��ѧ����


��������
�������о����塿�ջ���Chrysanthemum×morifoliumRamat.�����й���ͳ�������������ǵ�ϲ���������緶Χ�ڣ��ջ����г�����Ҫ�����л�֮һ��Ҳ����Ҫ���軨���ر����ܡ��ջ���Դ���й����Ǿ�������Ұ������Ȼ�ӽ��پ��˹�ѡ���γɵģ�����Ҫ�ױ�Ϊë���գ�C.vestitum Hemsl.����Ұ�գ�C.indicum L.�����ϻ�Ұ�գ�C.zawadskii Herb.���ȣ�����������Ȼ�ӽ����˹�ѡ��������������˴��������±��죬���зḻ���Ŵ������ԡ��ջ��ı�����״����������״�����켫��ḻ�����伫���ܻ������ص�Ӱ�죬�������Ծջ�������״���о�ʼ�ռ�Ϊ���ѡ��Ծջ���Ҫ������״�Ļ����ͽ��н�����Ѱ����Ŀ����״��صķ��ӱ�ǣ���Ϊ����������״���Ŵ�ѧ�о���Ʒ�ּ�����Ʒ�ֱ����ͷ��ӱ�Ǹ������ֵ춨��Ҫ��������ǰ���о���չ�����ӱ�Ǽ����ѳɹ�Ӧ���ھջ�����Դ�о���Ʒ�ּ������Ŵ������Է������������ŷ��ӱ�Ǽ����ķ�չ������ȫ������ķ��ӱ�Ǻͺ��ʵķ���Ⱥ��Ѱ��������ǡ���չ�Ŵ�������ͼ��Ŀǰֲ��������״�о�����Ҫ����֮һ���ջ����ڽ������Է�ֳ��������߶��Ӻϣ��Ҵ��ڽ���˥���������ԣ��ջ����Ŵ�ͼ����һֱ�Ǹ����⣬�ڴ˷�����о������ո�������µ����õر��պ�����С�յ��ӽ� F1��Ⱥ�壬ͨ�����ŷ��������bulked segregant analysis��BSA����Ѱ�ҵ���ջ�������������ص� RAPD ��ǡ�Zhang �ȸ���“˫-�ٲ⽻”��ͼ���ԣ����������߶��Ӻϵľջ�Ʒ����Ϊ�ױ��������ӽ� F1����Ϊ��ͼȺ�壬ʹ��RAPD��ISSR��AFLP �� SRAP �ȷ��ӱ�dzɹ�������˫���Ŵ�����ͼ�ף���ɸѡ����ջ������ںͿ�����������ص� SRAP ��ǣ����Ի�������״�����Ȼ�����״������������״λ�㣨quantitative trait loci��QTL����λ�����ܲ�������ͼ�Ѿ��ھջ��н������������DZ���ܶȽϵ͡�����Ⱥ����������Լ����Ӧ��DZ���������о�����㡿�����������ֳƹ�����ͼ��������ƽ����ͼ�����㷺���������Ŵ�ѧ�о�����һ���о�����������״���Ŵ�ѧ��������������ȻȺ��Ϊ�о������Գ���������������ĵ�λ����λ�㣩��������ƽ�⣨linkage disequilibrium��LD��Ϊ��������Ŀ�������״�Ķ����������λ�㣩�Ķ�̬�Խ��������������ֱ�Ӽ���������ͱ�����������Ҿ����ض����ܵĻ���λ�����λ�㡣�봫ͳ��������ͼ��ȣ����������������ŵ㣺����ȻȺ��Ϊ���ϣ����蹹����ͼȺ�壻����ͬʱ���ͬһ��λ�Ķ����λ����������ͼֻ���漰�����ױ���2 ����λ���㷺���Ŵ����Ͽ�ͬʱ��������״�Ĵ���� QTL �Ĺ���λ�㼰���λ���죬����������ͼ�����ױ���Χ��Ⱥ�������������ơ���������������ͼ��չ�ջ�����״ QTL ��λ���Ѷȴ����ڳ�����˿��Կ������ù����������ص㣬��ַ����й��ջ�������Դ�ḻ���������ƣ�����ȻȺ��Ϊ������չ������������������Ҫ������״�����ķ��ӱ�ǣ��Ӷ�Ϊ�ջ����Ӹ������ַ��������Ĺؼ����⡿���о��ڷ���Ⱥ��ṹ�Ļ����ϣ��ù��������� 58 �����Ʒ�ֵ� 18 ��������״�� SRAP ���ӱ�ǽ��й�����ϣ����������������״������ı��λ�㣬Ϊ�ջ����ֹ��������뿪չ��Ʒ�ּ����ṩһЩ�м�ֵ�IJο����ϡ�
����1 �����뷽��
����1.1 ���Բ���
�������о����в��Ͼ����Ա�����ҵ��ѧ�ջ���Դ�ԡ��� 2008—2010 ���� 3 ��Ծջ���Դ���� 800������Ʒ�ֱ�����״���й۲�����Ļ����ϣ��Ծ����ܸ��Ǵ��Ʒ�ֳ��ֵ�������״����Ϊԭ������Դ����ѡ����д����Ե� 58 �����Ʒ�֣����� 5 �����ͣ�30 �����ͣ����� 50 ���й�Ʒ�ֺ� 8���ձ�Ʒ�֣���Ϊ��ֱ����Ե��ϵ����ȻȺ�壨��1����
��������Ʒ���� 5 �����չ�������Ǥ�己ֳ��6—7 �¶�ֲ��ֱ�� 30 cm �����У����ն����յ����༼��������������
����1.2 ����
����1.2.1 ������״�IJⶨ����� ���о��вⶨ 18 ��������״��������������״ 11 ����������С�����곤�ȡ�������ȡ��������ȡ����ߡ������ֶȡ���״С������Ͳ״С������Ͳ״�����ȡ�Ͳ״����ֱ�������д�С����Ҷ����״ 4 ����Ҷ�����ȡ�ҶƬ���ȡ�ҶƬ���ȡ�Ҷ��������״ 2 �����ڼ䳤�ȡ����֣���������״ 1 ����ֲ��߶ȣ�����״�IJⶨ�������������ȵ�������ÿ��Ʒ�ֲ��� 10 �����꣬ȡƽ��ֵ���������״��Ʒ���ں�Ʒ�ּ�ı���ϵ�������㹫ʽΪ��CV=S/X��S={[EX2-(EX)2/N]/N-1}1/2���У�CV Ϊ����ϵ����S Ϊ���X Ϊ������ֵ��
����1.2.2 DNA ��ȡ ���ȡֲ����Ҷ���ø��� CTAB ����ȡ������ DNA���� 1%��֬��������Ӿ������ֹ��ȼƲⶨ DNA ������Ũ�ȣ�����Ũ��ϡ���� 50ng·μL-1����Ʒ��-20���±��汸�á�
����1.2.3 SRAP ���� ѡ���������� 13 ������������ 10������ 130 �� SRAP ������������ɸѡ��SRAP ���������ŷɵȵķ�����������ϵ�Ż�������ɸѡ�Թ���Ʒ�ֽ�����������������ͬǨ���ʵ�����Ƭ�Σ����ն����Ʒ������м�¼�����д��ļ�Ϊ 1�����ļ�Ϊ 0������¼�������ȶ������������������γ� 0/1���������������Ŵ������Է������ö�̬����Ϣ����Polymorphism Information Content��PIC��ָ�ꡣ���� Anderson �ȵļ��㷽������� i �� PIC ֵ���㹫ʽΪ����1��
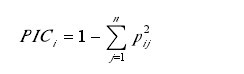
����
�������У�pij��ʾ��� i �� j �ִ��ͳ��ֵ�Ƶ�ʣ���� i�Ĵ������� 1 �� n��PIC ֵ�ķ�ΧΪ 0—1��0 ��ʾ��̬�ԣ�1 ��ʾ���зdz��ߵĶ�̬�ԡ�
����1.2.4 �� �� ͳ �� �� �� �� Ⱥ �� �� �� �� �� ��STRUCTURE�����Ծջ�Ʒ��Ⱥ��ṹ���л�����ѧģ�͵���Ⱥ���֡����趨Ⱥ����Ŀ��K��Ϊ 2—9���� MCMC��Markov chain Monte Carlo����ʼʱ�IJ�����������Length of burn-in period����Ϊ 10 000 �Σ��ٽ������������ MCMC ��Ϊ 100 000 �Σ�Ȼ������ÿ�� K ֵ��Ӧ�� lnP��D��ֵ��������Ȼֵ����ԭ��ѡ����ʵ�Kֵ��������õ�ÿ��Ʒ����Ӧ��Qֵ����i Ʒ������������Դ�ڵ� k Ⱥ��ĸ��ʣ�����������Ӧ�� TASSEL ���� GLM��General Linear Model��һ������ģ�ͣ����������ջ�Ʒ�ֵ� Q ֵ��ΪЭ��������18��������״�ı�����״���ݷֱ��SRAP��DZ�����һ���лع������ȷ�ϱ�����״����λ�㲢����λ��Ա��ͱ���Ľ����ʡ�GLM �ع鷽����:��2��
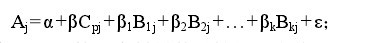
�������У�Aj�ǵ� j ������������״�ⶨֵ��β ��Ⱥ���λ�����λ�����ƽ��ЧӦ��Cpj�ǵ� j ���ϵ� P ��λ������ֵ�ָʾ������B1j—Bkj�ǵ� j �����ϻ��������Դ�ڵ� l—k Ⱥ��ĸ��� Q ֵ��β1—βk����Ⱥ���λ�����λ�����ƽ��ЧӦ��ε �Dzв
����2 ���
����2.1 ������״�ı������
�������о���Ӧ�ñ���ϵ������ʾ�۲��������״��Ʒ���ں�Ʒ�ּ����ɢ�Գ̶ȡ�Ʒ���ڱ���ϵ����ӳ����״��ͬһƷ���ڵ��ȶ��̶ȣ�Ʒ�ּ����ϵ����ӳ����״�ڲ�ͬƷ�ּ����ɢ������ɸ���״����ϵ������������ 2����֪��18 ��������״��Ʒ�����ȶ��ԽϺã���������Ʒ���ڱ���ϵ����ߣ�Ϊ 0.22�������δ���� 0.2����ֵΪ 0.13��������״��Ʒ�ּ����ϵ���ϴ�Ҷ������������״��Ʒ�ּ�IJ��첻�绨����״���ԡ���ˣ��ڹ��������п��ܸ������ڲ�ͬ�����ͼ��ҵ��뻨����״�����ı�ǡ�
����
����2.2 SRAP ������ 130 �� SRAP ���������ɸѡ�� 19 �����������ȶ���������϶���ѡ��� 58 �����Ʒ�ֽ�������������������Ƭ�� 283 �������ж�̬��Ƭ��Ϊ 225������̬��λ��ռ 80%��ÿ��������ϼ���λ������Ϊ 7—23 ����ƽ��Ϊ 11.84 ������ 3������̬����Ϣ�� PIC ֵ�� 0.76—0.94��ƽ��Ϊ 0.87��Ϊ�߶ȶ�̬����λ��˵��ѡ��Ĵ��Ʒ��Ⱥ����Ŵ�����dz����зḻ���Ŵ������ԣ����������ڹ��������з����������״������λ�㡣ͼ 1 ������ Me1/Em6 �ڹ���Ʒ������������� 6%�۱�ϩ����������Ӿͼ��
����2.3 ����Ʒ�ֵ�Ⱥ��ṹ����
�������� SRAP ��ǣ�������ѧģ�͵�Ⱥ��ṹ���������������ĵ�λ����Ƶ������������ K=5��������Hardy-Weinberg ƽ�����Ⱥ��ĿΪ 5��ʱ����ģ�͵ĺ������[lnP(D)]���ͼ 2�����ɴ��ж� 58 �����Ʒ��Ⱥ��ɱ���Ϊ 5 ����Ⱥ�����������Ʋ�� K ֵ���ƹ���Ʒ��Ⱥ��ṹͼ��ͼ 3������Ⱥ��ṹͼ���Կ�����58 �����Ʒ���д��� 5 ����Ⱥ����������Ⱥ������ѧ���壬���־ջ�Ʒ�ֻ�������ͺ͵�����أ��ɻ�����ʶ��Ϊƽ���ࡢ�ܰ��ࡢ�����ࡢ�������ձ�Ʒ����Ⱥ������Ⱥ�������ԵIJ��죬����һ�������ԣ���˵������Ⱥ����������Ե�Ⱥ��ṹ���ձ�Ʒ����Ⱥ��������ǿ�����������Ⱥ��ƽ���ࡢ�ܰ��ࡢ�ձ�Ʒ����Ⱥ�����������ߣ��ܰ�����Ⱥ��ƽ������Ⱥ��һ�����ԡ�
����Ϊ����Ⱥ��ṹ�Ĵ���ͨ��Ӱ��λ�� LD ����Ӱ�����������ȷ�ԣ����о�����Ʒ�� Q ֵ����Ʒ�ָ���������Ⱥ�ĸ��ʣ���ΪЭ�������� SRAP ����������״����Ļع�����С�
����2.4 SRAP ����������״�Ļع��������
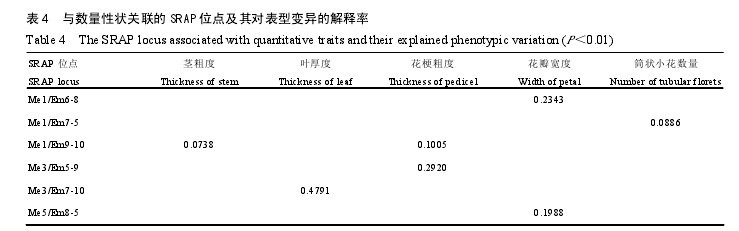
������58�����Ʒ�ָ�Ʒ����Ӧ��Qֵ��ΪЭ��������18��������״�ı��ͱ����SRAP��DZ�����лع������Ѱ������״������ı�Ǽ����λ���졣�����ʾ���������� 225 �� SRAP ���λ���У�����6 �� SRAP ���λ���� 5 ��������״�����ֶȡ�Ҷ��ȡ������ֶȡ�������Ⱥ�Ͳ״С���������� P��0.01ˮƽ����ء��뻨����״�������ֶȡ�������Ⱥ�Ͳ״С�����������λ�㹲 5 �����뾥�������ֶȣ���Ҷ����״��Ҷ��ȣ����λ��� 1 ��������λ��Me1/Em9-10 ͬʱ�뾥�ֶȡ������ֶ���ء���λ��Ա��ͱ���Ľ������� 0.0738—0.4791�����������0.4791���� SRAP λ���� Me3/Em7-10����Ҷ�����أ�����������С��0.0738����λ���� Me1/Em9-10���뾥�ֶ���أ��� 4��������4��
����
����3 ����
����
����3.1 ������ѧģ�͵ľջ�Ⱥ��ṹ����
�������� SRAP ��ǣ�ʹ�� STRUCTURE �����Թ��Ծջ����ʽ��еĻ�����ѧģ�͵���Ⱥ���֣���������Ŵ������ SRAP ��� UPGMA ��������δ�г�������һ�¡������й����Ʒ�ֿɷ�Ϊƽ���ࡢ�ܰ��ࡢ���������� 4 ����Ⱥ�ṹ������Ⱥ�ṹ���ڹ㷺�Ļ��������װ���Ʒ�ֲ�δ�����γ���Ⱥ�����Ƿ�ɢ��ƽ����ܰ�����Ⱥ�ڣ�˵���װ���Ʒ�����ݻ���ϵ���Ǵ���ƽ����ܰ���֮��Ĺ���״̬����������������IJ��ѳװ���Ϊ�������͵Ĺ۵���һ�¡�������ѡ�ձ�Ʒ�ֶ�Ϊƽ����ܰ��࣬����δ���й���յ�ƽ������ܰ����ΪһȺ�����ǵ�����Ϊ 1 ����Ⱥ��˵���������ڵ�����������أ���������Դ���ݻ��������й�Ʒ�������һ���IJ��죻���������ȺҲ�����ڻ������������ձ�Ʒ�ֺ��й�Ʒ���ھջ�����������ʷ�ϴ��ڹ㷺������������
�������о����� SRAP ������ѧģ�͵ľջ�Ⱥ��ṹ������ǰ��������̬ѧ���ݻ��������ӱ�ǻ����Ŵ�����ľ�����õ��Ľ������һ�£�֤�������Ч�Ծջ�Ⱥ��ṹ�����жϺͻ��֡���Ȼ����Ŵ�����ľ������������ѧģ��������Ⱥ��ṹ���ų���Ⱥ���ֵ���Ϊ���أ�����ȷ��ȷ����Ⱥ��Ŀ�����ҿɸ������ع۲����Ⱥ��Ļ����������
����3.2 �������״�����ķ��ӱ��λ��
������ֲ�����״ν���ȫ����������������� Hansen�ȶ�Ұ���������ϰ�Ե��о������� 17 ��������������� 440 ��ȫ�����鷶Χ�ڵ� AFLP ����У���2 ������Ƴ�ǰ�Ƿ���Ҫ������ B �������Ź�����
����Kraakman �ȶ�236��AFLP��Ǻʹ�����Ʒ�ֵIJ����������ȶ��Խ����˹����������ֱ��� 8 ���� 5�� AFLP ���������Ͳ����ȶ�����������ξ���Ӧ��SRAP��EST-SSR���ӱ�Ƕ�ľ��Ʒ��ũ����״���й����������� 1471 �� SRAP ���λ���й���73 ��λ���� 21 ��ũ����״����������� 46 �� SRAP���λ��ͬʱ������״��������� 993 �� EST-SSR���λ������ 20 ��λ���� 20 ����״������ء����о��� 225 �� SRAP ���λ���й��� 6 ��λ������Ʒ�� 5 ��������״��أ�P��0.01������������״������� SRAP λ����٣����������� SRAP�������������ɡ��ϸ������Ͻ���ȫ���������������Ҫ��ǧ����ı���Լ������ܶ������Ե��ϵ�ĸ��塣��ʹ�ǻ������С�����Ͻ棬Ҳ��Ҫ��� 2 000�����ӱ�ǡ����о��� 18 ��������״��ֻ�ҵ� 5���б����֮��������״�����˱�����������⣬����Ⱥ��Ĵ�СҲ��Ӱ������֮һ�����о�ѡ��ľջ�Ʒ��Ⱥ���С��ÿ�����͵Ĵ�����Ʒ��Լ 10 ����ÿ�����͵Ĵ�����Ʒ��Լ 2 ��������ڱ�����״�����Է����п��ܲ�δ��ȫ���Ǹ�����״��ȫ�����죬SRAP��Ǽ��ı���λ��Ҳʮ�����ޡ���˱��о�ֻ������������ϵ�ȫ����������������ڽ�һ���Ĺ��������о��л������ӱ���ܶȣ�����Ⱥ���ģ��
�������о����ҵ��Ĺ���λ���У��뻨����״������� SRAP λ�㹲 5 �������뾥����Ҷ����״���λ��� 1 ����������뻨����״��Ʒ�ּ���ڽϴ�̶ȱ����йأ������ڹ�����ǵķ��֡�λ�� Me1/Em9-10ͬʱ�뾥�ֶȡ������ֶ���ء��뻨���ֶ���ص�λ���� 2 �����ֱ��� Me1/Em9-10 �� Me3/Em5-9���뻨�������ص�λ���� 2 �����ֱ��� Me1/Em6-8 ��Me5/Em8-5�����й�����У���Ϊ���������ߵ�����֮һ�������ڻ�����ϸ��ɻ�ͷ���ۣ���Ҫ�������ķ��������������о������뻨���ֶ��������λ�� 2������Ϊ���������������׳Ʒ���ṩ�����ݡ�
����ĿǰӦ����ֲ����������ķ��ӱ����Ҫ�� SSR�� SNP ��ǣ����о�Ӧ�õ��� SRAP ���ӱ�ǡ�SRAP���λ��Ķ�̬�Ա� SSR �� EST-SSR �ߣ��������RAPD �� AFLP ���ŵ㣬����Ķ�̬�Կ������ڰ������ں��ӡ������Ӻͼ��������Ʒ�ּ�ı�������������м�㡢�ȶ�����ͨ�����ص㡣���� SRAP Ӧ���ڹ�������Ҳ����ȱ�㣺SRAP ��������Ա�ǣ��������ִ������Ժ��Ӻ����ԣ�����ֱ�ӻ��λ���С��Ϣ����Ҫ���� marker Ƭ�ι��⣬������λ��ͳ����������Ժ�ľջ��������������� SSR����EST-SSR����SNP �ȹ����Է��ӱ�����о������ƣ����ջ�����δ�������������ǡ�����Ŀǰ�ջ���������о�ˮƽ�����ø��ַ����������ջ��� SSR �����Ŀǰ���в��ҿ��е��о�����
����3.3 �ջ��Ĺ���������������ͼ
������ϵ������ͼ��Family-based mapping��FBLmapping������������ǽ���ֲ����Ҫ������״�����͵���Ҫ���������о����ù��������ķ���ͨ���ع�������ջ�Ʒ�ֱ��ͱ���� SRAP λ���λ����Ĺ����ԡ���ȱ�����ܶȾջ������Ŵ�����ͼ������£�������ȻȺ�忪չ QTL ������������һ�ֽϼ������Ч�ķ������о����������ù����������ķ��ӱ��λ�㣬����붨λ���Ŵ�����ͼ�ϵ� QTL λ����һ�¡�������������ȱ���Dz��ܹ��� QTL �ľ���λ�ü�����Ի���λ��ЧӦ���������������������ͼ���ϣ������ٽ�����������״�������������ڸ�ȷ������ȷ�ؼ��������ƾջ���Ҫ����״�� QTLλ�㡣
����4 ����
�������о����״��ھջ�Ʒ���п�չ������ȻȺ��Ĺ���������ʹ��ɸѡ���� 19 �� SRAP ������϶� 58�����ʹ��Ʒ�ֽ��ж�λ��ɨ��������ڶԹ��Բ��Ͻ��л�����ѧģ�͵�Ⱥ��ṹ�����Ļ����ϣ����ջ�18 ����Ҫ������״���ݷֱ�� SRAP ��DZ�����лع�������� 5 ��������״�� 6 �� SRAP ���λ���������������뾥�ֶȡ�Ҷ��ȡ������ֶȡ�������Ⱥ�Ͳ״С�����������ı��λ�㡣���о�֤��������ѧģ�͵�Ⱥ��ṹ��������Ч�Ծջ�Ⱥ��ṹ�����жϺͻ��֣�ҲΪ���Ʒ�ַ����ṩ���µ��ֶΣ���������Ϊ�ջ����ӵ�������״�����ṩ���µ�;�����ܹ���Ч���ҵ���ջ�������״������ SRAP ��ǡ��о������Ϊ�ջ���Ʒ��ѡ�������ӱ�Ǹ������ֺͿ���������״����ػ���Ŀ�¡�춨������
����References
����
����[1] ��˼��. �й��ջ������绨����. �ӱ��Ƽ�ʦ��ѧԺѧ��, 2004,18(2): 1-7.Dai S L. Chinese florist’s chrysanthemum and the world flowerhorticulture. Journal of Hebei Normal University of Science �}Technology, 2004, 18(2): 1-7. (in Chinese)
����[2] ����, ��˼��. �ջ�������Դ�о���չ. ֲ��ѧ��, 2009, 44(5):526-535.

�ٺ�(Liliumspp.)�����������л�֮һ�����кܸߵĹ��ͼ�ֵ��һֱ��������ϲ��������Ҫ����Ʒ���ܺ����̻�ֲ���Robb�״����������ٺ��۾���������֯���������ڰٺ��ϵ�Ӧ����ȡ�������ŵĽ�չ�����ٺ�����࣬�����죬Ŀǰ�ܶ�Ʒ����δ�γɹ�ģ����...
ֲ�������������������ܵ����ַ��������ӵ�в�ȣ������¶���Ӱ��ֲ����������Ҫ���ӡ���������ȫ�����²������ߣ���ʢ�������¶ȸ���ֲ�������������¶ȣ�����������ֲ������������¶ȣ���ֲ�������������������������������в����ˣ��о�ֲ���ڸ���в��...
��Ƥ����������Ʒ�����������˹�ɸѡ����
1������Ŀ�������������������ˮƽ�����,���ǵĽ�����ʶ������ǿ,��ʳ����ת����ʳƷ��ũҩ������������Ⱦ������ʹ���Ƕ�ʳƷ��ȫ�ĵ��Dz��ϼӾ�,��������ϣ��ʳ����������Ȼ����...
�������ֳƵ���,��������,���仨����ʵ��������������������һ�ֶ���������ֲ����곣�̲ݱ�,�����֦�����۾�,Ҷ�̲��ʺ�,���⡣��ϵ����,�ɱ�ƽ��״,�ӽڲ���������״����Ҹ��,�´�,����10~20�䡣��ɫ�������а�ɫ����ɫ����ɫ,���ڻ��л�ɫ�ȵȡ�...
��ͬ�ɺ�в�ȶԺ컨������������������Ӱ��
ˮ����ֲ�������ı�Ҫ����,����ˮ����Ӱ��ֲ������ˮ��״������Ҫ����.�������ֲ���ڸɺ�в�����������������仯���о��϶�,�����о�����:�ɺ�в�Ȼᵼ��ֲ��ϸ���ڻ��������ɻ�����������IJ�ƽ��,�Ӷ����ֻ��������ɻ��Ĵ�������,���ɴ������ͼӾ���ϸ��...
�ɺ�в���´�����ҶƬ��������ָ�꼰������ø
������(Dahliapinnata)ԭ����ī���硢Σ�����������ױ���һ�����Ǿտƴ��������Ķ���������ݱ����ܣ����ڳ���ɫ����������м��ߵĹ��ͼ�ֵ��Ŀǰ�Դ��������о���Ҫ���������༼������ѿ�ֻ���Ʒ����Դ����ࡢ��֯����������ͻ����Ͳ��溦���εȷ��棬��...
Ұ��ֲ��,��ָԭ������Ȼ������ֲ�Ұ������ֲ��,�ӹ�����˵�Ǿ��й��ͼ�ֵ,�������н���Ӧ�õ�Ұ��ֲ��,������������ۻ�����Ҷ���۹��ȡ�Ұ��ֲ����Դ��һ��������Ȼ��Դ����Ҫ��ɲ���,ϵͳ�ؿ�չҰ��ֲ����Դ����,�������˽�һ������ֲ����Դ�Ļ�...
�����ѣ�IrisensataThunb��������������������ݱ�����״���̴֣��������ϸ���Σ����β���β��������ֱ���ɴ�15cm���������ڴ��꣬�л�ɫ���ʺ�ɫ����ɫ����ɫ�ȣ�������ɫ����ɫ����ɫ���ߵ�����ƣ�ɫ�ʶ�䣬��Ʒ�ִ�����֮...
��������(Syringareticulatavar.mandshu-rica),�׳Ʊ����ӡ��ɻ�����,ľϬ�ƶ�������Ҷ���ľ��С��ľ,��4~15m����Ҫ�ֲ��ں������Ĵ�С�˰��롢���ֶ���ɽ�������������������������е���,���ʡ�����˹���ձ�Ҳ�зֲ�����������ϲ��ůʪ������,���Ϻ�,��...
ֲ�ﻯ��������ֲ��ͨ�����ܡ��ӷ�������ֽ��ϵ���ڵȷ�ʽ���ͷŻ�ѧ���ʣ��Ӷ�����Χֲ�����ֱ�ӻ��ӵġ��к������������á�ֲ�������ֻ�ѧ���ö�����������ɭ�ָ�����ֲ�ﱣ����������εȷ������Ź㷺��Ӱ�졣Ŀǰ��ֲ�ﻯ��������...