园艺学论文

红瑞木(SwidaalbaOpiz)是山茱萸科梾木属(SwideOpiz)灌木,产自黑龙江、吉林、辽宁、内蒙古、河北、陕西、甘肃、青海、山东、江苏、江西等省区[1]。金叶红瑞木,其叶春季、秋季金黄,夏季变绿,叶色随季节变化明显,比普通红瑞木相具有更高的观赏价值。目前,关于金叶红瑞木的研究大部分集中在快繁技术[2,3]、抗逆性等方面[4-6],对其光合生理生态的研究未见报道。本文通过对金叶红瑞木的光合、水分生理系统的研究,探讨其适宜的生长环境条件并为其在园林中植物造景设计提供理论依据。
1 材料与方法
1.1 试验材料
试验材料选择中铁大桥局集团园林环境工程有限公司合肥市科研基地露地栽植二年的金叶红瑞木(SwidealbaOpiz'Aurea)。
1.2 试验仪器与测定方法
在晴朗天气的 2014 年 8 月 5 日,采用美国 LI-COR公司生产的 Li-6400 便携式光合仪,选择生长健康植株中部向阳面的叶片,从 6:00~17:00 时每隔 1 小时测定 1 次净光合速率(Pn,μmolphotons·m-2·s-1),同时记录气孔导度(Gs,μmolphotons·m-2·s-1)、胞间 CO2浓度(Ci,μmolCO2·mol-1air)、 蒸 腾 速 率(Tr,mmolH2O·m-2·s-1)、叶面水汽压亏缺(VpdL,Kpa)、气温(Ta, ℃)、 叶 温(Tl, ℃)、 空 气 CO2浓 度(Ca,μmolCO2·mol-1air)、空气相对湿度(RH,%)、光合有效辐射(PAR,μmolphotons·m-2·s-1)等生理生态学指标进行测定。选择该种 3 个植株,每株测定 1 次,共重复 3 次。
为了避免不同叶片对光照强度反应的差异,每次取同一叶片相同部位进行测量,取测量平均值进行各项分析。光响应曲线的测定利用 LED 红蓝光源,使光合有效辐射(PAR)从最大值(2000μmolphotons·m-2·s-1)逐渐降低至最小值(0μmolphotons·m-2·s-1),测定在不同光合有效辐射下的光合速率,绘制金叶红瑞木的光响应曲线。光强的梯度划分为 :2000、1800、1500、1300、1100、1000、900、800、700、500、300、200、150、100、80、20、0μmol·m-2·s-1。
1.3 数据处理
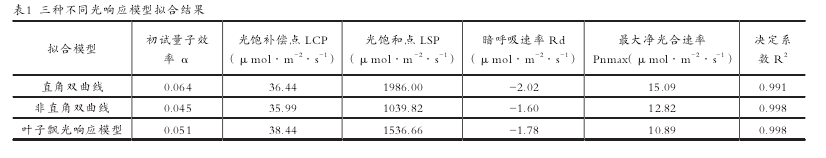
利用 SPSS20.0 统计软件,分别用直角双曲线模型[7]、非直角双曲线模型[7]和叶子飘光响应模型[8]对金叶红瑞木的光响应曲线进行拟合比较,并估算植物光响应特征参数,如光补偿点(LCP,lightcompensationpoint)、光饱和点(LSP,lightsaturationpoint)、最大净光合速率(Pnmax)、暗呼吸速率(Rd,darkrespirationrate)。用直线方程拟合金叶红瑞木弱光条件下(PAR ≤ 200μmol·m-2·s-1)的光响应数据,所得直线方程的斜率即为表观量子效率(AQY,apparentquantaefficiency)。使用 Excel 对原始数据处理并制图,使用 SPSS20.0 对数据进行 Bivariate 相关性分析。
2 结果与分析
2.1 金叶红瑞木的光合生理特点
2.1.1 光补偿点和光饱和点
金叶红瑞木光补偿点为 19-80μmol·m-2·s-1,光饱合点为 1500-1800μmol·m-2·s-1(图 1)。运用 3 种不同的光响应模型对金叶红瑞木的光响应曲线进行拟合得出光补偿点和光饱和点(表 1)。与实测值相比直角双曲线拟合的光饱和点的值高于实测值所在范围,而非直角双曲线的拟合值远低于实测值所在范围。叶子飘光相应模型所拟合的光饱和点的值与实测值较为接近。用 3 种模型拟合的光补偿点的值均在实测值范围内且数值大小接近。因此,运用叶子飘光响应模型拟合金叶红瑞木的光饱和点和光补偿点所得结果较为理想,分别为 1536.66μmol·m-2·s-1和 38.44μmol·m-2·s-1。【1-2】
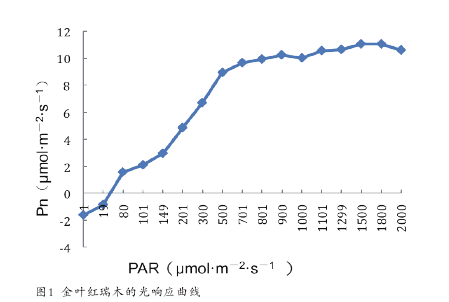
2.1.2 耐阴性
用直线方程拟合弱光强条件下(PAR≤200μmol·m-2·s-1)的光响应数据,所得直线方程的斜率即为表观量子效率(AQY)[9]。根据拟合直线方程 Y=0.0314X-1.3629,金叶红瑞木表光量子效率为 0.0314(图 2)。自然条件下一般植物的表观量子效率范围为 0.03-0.05[10],说明拟合的数据是合理的。表观量子效率反映了植物光合作用的光能利用率,尤其是对弱光的利用能力,AQY 值高,说明其叶片光能转化效率高[11]。金叶红瑞木表观量子效率较低,说明其对弱光的利用能力较低,耐阴能力差。【3】
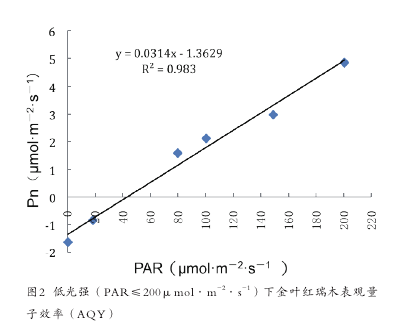
金叶红瑞木的光饱和点为 1536.66μmol·m-2·s-1,其 光 饱 和 点 与 阳 性 树 种 马 尾 松(Pinusmassoniana,1382.76μmol·m-2·s-1)比较接近,高于中性植物杉木(Cunninghamialanceolata,751.5μmol·m-2·s-1) 和 青檀(Pteroceltistatarinowii,881.76μmol·m-2·s-1)[12],因此金叶红瑞木属于阳性植物。
植物光补偿点是体现植物在弱光条件下光合作用能力的重要指标[13],光补偿点低说明植物利用弱光能力强,有利于有机物质的积累。光补偿点低且光饱和点相应也低的植物具有很强的耐阴性。光补偿点低,光饱和点较高的植物,能适应多种光照环境。光补偿点较高,而光饱和点较低的植物,应栽植于侧方遮阴或部分时段荫蔽的环境。光饱和点和光补偿点均较高的植物则为喜光的阳生植物[14]。
金叶红瑞木的光补偿点较高(38.44μmol·m-2·s-1),光饱和点也高(1536.66μmol·m-2·s-1)而且表观量子效率较低(0.0314),因此金叶红瑞木属于不耐阴的阳性树种。
2.1.3 净光合速率的日变化
夏季气温较高,金叶红瑞木的净光合速率(Pn)日变化成双峰曲线型(图 3),在 10 :00 时和 14:00 时分别出现 峰 值 16.16μmol·m-2·s-1和 12.10μmol·m-2·s-1,下午的高峰值为上午高峰值的 74.9%。在两峰之间即 13:00时净光合速率下降到最大值的 56.4%,形成低谷,表现出明显的光合“午睡”现象。图中 13:00 时左右光合有效辐射略有下降,原因是多云天气造成的。【4】
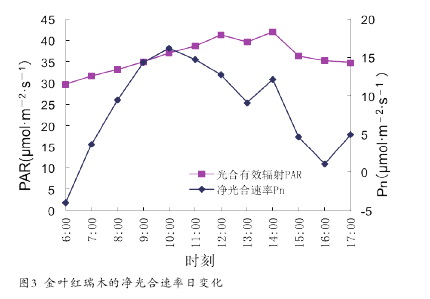
2.2 净光合速率日变化的影响因子
本文在多个影响金叶红瑞木净光合速率和光能利用率的因子中除去了胞间 CO2浓度(Ci)这个影响因子。因为现代化的光合作用测定仪器计算的Ci值仅仅在叶片上全部气孔的开放或关闭行为均匀一致的前提下才是正确的[15]。
但是实际日变化的测量中气孔不可能总是全开全闭,所以得出的 Ci 的值不具备参考价值。
金叶红瑞木净光合速率与各影响因子测定数据见表 2。
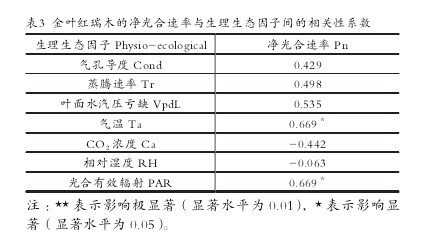
使用 SPSS 软件得出的相关性分析结果(表 3)表明 :生理生态因子对金叶红瑞木净光合速率的影响由大到小依次为PAR=Ta>VpdL>Tr>Ca>Cond>RH。光合有效辐射和气温对净光合速率的影响最大,其次叶面水汽压亏缺。各影响因子中 Ca、RH 与净光合速率呈负相关,其余因子与其呈正相关。【5-6】
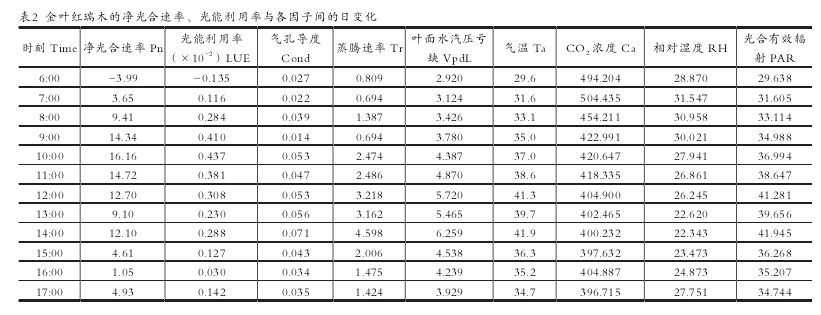
2.3 光能利用率及其影响因子的分析
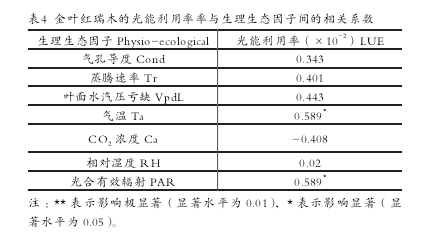
光能利用率(LUE)是指植物光合作用积累的有机物所含的能量占照射在单位地面上日光能量的比率。金叶红瑞木的光能利用率日变化见表 2。使用 SPSS 软件得出的相关性分析结果表明(表 4),光合有效辐射和气温对其光能利用率影响显着,其次为叶面水气压亏缺。各影响因子中 Ca与净光合速率呈负相关,其余因子与其呈正相关。【7】
3 结论与讨论
本文所得的结论有 :(1)金叶红瑞木的光饱和点为1536.66μmol·m-2·s-1,光补偿点为38.44μmol·m-2·s-1,表观量子效率为 0.0314,光补偿点较高,光饱和点较高且表观量子效率较低,因此属于不耐阴的阳性树种,适合种植于光照充足的环境。
(2)金叶红瑞木在晴朗的夏天,叶片的净光合速率日变化呈双峰曲线型,即 10 :00 时和 14 :00 时分别为一高峰 ;在 13 :00 是净光合速率为低谷,表现出明显的光合“午睡”现象。
(3)影响金叶红瑞木净光合速率的生理生态因子由大到小依次为 PAR=Ta>VpdL>Tr>Ca>Cond>RH。光合有效辐射和气温对净光合速率的影响最大,其次叶面水汽压亏缺。各影响因子中 Ca、RH 与净光合速率呈负相关,其余因子与其呈正相关。
(4)影响光能利用率最显着的因子是光合有效辐射和气温,其次为叶面水气压亏缺。各影响因子中 Ca 与净光合速率呈负相关,其余因子与其呈正相关。
前人对金叶红瑞木光合特性的研究较少。文慧娟曾对花叶红瑞木的光合特性进行研究得出其光合作用日变化呈双峰曲线且“午休”现象明显的结论,与本文金叶红瑞木光合作用日变化的结论基本一致。
夏季为了提高金叶红瑞木的净光合速率和光能利用率应适当增加光合有效辐射和环境温度,但是在炎热干燥的正午期间为缓解“午休”现象应注意降温、保湿,如采取叶面喷雾的措施以降低小环境的温度。作为城市“彩化”的园林植物金叶红瑞木适宜栽植于光照条件良好的环境中,也可以与其他草本和灌木相组合种植于稀疏的林地中,但不适宜栽植于郁闭度过高的林下或建筑物阴面。
参考文献
[1] 中国科学院中国植物志编辑委员会.中国植物志·第56卷[M].北京:科学出版社,1990.
[2] 张明丽.金叶红瑞木快繁技术研究[D].北京:北京林业大学,2004.
[3] 段新玲,任东岁.金叶红瑞木嫩枝扦插繁殖试验[J].林业科技1999(6):46-48.
[4] 种培芳,苏世平.4种金色叶树木对SO2胁迫的生理响应[J].生态学报,2013(15):4639-4648.
[5] 陈兴玲,等.三种彩色树对盐碱胁迫的生理响应[J].吉林林业科技,2009(5):10-14.
[6] 张树宝,等.电导法等测定欧美彩叶树抗寒性试验研究[J].北方园艺,2009(2):187-189.

红掌(Anthuriumandraeanum)属天南星科花烛属,为多年生附生常绿草本植物,原产于中美洲和南美洲热带雨林,是典型的热带花卉。在世界花卉贸易中,红掌的销量仅次于热带兰,是近年来国内外最为流行的名贵花卉之一。红掌的生长发育对高温敏感,特别是在...
引言【研究意义】菊花(ChrysanthemummorifoliumRamat.)是中国传统名花,深受人们的喜爱。在世界范围内,菊花是市场上主要的鲜切花之一,也是重要的盆花、地被花卉。菊花起源于中国,是菊属部分野生种天然杂交再经人工选育形成的,其主要亲本为毛华菊(C.v...
百合(Liliumspp.)是世界着名切花之一,具有很高的观赏价值,一直深受人们喜爱,是重要的商品花卉和园林绿化植物。自Robb首次离体培养百合鳞茎以来,组织培养技术在百合上的应用已取得了显着的进展,但百合种类繁多,类型各异,目前很多品种尚未形成规模化生...
植物在生长过程中往往受到各种非生物因子的胁迫,其中温度是影响植物生长的重要因子。近年来,全球气温不断升高,在盛夏由于温度高于植物生长的最适温度,甚至超过了植物生长的最高温度,对植物生长发育甚至生命都造成了严重威胁。因此,研究植物在高温胁迫...
1、育种目标近年来随着人们生活水平的提高,人们的健康意识不断增强,而食物中转基因食品、农药残留、环境污染等因素使人们对食品安全的担忧不断加剧,所以人们希望食到健康、天然、保...
凤仙花属(ImpatiensL.)植物是中国着名传统花卉之一,也是世界着名观赏花卉[1]。全球范围内最主要的分布区有5个,分别是:东亚区、部分东南亚地区、印度南部和斯里兰卡、热带非洲区和马达加斯加区[2-5]。该属是凤仙花科Balsaminaceae(仅有凤仙花属和水角属Hydro...
1前言大花蕙兰又名虎头兰、喜姆比兰和蝉兰,为兰科兰属植物。大花蕙兰叶长碧绿,花姿粗旷,是世界着名的兰花新星它具有国兰的幽香典雅,又有洋兰的丰富多彩,花朵大,花色多,深受人们的喜爱。我国虽然是大花蕙兰的原产地,又拥有丰富的种子资源,但仅是原...
干旱是一个长期存在的世界性难题。太原市水资源尤为匮乏,选育优良的抗旱种类应对缺水干旱现状,成为选育研究工作中急需解决的重要问题。因此,研究者纷纷开展了对地被植物抗旱性的研究[1-4]。笔者对5种观赏性较强的园林地被植物婆婆纳、紫花地...
随着经济的发展和人们对物质、精神文化需求的提高,草坪在园林绿化中的作用日趋重要。冷季型草坪草在我国许多北方城市的园林绿化中应用广泛,因其养护管理需要耗费大量的水资源,对干旱少雨的北方地区造成了很大的市政供水困难,水资源短缺成为北方地区限制草坪...
大马士革玫瑰(RosadamasceneMill.var.kazanlika)为蔷薇科蔷薇属灌木,花香纯正,是萃取玫瑰精油和加工玫瑰纯露的优良品种。玫瑰精油含有多种化学成分(如香茅醇、香叶醇等芳香物质、有机酸等有益美容的物质)[1],被认为是玫瑰精油的极品。大马士革玫瑰...