植物学论文


TEOSINTE BRANCHED1 / CYCLOIDEA / PRO-LIFERATING CELL FACTOR1( 简称 TCP) 家族是一类植物特有的转录因子家族。最早发现的家族成员是玉米 TB1( teosinte branched 1) 基因,金鱼草CYC( cycloidea) 基因和水稻 PCF1、PCF2 基因,这4 个基因编码的蛋白都包含一段由约 60 个氨基酸组成的保守序列,该保守序列能形成一种非典型的螺旋 -环 - 螺旋结构( non - canonical basic -Helix - Loop - Helix structure)。根据其代表成员 TB1、CYC 和 PCFs 的首字母缩写,把能够编码这段保守氨基酸序列的基因命名为 TCP 基因,把这段保守的氨基酸序列命名为 TCP 结构域。
TCP 结构域中的螺旋 - 环 - 螺旋区域( bHLH) 含有两个由保守的亲水氨基酸构成的中性螺旋结构和一个负责连接两个螺旋区域的环结构。在第二个螺旋区域有一个特异的“LXXLL”基序,动物bHLH 蛋白的研究结果表明这段基序可以通过调控转录的共激活单元结合到核定位蛋白上。
TCP 家族成员除了含有 TCP 结构域外,还存在一个保守的 R 结构域,R 结构域并不是所有的TCP 转录因子共有的,它富含精氨酸、赖氨酸和谷氨酸等极性氨基酸,可以形成一个 亲水性的 α 螺旋。
前人对 CYC、TB1、PCFs 和其他9 条预测的拟南芥和玉米 TCP 基因进行了进化分析,结果表明,这 12 个基因可以被分为两个亚家族,一个包含有 CYC 和 TB1,命名为 CYC/TB1 亚家族; 另一个包括 PCFs,命名为 PCF 亚家族。其中 TCP 结构域普遍存在于所有的 TCP 家族成员中,而 R 结构域则特异存在于 CYC/TB1 亚家族的一些基因中。对多种菊亚纲植物的 TCP 基因家族成员之间的进化关系进行聚类分析,表明 TCP 基因家族在各物种间的多样化对其形态学上的进化具有重要作用。Yao 等通过对拟南芥 23 个 TCP 基因及水稻 22 个 TCP 基因的进化分析,将 TCP 家族分为三个亚家族: Class Ⅰ、Class Ⅱ和 Class Ⅲ,同时对其结构域进行了序列比对,并分析了不同基因在不同组织中的表达模式。
TCP 家族转录因子主要在快速生长的组织或器官中表达,与植物细胞的分化和发育有密切关系。玉米 TB1 基因影响侧生分生组织的生长和发育; 水稻 TB1 基因被认为是水稻侧枝发育的负调控元件; 拟南芥 TCP16 基因对早期花粉的发育有重要作用。拟南芥 TCP20 基因能够结合 CYCB1 的 GCCCR 元件,具有调控细胞分裂和生长的作用,可与 AtTCP9 基因对抗性地调控拟南芥叶片发育和茉莉酸代谢过程。研究表明,拟南芥中的 5 个 TCP 基因: TCP2、TCP3、TCP4、TCP10 和 TCP24 是 microRNA 319a 的靶基因,参与调控叶片的形态发生。豆科模式植物百脉根中的 LjCYC1 和 LjCYC3 参与调控花分生组织的生长发育过程,另一类豆科植物豌豆中的CYC 类 TCP 基因也参与控制背腹轴向不同类型花瓣的发育。
2010 年 8 月,Velasco 等在 Nature Genetics 上发表了关于‘金冠’苹果基因组测序工作的文章,标志着苹果全基因组序列的测定已经完成,这一成果将苹果生物学研究带入了崭新的系统生物学时代,为研究者从全基因组水平对苹果进行研究奠定了基础。苹果 RING finger、MAPK 和 MAP-KK、MdWRKY 转录因子及 DREB 转录因子家族基因已通过生物信息学的方法鉴定出来,并进行了全基因组分析和基因功能预测。目前TCP 转录因子家族的研究主要集中在模式植物拟南芥、水稻和玉米中,蔬菜和果树尤其是苹果中的报道还非常少。本文从苹果全基因组出发,利用生物信息学的方法,鉴定出苹果全部的 TCP 转录因子,对其家族进化关系及结构域序列保守性进行了系统预测和分析,以期为进一步研究 MdTCP基因的作用奠定一定的理论基础。
1 材料与方法
从苹果功能基因组数据库下载 TCP 转录因子家族序列; 从GDR 数据库下载‘金冠’苹果全基因组序列,构建本地 Blast 数据库,以拟南芥 TCP 转录因子家族基因序列执行本地 Blast( 1e -003) 搜索; 合并两部分结果,利用 Perl 程序筛选,去掉重复序列,所得结果再利用 PFAM 及 NCBI - CDD 工具进行蛋白结构预测,删除不含 TCP 结构域的基因。利用 ExPASy Proteomics Server,对所有 TCP 基因编码蛋白进行分子量、等电点预测。
从 GDR 数据库下载‘金冠’苹果基因组信息文件( assembly gff3 file) ,利用 Perl 程序选取 TCP基因的染色体位置信息,并利用 MapDraw 工具进行染色体定位作图。
用 MUSCLE 进行序列比对,选取 TCP 结构域序列,再利用 MEGA5 构建进化树。进化树生成算法采用邻接法( Neighbor - Joining,NJ) ,校验参数Bootstrap 重复 1 000 次; 基因结构利用生物学软件 Gene Structure Display Server( GSDS) 分析获得; 保守性分析则采用DNAMAN 生物学软件进行保守序列比对。
2 结果与分析
2. 1 苹果 TCP 转录因子家族成员的鉴定
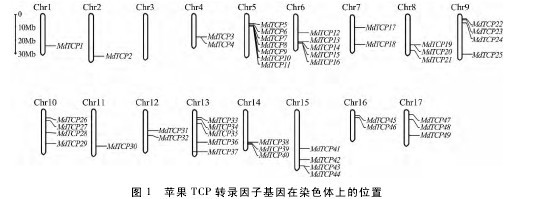
利用生物信息学方法,从苹果全基因组鉴定得到 52 个 TCP 转录因子家族成员,根据其系统进化树分析结果,对其进行了系统编号。MdTCP对应的基因编号、基因组登录号、编码序列长度、外显子数量、蛋白长度、分子量、等电点、所在染色体位置和拟南芥同源基因等特征。由表 1可知,MdTCP 蛋白长度在 115 aa( MdTCP50) ~612 aa ( MdTCP27 ) 范 围 内,等 电 点 在 5. 41( MdTCP32) ~ 10. 65 之间( MdTCP3) 。如图 1 所示,有 49 个 MdTCP 基因存在于不同染色体上,其中 5 号染色体最多,有 7 个; 3 号染色体上没有分布; 其余染色体上各有 1 ~5 个 MdTCP 基因分布。另外有 3 个 MdTCP 基因 MdTCP50、MdTCP51 和MdTCP52 未发现相应的染色体定位信息。【图1】
2.2 苹果 TCP 转录因子家族系统进化及基因结构分析
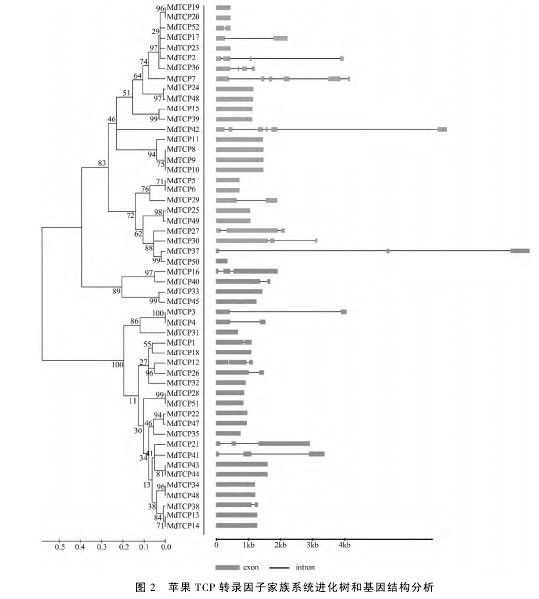
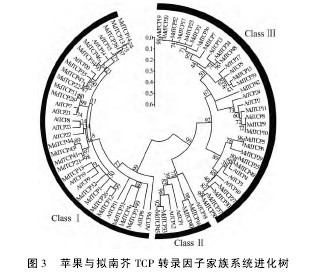
如图2所示,MdTCP 基因的内含子和外显子数量表现出了较高的保守性,其中没有内含子的 MdTCP 基因有32个,占总数的61.5%;有1个内含子的有9个,占17.3%; 有2 个内含子的有7 个,占13.5%; 有3 个和5 个内含子的MdTCP 基因分别有2个,各占3.8%。2. 3 苹果与拟南芥 TCP 转录因子家族系统进化及保守结构域分析进化树分析( 图 3) 表明,52 个 MdTCP 转录因子可分为 ClassⅠ、ClassⅡ和 Class Ⅲ三类,分别包括22、4 和 26 个,其中第二类数量较少,这与拟南芥和水稻中的规律是一致的。前人研究表明,拟南芥中有24 个 TCP 家族成员,由图3 可知,不同的AtTCP 转录因子与 52 个 MdTCP 转录因子有着不同的同源性,例如,AtTCP20 与 MdTCP22、MdTCP35、MdTCP47 同源性较高,AtTCP9 与 MdTCP18 同源性较高。对预测的52 个苹果 TCP 转录因子蛋白序列进行保守结构域序列比对,结果发现,三类苹果 TCP蛋白中均包含有 TCP 结构域,即 TCP 转录因子的特征序列———bHLH 结构域( 图4) 。【图2-4】
3 讨论与结论
随着越来越多物种全基因组测序工作的完成,海量的基因组数据、信息摆在我们面前,如何从这些数据中找到我们所需要的部分成为亟待解决的难题。利用生物信息学方法对基因组数据进行分析,研究各个物种间的进化关系和相互间的亲缘关系,以及物种内各基因家族成员的进化关系,能够为后续基因功能的研究提供一定的借鉴。
本文利用生物信息学方法,对苹果基因组中的TCP 转录因子进行了鉴定与分析。以期为今后研究苹果 TCP 转录因子的具体生理作用奠定一定的理论基础。
TCP 转录因子在植物中分布较广,各物种间的分布数量存在一定差异。拟南芥 TCP 转录因子家族有 24 个成员,水稻有 22 个成员,本研究鉴定了 52 个苹果 TCP 家族成员。通过系统进化分析发现,MdTCP22、MdTCP35、MdTCP47 与AtTCP20 的同源性较高,MdTCP18 与 AtTCP9 的同源性较高,结合拟南芥研究结果,这些基因可能在苹 果 生 长 发 育 及 代 谢 过 程 中 起 作 用; 而MdTCP8、MdTCP9、MdTCP10、MdTCP11、MdTCP25和 MdTCP49 与 AtTCP2、AtTCP3、AtTCP4 和 At-TCP10 的同源性较高,说明它们有可能作为 mi-croRNA 的靶基因参与苹果叶片的发育。借鉴拟南芥、水稻及其它物种中 TCP 转录因子在植物生长发育和相关代谢过程中的功能研究,探索MdTCP 转录因子在苹果生长发育尤其是叶片和侧枝、某些激素代谢过程中的功能及其相互作用机制是今后苹果相关研究的重点。
参 考 文 献:
[1] Doebley J,Stec A,Gustus C. Teosinte branched1 and the ori-gin of maize: Evidence for epistasis and the evolution of domi-nance[J]. Genetics,1995,141( 1) : 333 - 346.
[2] Luo D,Carpenter R,Vincent C,et al. Origin of floral asym-metry in Antirrhinum[J]. Nature,1996,383: 794 - 799.
[3] Cubas P,Lauter N,Doebley J,et al. The TCP domain: A mo-tif found in proteins regulating plant growth and development[J]. Plant Journal,1999,18( 2) : 215 -222.
[4] Heery D M,Kalkhoven E,Hoare S,et al. A signature motif intranscriptional co - activators mediates binding to nuclear re-ceptors[J]. Nature,1997,387: 733 - 736.
[5] Kosugi S,Ohashi Y. PCF1 and PCF2 specifically bind to ciselements in the rice proliferating cell nuclear antigen gene[J].The Plant Cell,1997,9( 9) : 1607 - 1619.

通过定量PCR, 发现FT基因会影响到根毛极性生长相关基因的表达.FT是一个转录因子, 可以直接结合特定基因调控表达, 而关于FT是否直接绑定根毛相关基因的启动子从而调控基因表达, 还需要进一步的研究....

干旱、低温和高盐等非生物逆境胁迫是农业生产中严重的自然灾害,严重影响了植物的生长发育及农作物的产量。...