 个辣椒样品的 rDNAITS 序列中 ITS1 和 ITS2 与 3 个编码区 18S、5.8S 和 26S 的界限。17 个辣椒样品的 ITS 序列(只包含 ITS1,5.8S rDNA 和 ITS2)长度变化范围在 632 ~ 651 bp(表2),14 个一年生辣椒中,编号 1 的辣椒材料 ITS 序列最短,为 632 bp,编号为 5、12 和 13 的 3 个一年生辣椒材料 ITS 序列全长为 650 bp,其余 10 个一年生辣椒材料 ITS 序列全长为 651 bp。编号为 6、7 和 8 的 3 个中华辣椒 ITS 序列全长为 644 bp,比一年生辣椒 ITS 序列全长短。(G + C)含量为 49.38% ~ 63.51%,转换颠换比 3.37。当空位(gap)作缺失处理时,ITS 区全序列排序后的长度为 660 个位点,其中有 237 个变异位点,173 个简约信息位点,分别占 35.91%和 26.21%。在 17 个ITS 序列中,A 占碱基总数的 25.5%,T 占 24.0%,C 占 28.8%,G 占 23.7%。
分析表明,ITS1 序列长度范围 227 ~ 238 bp,相差 11 个碱基。编号为 5、12 和 13 的 3 个一年生辣椒 ITS1 序列长为 237 bp,其它 11 个一年生辣椒 ITS1 序列长为 238 bp。编号为 6、7 和 8 的 3个中华辣椒 ITS1 序列长为 227 bp,比一年生辣椒 ITS1 序列短。A 占碱基总数的 24.4%,T 占 20.5%,C 占 31.2%,G 占 23.8%,(G + C)含量变幅为 50.0% ~ 66.52%。当空位(gap)作缺失处理时,ITS1区全序列排序后的长度为 240 个位点,其中有 89 个变异位点,占总位点的 37.08%,70 个简约信息位点,占总位点 29.17%。以野生种 C. eximium 的 ITS 序列为参考序列,利用 Clustalx 2.1 软件进行比对,发现编号 6、7、8、20 和 22 在 ITS1 区有 15 个碱基缺失。
ITS2 序列长度在 234 ~ 257 bp,相差 23 个碱基,相对 ITS1 区来说变化较大。编号为 1 的一年生辣椒 ITS2 序列长为 234 bp,其它 13 个一年生辣椒 ITS2 序列长为 253 bp。编号为 6、7 和 8 的 3个中华辣椒 ITS2 序列长为 257 bp,中华辣椒 ITS2 序列长度较一年生辣椒的要长。A 占碱基总数的18.5%,T 占 23.9%,C 占 32.1%,G 占 25.5%,(G + C)含量变幅为 52.96% ~ 68.09%。当空位(gap)作缺失处理时,ITS2 区全序列排序后的长度为 257 个位点,其中有 96 个变异位点,占总位点的37.35%,65 个简约信息位点,占总位点 25.29%。
5.8S rDNA 序列长度为 160 bp,长度高度保守。A 占碱基总数的 29.3%,T 占 26.5%,C 占 21.5%,G 占 22.7%,(G + C)含量变幅为 41.25% ~ 53.75%。5.8S rDNA 区全序列有 52 个变异位点,占总位点的 32.5%,40 个简约信息位点,占总位点 25%。
2.2 ITS 序列遗传分歧及同源性分析
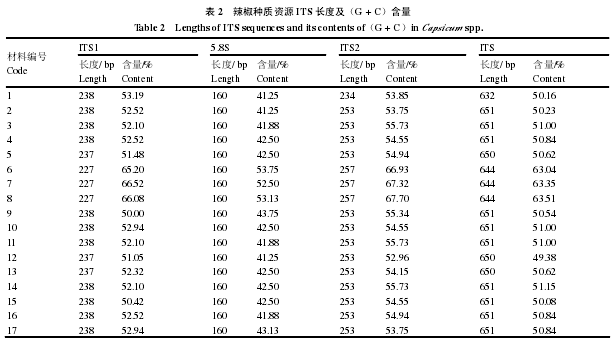
用 DNAStar 软件计算 17 个 ITS 序列的遗传分歧及同源性百分比差异,结果表明,这 17 个 ITS序列的同源性(percent identity)约 25.2% ~ 100%,遗传分歧在(divergence)约 0 ~ 3.39(表 3)。材料 13 与材料 5、7、8 的遗传分歧较大,分别为 3.06、3.32、3.28,材料 4 与材料 7 遗传分歧最大为 3.39。
2.3 系统进化和聚类分析
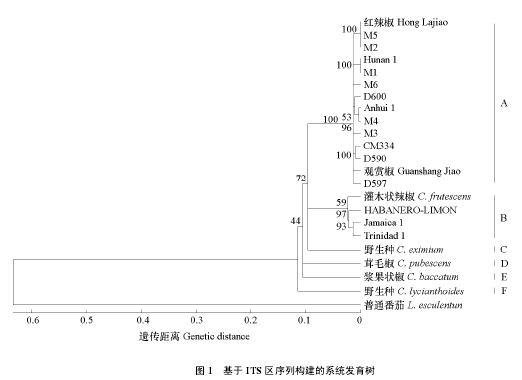
以茄科番茄属栽培种‘上海 906’作为外类群,根据试验获得的 17 个 ITS 序列以及从 NCBI 获得的 5 个不同辣椒种的 ITS 序列,用 MEGA4 进行系统进化聚类分析,结果表明,茄科番茄属与辣椒属的植物分化十分明显,分别为一单支系(图 1)。进化树中 23 个 ITS 的平均遗传距离为 0.238,22 个辣椒 ITS 之间的平均遗传距离为 0.159,外类群茄科番茄属 ITS 与 22 个辣椒属 ITS 之间的平均遗传距离为 1.120。在进化树标尺约 0.11 处,22 个辣椒属 ITS 序列可分为 6 个分支。
分支 A 共 14 个辣椒样品全为一年生辣椒,包括 7 个国内收集的资源,还包括 6 个引自马来西亚的辣椒资源,以及从美国收集的材料,这 14 个辣椒样品平均遗传距离为 0.041。分支 B 包括 3 个中华辣椒和灌木状辣椒,该支平均遗传距离为 0.039,表明中华辣椒与灌木状辣椒亲缘关系相对较近。分支 C、D、E、F 等分别只有 1 个辣椒样品,分别为野生种(C. eximium)、茸毛椒、浆果状椒、野生种(C. lycianthoides)。
由图 1 可知,14 个一年生辣椒分别与灌木状辣椒和中华辣椒有较近的亲缘关系。分支 D 与分支A、分支 B、分支 E、分支 F 的遗传距离平均分别为 0.077、0.055、0.086、0.089,由此表明浆果状椒与野生种(C. eximium)和茸毛椒亲缘关系相对较近。而野生种(C. lycianthoides)与茸毛椒亲缘关系相对较近。
3、 讨论
本研究中 17 个辣椒样品通过 ITS 区域克隆所获得片段长度均在 750 bp 左右,这与前人研究辣椒 ITS 序列的结果(Ryzhova et al.,2002;Purkayastha et al.,2012)相吻合。有研究表明(Bruce etal.,1995),有花植物(flowering plant)的 ITS 序列一般比其它生物要短,在 187 ~ 298 bp 之间。
本研究中的 17 个辣椒样品的 ITS(ITS1 和 ITS2)序列变幅为 227 ~ 257 bp 之间,均在有花植物 ITS序列长度变化范围之内,与前人研究的辣椒 ITS 序列长度(Whitson & Manos,2005;Hernández et al.,2010;蒋向辉 等,2010)基本一致。分析表明辣椒 ITS1、ITS2 变异位点分别是 89 和 96 个,占总位点的 37.08%和 37.35%,由此表明 ITS2 区变异稍大于 ITS1,这与 Purkayastha 等(2012)的观点相一致。而分析表明,5.8S 区均为 160 bp,在长度方面高度保守。
本研究中取材 17 份,对 ITS 序列进行分析,这 17 个 ITS 序列的同源性约 25.2% ~ 100%,遗传分歧在 0 ~ 3.39 之间,说明辣椒 ITS 序列同源性变幅相对广泛,这可能与试材选取以及试验方法有关,尚需进一步研究。另外,引自马来西亚的一年生辣椒(6 个)与收集于中国内地的一年生辣椒(8 个)处于同一分支 A,未发现种内地理迁移的变化规律,这从另一角度说明一年生辣椒 ITS序列具有一定的保守性,种性比较稳定,非近期分化类群。
ITS 为中度保守序列且长度不大,在种内具有较高的保守性,同时进化速率较快;但在不同种间有不同的变异,不仅广泛用于物种间的鉴定,而且可以用来进行种内居群间的差异性分析。辣椒ITS 序列比对结果发现中华辣椒(编号 6、7、8、20)和灌木状辣椒(编号 22)在 ITS1 区有 15 个碱基缺失,为证实灌木状辣椒与中华辣椒被分在一起提供了分子证据。
本研究结果表明,一年生辣椒与中华辣椒以及灌木状椒有较近的亲缘关系,这与陈学军等(2007)采用表型数据以及 RAPD 和 ISSR 标记研究的结果一致,也与前人基于形态学和细胞学的研究结果(Pickersgill et al.,1979;Pickersgill,1991)一致。在辣椒的系统进化、聚类分析中,基于 ITS 序列(包括 ITS1、5.8S 和 ITS2)分类结果表明,不同地理种群的一年生辣椒处于同一分支 A。
但在本研究的NJ系统树中,灌木状辣椒与中华辣椒被分在一起,未能将它们区分开来,与Purkayastha等(2012)报道的研究结果相一致。同时也表明,借助于 ITS 序列差异还不能够把辣椒属不同种完全鉴别,需要借助其它的分子标记才能进行有效区分。

山葡萄是东北地区的特产,可酿造出优质葡萄酒,深受广大消费者欢迎。山葡萄从建园开始,到进入盛果期,一般需5年左右。目前大面积栽培山葡萄多数采用篱架,少部分为小棚架。山葡萄整形与修剪目的在于调节生长和结果,合理配置枝蔓。1整形山葡萄在东北可以露地越...
凤仙花属(ImpatiensL.)植物是中国着名传统花卉之一,也是世界着名观赏花卉[1]。全球范围内最主要的分布区有5个,分别是:东亚区、部分东南亚地区、印度南部和斯里兰卡、热带非洲区和马达加斯加区[2-5]。该属是凤仙花科Balsaminaceae(仅有凤仙花属和水角属Hydro...
正红菇(RussulagriseocarnosaX.H.Wang,ZhuL.Yangudsen)属真菌界担子菌门(Basid-iomyeota)担子菌纲(Basidiomycetes)红菇目(Rus-sulalesDreiselexP.F.CannonJ.C.David)红菇科(RussulaceaLotsy)红菇属(RussulaPers)[1],具有食用、药用...
芋艿[Colocasiaesculenta(L.)S.]俗称芋头,为天南星科植物芋的块茎。万年芋艿属多子芋类中的香沙芋艿品种,主要分布在江苏海门、启东、靖江、上海崇明等长江下游地区。因生长于黏性壤土(俗称犟黄泥)中,其质地细腻,干香可口,易酥不糊,营养丰富,...
大马士革玫瑰(RosadamasceneMill.var.kazanlika)为蔷薇科蔷薇属灌木,花香纯正,是萃取玫瑰精油和加工玫瑰纯露的优良品种。玫瑰精油含有多种化学成分(如香茅醇、香叶醇等芳香物质、有机酸等有益美容的物质)[1],被认为是玫瑰精油的极品。大马士革玫瑰...
直播青菜是指在前茬作物收获后将青菜种子直接播种的一种轻简栽培方式,省去了育苗移栽程序。直播青菜具有省工、节本、高产、高效等特点,在国外应用较为广泛,但在我国应用不多。在长江流域青菜生产区,由于一直采取育苗移栽的方式,对直播青菜栽培技术研究...
长江流域及其以南地区,水资源相对丰富,许多地方适宜发展水生蔬菜种植,各地也有较多的地方水生蔬菜品种资源。慈姑作为水生蔬菜的一种,近年来越来越受到消费者的欢迎。长江中下游地区慈姑种植面积较广,生产上慈姑主要采用育苗移栽,对育苗环境和育苗技...
野生植物,是指原生地天然生长的植物。野生观赏植物,从广义上说是具有观赏价值,可在园林中进行应用的野生植物,其观赏特性包括观花、观叶、观果等。野生植物资源是一个地区自然资源的重要组成部分,系统地开展野生植物资源调查,有助于了解一个地区植物资源的基...

葡萄为温带水果,为葡萄科葡萄属木质藤本植物。葡萄种植过程中易发生多种病虫害,严重制约了葡萄产量和品质的提升。因此,探究葡萄病虫害绿色防控措施对促进葡萄产业绿色健康发展有重要意义。...
黄金梨(HuangjinPyrusL.)是韩国园艺场罗洲支场1967年用新高梨廿世纪梨培育成的中晚熟品种,1981年命名[1],1984年定名。1997年引入我国山东烟台。该品种为梨中珍品,外观诱人、品质优良、较耐贮运,是目前世界公认的高档优质梨新品种,具有较高的栽培推广...