植物学论文


0、 前言
种群是在一定空间和时间内的同种生物个体的总和,种群结构和动态变化是种群生态学的重要研究内容. 种群结构能够反映种群在时间上和空间上的变化规律,从而体现种群动态及其群落的演替趋势. 分析种群结构有利于认识种群的生物学特性,种内和种间关系、种群与环境因子之间的相互关系,从而掌握种群动态变化规律及其生态过程.种群结构作为种群特定时间和空间上的外在表现形式,在种群的发展过程中存在相互密切联系. 种群的年龄结构不仅在不同树种之间存在差异,而且同一树种在不同群落类型、不同生境和不同发育阶段其表现也有所不同. 优势种的生长发育、资源利用及竞争等过程对群落结构的形成和维持、甚至群落演替产生直接或间接的影响,因此,森林群落优势种的结构正受越来越广泛的关注.胡桃楸 Juglans mandshurica 属胡桃科Juglandaceae 胡桃属 Juglans 落叶高大乔木,为第三纪孑遗植物,被《中国植物红皮书》列为三级保护植物. 胡桃楸树型优美,材质良好,果实可食,具有很好的观赏和经济价值,主要分布于东北、华北及河南、山西等地,雾灵山自然保护区是胡桃楸分布较为集中的地区之一. 以往对雾灵山森林植被的研究主要是对油松、山杨等森林群落的研究,尚未有过对胡桃楸种群结构研究的报道. 本文通过对雾灵山自然保护区胡桃楸种群结构的分析,探讨其动态变化规律及其影响机制,以期对雾灵山植物种群的保护提供理论依据和科学研究的基础材料.
1、 研究区概况
雾灵山位于河北省兴隆县西北部,与北京市密云县、承德地区的滦平县、滦县、承德县相邻,面积143 km2,主峰海拔 2 118 m,为京东第一高峰,属暖温带半湿润大陆性季风气候区、雨热同季、冬长夏短、昼夜温差大. 雾灵山自然保护区是暖温带落叶阔叶林带向温带针阔混交林带的过渡地带,也是中国东部湿润森林向西部半干旱、干旱区森林、草原变化的地带,还是华北、东北、内蒙古三大植物区系交汇区域,该区域植被既有暖温带带阔叶林特征,又有温带针阔混交林特征. 地带性植被为以栎属 Quercus,桦木属 Betula,杨 Populus 等为主的暖温带落叶阔叶林和以油松 Pinus tabulaeformis 占优势的温带针叶林. 雾灵山山地植物垂直特征明显,海拔900 m 以下为低山农田果林灌丛带,900 m ~1 500 m 为中山下部落叶阔叶林带,1 500 m ~ 1 700 m 为中山上部针阔混交林带,1 700 m ~ 1 900 m 为中山上部寒温性针叶林带,1 900 m 以上为山顶次生草甸带. 土壤以山地褐土为主,主要分布在中山地带,土层厚度一般为 20 cm ~50 cm.
2、 研究方法
2. 1 样地调查
对雾灵山自然保护区胡桃楸群落采取传统的典型性样方法,对保护区胡桃楸群落分布较为集中的杵榆沟进行调查. 对样方内的胡桃楸进行每木必测,测定其树高、胸径、冠幅等. 此外,对样方内胡桃楸个体进行定位,方法为以每个样地东北角为原点,测定其坐标系中的位置(横坐标用 x 表示,纵坐标用 y 表示) . 在每个样方的原点为起点测量一个 5 m × 5 m的灌木层和1 m ×1 m 的草本层进行群落调查,观察乔木层以下的灌木层和草本层的生长情况. 同时,记录样方所在地的林地郁闭度、海拔、坡度、坡向、物候相和生活力等环境因子.
2. 2 种群结构
2. 2. 1 种群径级结构
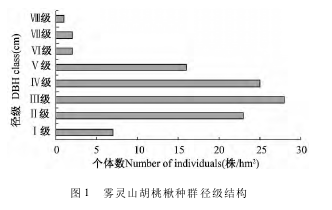
将样方内林木以胸径大小分级,以立木级结构代替年龄结构分析种群动态. 虽然龄级和径级有所不同,但在一定环境下同一树种的龄级和径级对环境的反应规律具有一致性. 种群的径级结构以胸径大小为标准,0 ~5 cm 为第 Ι 级,以后胸径每增加5cm 径级增加 1 级,由此将雾灵山自然保护区胡桃楸种群径级共划分为 8 个径级(Ⅰ ~ Ⅷ) . 把树木径级从小到大的顺序看作时间顺序,统计样地各径级胡桃楸的株数. 依据径级将雾灵山自然保护区胡桃楸种群划分为幼苗(Ⅰ级) 、幼树(Ⅱ级) 、中龄树(Ⅲ ~ Ⅳ级) 和成龄树(Ⅴ ~ Ⅷ级) 并绘制种群径级结构图.
2. 2. 2 种群静态生命表的编制
以树木的胸径作为度量树木年龄的指标,统计各个龄级内的胡桃楸个体数,编制雾灵山自然保护区胡桃楸种群静态生命表.
2. 2. 3 存活曲线、死亡曲线及消失曲线的绘制
根据静态生命表,以径级为横坐标,以标准化存活个体数 lx 为纵坐标,绘制存活曲线. 以 qx、kx 为纵坐标,以径级相对的龄级为横坐标绘制死亡率曲线和消失率曲线.
2. 2. 4 龄级分布与微地形的关系
统计不同海拔高度和坡度胡桃楸种群龄级结构的分布情况,绘制雾灵山自然保护区胡桃楸种群龄级分布与海拔及坡度等微地形关系的柱状图.
3、结果与分析
3. 1 种群径级结构
雾灵山自然保护区胡桃楸种群径级结构见图1. 从图 1 可以看出雾灵山自然保护区胡桃楸径级结构呈现典型的“金字塔”型,种群年龄属于稳定型. 各径级中Ⅱ级、Ⅲ级和Ⅳ级的胡桃楸所占比重大,除此外,胡桃楸个体数基本随径级增加而减少. 然而,种群径级结构在不同生境的样地中存在明显差异,样地 a 胡桃楸种群主要集中在第Ⅱ、Ⅳ径级,幼树和中龄树数量较多,而样地 b 的胡桃楸种群主要集中在第Ⅳ、第Ⅴ径级,成龄树数量较多. 而且,样地 a 的第Ⅶ级和样地 b 的第Ⅲ级及第Ⅷ级出现“断层”现象,这也某种意义上与种群“潮动”发展现象类似.
3. 2 种群静态生命表的绘制
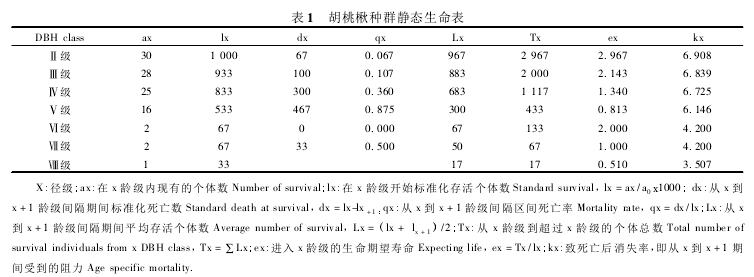
根据群落调查资料,将两个样地对应的径级个体数合并,编制了雾灵山自然保护区胡桃楸静态生命表. 由于第 Ι 径级存活数小于第Ⅱ径级的存活数,这与实际不符,为了满足生命表的编制要求,把第 Ι径级的幼苗存活数并入第Ⅱ径级幼树当中编制种群静态生命表. 由表 1 可以看出,雾灵山自然保护区胡桃楸种群死亡率随着径级的增加大致为上升趋势,这说明雾灵山自然保护区中胡桃楸种群的生长发育主要受其自身的生物学特性和种内竞争影响. 其中,第Ⅳ径级和第Ⅴ径级的死亡率较高,这是由于这两径级的胡桃楸个体数较多,个体间争夺资源、空间而发生激烈竞争,死亡率上升所导致. 第Ⅴ径级后的死亡率降低,进而接近平稳,随着龄级的增加,死亡率再逐渐上升,直到其生理年龄. 种群生命期望随着径级的增加大致呈递减趋势,这种趋势是符合种群生物学特征的. 而第Ⅵ径级的生命期望值高于第Ⅴ径级,这是与第Ⅲ径级开始的自疏作用和树种个体较少,生长空间变大而生命力增强作用有关,随后生命期望值又逐渐下降.
3. 3 存活曲线、死亡率曲线和消失率曲线的绘制
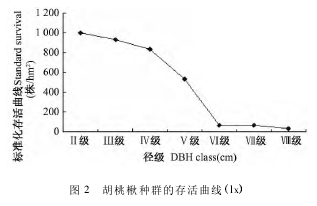
存活曲线是借助存活个体数来描述种群个体在各龄级的存活状况的曲线,可划分为 Deevey Ⅰ型、Deevey Ⅱ型和 Deevey Ⅲ 型. 根据种群静态生命表,以径级为横坐标,存活量的自然对数为纵坐标,绘制雾灵山自然保护区胡桃楸种群的存活曲线(图2) . 由图 2 可以看出,雾灵山自然保护区胡桃楸种群的存活曲线接近 DeeveyⅢ型,Ⅱ———Ⅳ径级的存活数目较多而后存活数目缓慢下降,从Ⅳ———Ⅵ径级的胡桃楸存活数目急剧下降,这是由于群落内部不同种群为争夺生存资源相互竞争,死亡率上升影响导致的.
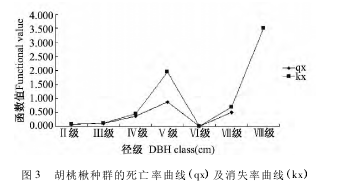
如图 3 所示,胡桃楸的死亡率曲线(qx) 和消失率曲线(Kx) 变化的趋势基本一致,死亡率和消失率都出现两个高峰期. 第一个高峰期出现在第Ⅴ径级,死亡率高达 87. 5%; 第二个高峰期出现在第Ⅶ径级和第Ⅷ径级,死亡率达 50%.
3. 4 龄级分布与微地形的关系
以各样方点所在坡度及海拔高度为横坐标,各径级胡桃楸个体数为纵坐标,分别绘制雾灵山自然保护区胡桃楸种群龄级分布与坡度关系的柱状图(图 4) ,以及与海拔高度关系的柱状图(图5) . 从图 4 可以看出,在坡度 30° ~ 40°范围中各龄级分布较广泛,其中Ⅱ龄级和Ⅲ龄级所占数量多.坡度 10° ~ 20°范围中Ⅰ龄级分布较多,Ⅱ龄级和Ⅲ龄级分布少,Ⅶ ~ Ⅷ龄级几乎没有分布,这是由于低坡度范围内适宜种植作物,便于灌溉,受农田利用影响,中龄树和成龄树种植的数量受到限制.而坡度越陡,区域的生境中幼苗和幼树不易生长,龄级分布也较为稀少,主要分布中龄树和数量较少的成龄树.

如图 5 所示,海拔高度在 400 ~500 m 范围中胡桃楸种群龄级分布稀疏,主要是因为在这片海拔高度中土地利用类型为农田用地,胡桃楸群落分布少.在海拔高度为 800 ~ 1 000 m 范围中胡桃楸种群各龄级分布广泛,其中,Ⅱ ~ Ⅳ的低龄级胡桃楸分布数量较多,对应的幼树和中龄树所占比重大,这片高程范围内属于雾灵山自然保护区中山松栎林亚带,属于森林林冠层群落植被集中分布区.
4、 结 论
4. 1 种群径级结构与稳定性
雾灵山自然保护区胡桃楸种群径级结构呈“金字塔”型,幼树和中龄树所占比重大,幼苗的补给少,在不同的样地中种群径级结构和个体数量存在较明显的差异.
4. 2 群落结构与胡桃楸种群的关系
林下灌木层以土庄绣线菊、小花溲疏、太平花、小叶椴和华北绣线菊等为主. 林下灌木层发育较好,与幼苗争夺光、空间和养分等,使得幼苗的生长受限制,成活率下降. 而幼苗进入幼树阶段,其竞争能力和生长能力增强,成活率明显提高. 由于幼苗的补给不足,林冠层不是由胡桃楸构成的纯树种.林冠层中的油松、棘皮桦和山杨也影响胡桃楸的生长. 同时,林冠层的种群结构直接影响林下的灌木层和草本层种类的组成和结构. 幼树进入林冠层集中在有限的空间里,为争夺空间和光照,与群落同冠层树种竞争,有部分个体死亡. 自疏作用不仅减低了种内竞争,同时为林下灌木层、草本层和立木更新层物种生长提供了空间和光照.
参考文献:
[1] 江洪. 云杉种群生态学[J]. 北京: 中国农业出版社,1992: 8 -26.
[2] 杨慧,娄安如,高益军,等. 北京东灵山地区白桦种群生活史特征与空间分布格局[J]. 植物生态学报,2007,31(2) : 272 -282.
[3] 王本洋,余世孝. 种群分布格局的多尺度分析[J]. 植物生态学报,2005,29(2) : 235 -241.
[4] 牛丽丽,余新晓,岳永杰. 北京松山自然保护区天然油松林不同龄级立木的空间点格局[J]. 应用生态学报,2008,19(7) : 1414 - 1418.
[5] 贺士元,邢其华,伊祖棠. 北京植物志[M]. 1993.
[6] 李宏,刘建中,郭逍宇,等. 雾灵山土壤———植物地理实习指导(上) [M]. 北京: 首都师范大学出版社,2012,6: 81- 104.
[7] 张桥英,罗鹏,张运春,等. 白马雪山阴坡林线长苞冷杉种群结构特征[J]. 生态学报,2008,1(1) .
[8] 阳含熙,伍业钢. 长白山自然保护区阔叶红松林林木种属组成、年龄结构和更新策略的研究[J]. 林业科学,1988,24(1) : 18 - 27.

1引言(Introduction)报春苣苔(Primulinatabacum)为中国特有,属苦苣苔科的多年生草本植物;因生境特殊,昆虫难以进行异地授粉,繁殖率极低,野外濒临绝灭,已被列为第一批国家Ⅰ级重点保护野生植物名录。国内外文献对报春苣苔的研究现涉及无性繁殖、体...
果实(种子)异型性(Heterocarpy)是指同一植株的不同部位产生不同形态或行为果实(种子)的现象。由于果实异型性可以导致植物在种子扩散、休眠与萌发等方面产生不同生态生物学特性,因此在降低密集制约、避免同胞竞争上效应明显,是植物在时空...