小肠缺血再灌注损伤( small intestinal ischemia-reperfusion injury,IIRI) 是一种临床常见的病理现象,常危 及 生 命。肠 系 膜 上 动 脉 ( superior mesentericartery,SMA) 阻塞或失血性休克等许多损伤因素都能导致小肠缺血再灌注损伤的发生。小肠缺血再灌注可导致严重的局部创伤并引起多器官功能障碍,常可累及到肺组织,而且有研究显示继发的急性肺损伤( acute lung injury,ALI) 或/和急性呼吸窘迫综合征( acute respiratory distress syndrome,ARDS) 是造成小肠缺血再灌注损伤病人死亡的主要原因。因此,为找到新的防治措施,降低小肠缺血再灌注损伤患者的死亡率,进行小肠缺血再灌注肺损伤分子机制的研究是非常关键和必要的。
具有 CCCH 锌指结构域的蛋白 12D( zinc fingerCCCH type containing 12d,Zc3h12d) ,也称为 TFL 或P34,是近期发现的包含单个锌指结构域蛋白家族中的一员,拥有相对分子质量 ( Mr) 58 000 和36 000 两 种 剪 接 体, 在 炎 症 因 子 或 脂 多 糖( lipopolysaccharide,LPS) 刺激单核细胞时其表达量会发生变化,从而对细胞的生理和病理过程产生影响。然而 Zc3h12d 在动物肺损伤中的表达尚未见报道。本实验通过建立大鼠 IIR 肺损伤模型,初步观察 Zc3h12d 在急性肺损伤大鼠肺中的表达情况并探讨其可能的作用及意义,为进一步了解 IIR 肺损伤发生、发展的分子机制奠定基础。
1 材料和方法
1. 1 材料 纯化 LPS、兔抗大鼠 β-actin 抗体、辣根过氧化酶标记的兔抗山羊 IgG、辣根过氧化物酶标记的山羊抗兔 IgG 购自 Sigma 公司; 山羊抗大鼠 Zc3h12d 抗体购自 Santa Cruz 公司; 大鼠 TNF-α、IL-1β ELISA 试剂盒购自北京鼎国生物技术公司; 两步法免疫组化试剂盒购自武汉博士德生物工程有限公司; TRIzol 试剂盒、逆转录试剂盒及实时定量 PCR( realtime quantitative PCR,qRT-PCR) 试剂盒购自大连 TaKaRa 公司; SDS 细胞裂解液、ECL 显色液购自西安晶彩生物技术公司; BCA 蛋白定量试剂盒购自上海碧云天生物技术有限公司。电热恒温干燥箱为北京科伟永兴仪器有限公司产品,荧光显微镜为徕卡公司产品,紫外分光光度计为 Beckman 公司产品,实时定量 PCR 仪、Western blot 蛋白电泳仪为 Bio-Rad公司产品。
1. 2 方法
1. 2. 1 实验动物分组与模型制备 健康雄性 SPF 级 SD 大鼠40 只,体质量 200 ~ 230 g,由第四军医大学动物实验中心提供。大鼠随机分为对照组( contro1) 、缺血再灌注 30 min 组( IR30 min) 、缺血再灌注 60 min 组( IR60 min) 和缺血再灌注120 min 组( IR120 min) ,每组 10 只,均适应性饲养 7 d 后用于实验。实验前禁食 12 h,自由饮水。10 g/L 戊巴比妥纳40 mg / kg腹腔注射麻醉,经大鼠口气管插管并连接呼吸机。
麻醉后常规备皮消毒,行正中开腹手术,切口 3 ~4 cm 入腹,探查大鼠腹腔内情况; 探查后将全部消化道及网膜向右掀起,找到腹主动脉分支肠系膜上动脉( superior mesenteric artery,SMA) 并钝性分离; 对照组大鼠缝合切口,3 个缺血再灌注组以无创伤动脉夹夹闭 SMA 起始部,缺血 60 min 后恢复再灌注,分别再灌注 30 min、60 min 和 120 min; 过量戊巴比妥纳腹腔注射,麻醉处死。
1. 2. 2 肺组织湿质量 / 干质量( W / D) 比值测定 取大鼠右肺上叶称取质量为湿重,而后放入70℃恒温干燥箱中烘烤12 h 称质量为干质量,以湿质量/干质量得出肺湿干质量( W/D) 比值。
1. 2. 3 ELISA 检测血清及肺组织上清液中 IL-1β 及 TNF-α 水平 心脏取血制备血清,取左肺叶一部分制备组织匀浆液,采用 ELISA 检测血清及肺组织上清 IL-1β 及 TNF-α 的水平。
心脏取血约3 mL,37℃水浴加热 30 min 后,4℃ 2 000 g 离心20 min,收集上清即血清,取肺组织约 300 mg,用 0. 01 mmol / LPBS( pH7. 4) 1. 5 mL 充分匀浆后,4℃ 12 000 g 离心 20 min,收集上清,两者均采用双抗体夹心 ELISA 进行检测。
1. 2. 4 肺组织病理学检查 取右肺中下叶置于 100 mL / L 的中性甲醛中固定 24 h、梯度酒精脱水、石蜡包埋,保存于4℃ 。使用时切片( 厚度 3 μm) 、HE 染色、中性树胶封片,光镜下观察肺组织病理形态学改变。
1. 2. 5 肺组织 Zc3h12d 蛋白免疫组化染色 肺组织石蜡切片常规脱蜡至水后,尿素消化; 柠檬酸缓冲液中微波加热修复抗原,50 g/L 过氧化氢封闭内源性过氧化物酶。按两步法免疫组化染色: 滴加 BSA 封闭 30 min,封闭后甩去 BSA,加山羊抗大鼠 Zc3h12d 抗体,4℃冰箱孵育过夜; 37℃水浴复温45 min,滴加辣根过氧化物酶标记的兔抗山羊二抗室温孵育45 min; DAB 显色、适时终止; 苏木素复染胞核,中性树胶封片。光学显微镜下观察拍照。阴性对照以 PBS 代替一抗。
1. 2. 6 qRT-PCR 检测 使用 TRIzol 试剂盒提取肺组织总RNA,紫外分光光度计测定 RNA 含量、纯度。根据含量将提取的 RNA 调制成相同浓度,进行逆转录,Zc3h12d 上游引物为 5'-GGTCAGCACAATCCACCATCA-3',下游引物为 5'-TG-TATCCCACCTACCCAAGCA-3'; 内参照为 β-actin,上游引物为5'-GGAGATTACTGCCCTGGCTCCTA-3',下游引物为 5'-GACT-CATCGTACTCCTGCTTGCTG-3'。 反 应 条 件 为 37℃ 孵 育40 min,70℃ 10 min。使用 SYBRGreenⅠReal Time PCR 法扩增 cDNA,扩增条件: 95℃ 10 min,94℃ 30 s,58℃ 30 s,72℃ 30 s,重复 30 个循环。以 β-actin 为内参照,使用 2- △△Ct相对定量计算公式计算 Zc3h12d 的 mRNA 相对表达量。
1. 2. 7 Western blot 检测肺组织 Zc3h12d 蛋白的表达 取肺组织100 ~200 mg,加入适量 SDS 细胞裂解液、PMSF,于冰上充分匀浆; 4℃、12 000 r/min 离心20 min,取上清,BCA 法蛋白定量; 加上样缓冲液煮沸 5 min,保存于 -70℃。每孔上样蛋白浓度为 40 μg 进行 SDS-PAGE,电泳后转移到 NC 膜上。
滴加 50 g/L 脱脂牛奶 50 mL 封闭 1 h,使用山羊抗大鼠Zc3h12d 抗体及辣根过氧化物酶标记的兔抗山羊 IgG 孵育目的蛋白,以观察 Zc3h12d 蛋白两种剪接体的表达情况,用兔抗大鼠 β-actin 抗体及山羊抗兔 IgG 孵育内参照 β-actin 蛋白。ECL 显色,凝胶成像系统观察并照相。
1. 2. 8 统计学分析 实验数据录入 SPSS16. 0 统计软件进行统计分析。各组数据以 x ± s 表示,组间比较采用单因素方差分析( One-way ANOVA) 。检验水准 α = 0. 05,P < 0. 05 表示差异有统计学意义。
2 结果
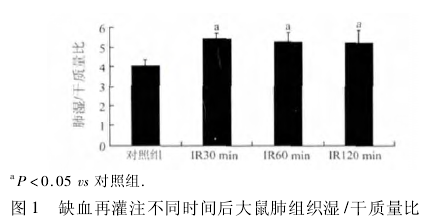
2. 1 肺组织湿 /干质量比值 小肠上动脉缺血再灌注肺损伤( IIRI) 模型组与对照组相比,IR30 min 组比值开始增大( F =6. 56,P <0. 05) ,表明肺微血管通透性开始增加,肺水肿形成,IR60 min 组比值大于对照组( F =3. 19,P <0. 05) ,IR120 min 组比值增幅较IR60 min 减小但仍大于对照组( F = 1. 66,P < 0. 05) ,表明在IR60 min 组和IR120 min 组大鼠仍有肺水肿( 图1) 。
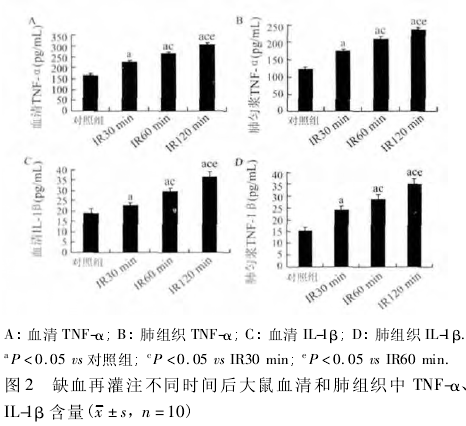
2. 2 血清和肺组织匀浆中 TNF-α 和 IL-1β 的变化ELISA 检测结果显示 IR 致伤后,IR30 min 组大鼠血清和肺匀浆中 TNF-α 浓度较对照组升高( F =43. 91,P < 0. 05; F = 41. 44,P < 0. 05) ,同时肺匀浆中 IL-1β浓度也较对照组升高( F = 9. 80,P < 0. 05) 。IR60min 组较 IR30 min 组血清和肺匀浆中 TNFα 浓度升高( F =28. 81,P <0. 05; F =20. 30,P <0. 05) ,血清中 IL-1β 浓度也升高( F = 8. 89,P < 0. 05) 。IR120min 组较 IR60 min 组血清和肺匀浆中 TNF-α 浓度升高( F =28.95,P <0.05; F =10.23,P <0.05) ,同时血清中 IL-β 浓度也升高( F =4.60,P <0.05,图2) 。
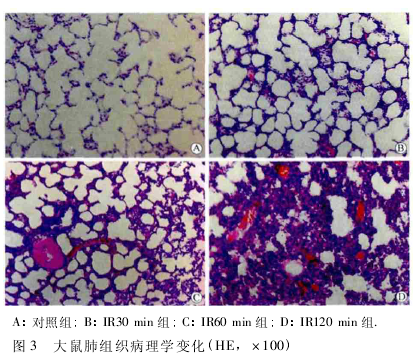
2. 3 肺组织病理学变化 对照组肺小叶结构完整,肺泡间隔无增厚,无渗出,血管周围未见炎症细胞浸润。IR 致 ALI 模型组可见肺泡毛细血管破坏,肺泡壁明显增厚,肺泡腔内可见大量粉红色渗出液和红细胞,间质大量炎症细胞浸润,有明显的炎症改变( 图3) 。
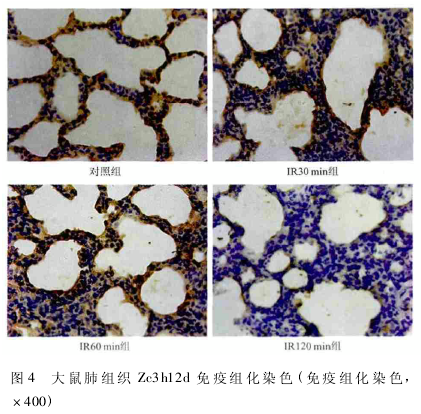
2. 4 肺组织 Zc3h12d 的表达 肺组织 Zc3h12d 主要在肺泡细胞和肺间质细胞中表达,分布于细胞质中,呈棕黄色,而细胞核中未见明显表达,IR30 min 和IR60 min 组 与 对 照 组 相 比 染 色 无 明 显 变 化,IR120 min组染色与对照组相比明显变浅,呈淡黄色,为弱阳性( 图 4) 。
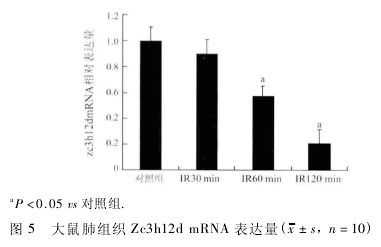
2. 5 qRT-PCR 检测样本肺组织 Zc3h12d mRNA 的表达 紫外分光光度计检测大鼠肺组织 A260 / A280比值,均在 1. 8 ~2. 0 之间,显示总 RNA 完整性好,纯度较高。肺组织中的 Zc3h12d mRNA 荧光定量扩增曲线呈 S 型,溶解曲线为单峰,无非特异性扩增及引物二聚体形成,结果可靠,故使用 qRT-PCR 测定Zc3h12d 的 mRNA 相对含量,发现 IR30 min 肺损伤模 型 组 中 mRNA 表 达 较 对 照 组 无 明 显 变 化( F =0. 22,P > 0. 05) ; IR60 min 组、IR120 min 组mRNA 与对照组相比表达明显减少 ( F = 5. 65,P < 0. 05; F = 15. 75,P < 0. 05 ) ,但 IR30 min 组 与IR60 min 组、IR60 min 组与 IR120 min 组 mRNA 表达比较无明显差异( F = 3. 14,P > 0. 05,P = 0. 0931;F = 4. 40,P > 0. 05,P = 0. 0504; 图 5) 。
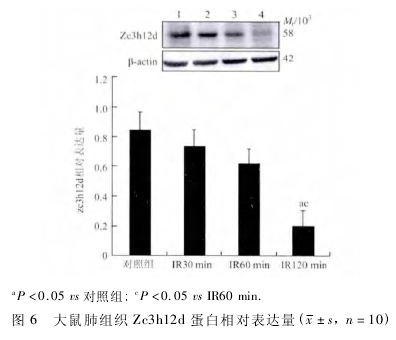
2. 6 Western blot 法检测肺组织 Zc3h12d 蛋白的表达 Western blot 法检测以 Zc3h12d/β-actin 吸光度值为结果,计算 Zc3h12d 蛋白的相对表达量,检测结果显示各组肺组织都有 Zc3h12d 蛋白的表达,但仅有相对分子质量( Mr) 58 000 剪接体而无 Mr36 000 剪接体表达; IR30 min 组、IR60 min 组和对照组相比无明显减少( F =0. 22,P >0. 05,P > 0. 05; F = 5. 65,P > 0. 05,P > 0. 05) ; IR120min 组与对照组相比表达明显减少( F =9. 11,P <0. 05) ; IR60 min 组与 IR30min 组比较无明显变化( F = 0. 35,P > 0. 05) 而 IR120min 组比 IR60 min 组表达量明显减少( F = 4. 85,P <0. 05,图 6) 。
3 讨论
临床上小肠缺血再灌注损伤( IIRI) 可由败血症、缺血、小肠移植、严重烧伤等多种病理生理改变引起,并导致一系列严重的组织器官损伤,其中主要包括 ALI 或/和 ARDS。目前 IIRI 的机制和治疗受到了广泛的研究,虽然其确切的机制还没有得到阐明,但是活性氧和炎症信号通路被认为在其中扮演了重要的角色。研究显示 NF-κB 通路在炎症反应和固有免疫中处于中心地位,并受到多种转录后调节,包括磷酸化和泛素化,而去泛素化酶( deubiquitinating enzymes,DUB) 被确定可以下调 NF-κB 的信号转导,从而达到抑制炎症反应的作用。Zc3h12d 是一种新发现锌指蛋白,因蛋白结构中有一个锌指结构域而得名。这一蛋白家族共有Zc3h12a、Zc3h12b、Zc3h12c、Zc3h12d 四种蛋白,其氨基酸序列同源性很高,但组织分布差异性很大。
Zc3h12d 主要分布于淋巴细胞、淋巴结、脾脏和肺组织中,且其 Mr58 000 及 36 000 剪接体的组合分布不同。这个蛋白家族研究较多的是 Zc3h12a,据文献报导 Zc3h12a 是一种 DUB,具有转录因子作用,炎症抑制作用和 miRNA 及 mRNA 的降解作用,其炎症抑制作用主要是通过去泛素化 NF-κB 等炎症通路来实现的,研究发现和 Zc3h12a 分子结构很相似的 Zc3h12d 不仅有调节细胞周期的作用,而且也和抑制肺癌有关,据猜测 Zc3h12d 是通过抑制 NF-κB信号通路起到抑制肺癌的作用,进一步的细胞实验证实 Zc3h12d 具有去泛素化作用,当巨噬细胞受到 LPS、鞭毛蛋白等多种外来抗原的刺激时,激活核转录因子 NF-κB,NF-κB 的激活会诱导 Zc3h12d 基因表达上调,而后产生大量的 Zc3h12d 蛋白再抑制NF-κB 信号通路,形成一个负反馈通路。这些研究表明 Zc3h12 蛋白可以通过调节 NF-κB 信号通路来抑制炎症反应,但是其具体的作用靶点及自身的调节机制仍然不清,动物实验也尚未见报道。
本实验通过夹闭 SD 大鼠肠系膜上动脉并进行再灌注,模拟临床小肠缺血再灌注肺损伤建立大鼠急性肺损伤模型。通过检测肺湿/干质量比( W/D)发现小肠缺血再灌注( IIR) 后大鼠形成持续性的肺水肿,ELISA 检测发现血清及肺组织中的炎症因子TNF-α、IL-1β 在缺血再灌后呈现不断升高的趋势,表明肺内炎症反应激活,炎症介质过度表达,引起了不可控的炎症反应,结合大鼠肺组织典型的肺损伤病理改变,表明大鼠小肠缺血再灌注肺损伤模型建立成功。通过免疫组织化学检查发现 Zc3h12d 蛋白存在于正常大鼠肺组织中,并分布于细胞质中,在再灌注损伤后,Zc3h12d 蛋白的表达呈下降的趋势但未见明显转位。qRT-PCR 及 Western blot 检查显示,急性肺损伤 120 min 后 Zc3h12d 在 mRNA 和蛋白水平的表达都明显降低,表明大鼠小肠缺血再灌注肺损伤后,炎症反应的激活下调了 Zc3h12d 的表达,这些结果结合先前的研究提示 Zc3h12d 蛋白在肺损伤中扮演保护性的角色,且受到了一定的调节。
目前研究认为 Zc3h12d 蛋白的两种剪接体无生物功能差异,而本实验通过 Western blot 检查发现在大鼠肺组织中仅有 Mr58 000 剪接体的表达,这些结果对研究 Zc3h12d 两种剪接体的功能提供了新的思路,为进一步研究小肠缺血再灌注肺损伤的分子调节机制奠定了基础。
参考文献:
[1]Harward TR,Brooks DL,Flynn TC,et al. Multiple organ dysfunctionafter mesenteric artery revascularization[J]. J Vasc Surg,1993,18( 3) : 459 -467.
[2]Mallick IH,Yang W,Winslet MC,et al. Ischemia-reperfusion injuryof the intestine and protective strategies against injury[J]. Dig DisSci,2004,49( 9) : 1359 - 1377.
[3] Brun-buisson C,Minelli C,Bertolini G,et al. Epidemiology andoutcome of acute lung injury in European intensive care units resultsfrom the ALIVE study[J]. Intensive Care Med,2004,30( 1) : 51- 61.
[4] Collard CD,Gelman S. Pathophysiology,clinical manifestations,and prevention of ischemia-reperfusion injury[J]. Anesthesiology,2001,94( 6) : 1133 - 1138.
[5] Oldenburg WA,Lau LL,Rodenberg TJ,et al. Acute mesentericischemia: a clinical review[J]. Arch Intern Med,2004,164( 10) :1054 - 1062.
[6] Huang SP,Qi DF,Liang J,et al. Putative tumor suppressorZc3h12d modulates Toll-like receptor signaling in macrophages[J].Cell Signal,2012,24( 2) : 569 - 576.
[7]Minagawa K,Yamamoto K,Nishikawa S,et al. Inhibition of G1 to Sphase progression by a novel zinc finger protein P58 TFL at P-bodies[J]. Mol Cancer Res,2009,7( 6) : 880 -889.
[8]Liang J,Wang J,Azfer A,et al. A novel CCCH-zinc finger proteinfamily regulates proinflammatory activation of macrophages[J]. JBiol Chem,2008,283( 10) : 6337 - 6346.
[9]杨 波,张志培,姜 鹏,等. N-myc 下游调节基因-2 在大鼠肠缺血再灌注肺损伤中的表达及其与 NF-κB p65 的相关研究[J].创伤外科杂志,2012,14( 6) : 543 -546.
[10] Khler HF,DeLucca IM,Sbragia Neto L. Enteral antioxidants inischemia / reperfusion injuries in rats[J]. Rev Col Bras Cir,2011,38( 6) : 422 - 428.
[11]Berlanga J,Prats P,Remirez D,et al. Prophylactic use of epidermalgrowth factor reduces ischemia / reperfusion intestinal damage[J].Am J Pathol,2002,161( 2) : 373 - 379.
[12]Chen ZJ. Ubiquitination in signaling to and activation of IKK[J].Immunol Rev,2012,246( 1) : 95 - 106.
[13]Tokunaga F,Iwai K. Linear ubiquitination: A novel NF-κB regulatorymechanism for inflammatory and immune responses by the LUBACubiquitin ligase complex[J]. Endocr J,2012,59 ( 8) : 641 - 652.
[14]Harhaj EW,Dixit VM. Deubiquitinases in the regulation of NF-κBsignaling[J]. Cell Res,2011,21( 1) : 22 - 39.
[15]Liang J,Song W,Tromp G,et al. Genome-wide survey and expressionprofiling of CCCH-zinc finger family reveals a functional module inmacrophage activation[J]. PLoS One,2008,3( 8) : e2880.
[16]Jura J,Skalniak L,Koj A,et al. Monocyte chemotactic protein-1-induced protein-1 ( MCPIP1) is a novel multifunctional modulator ofinflammatory reactions[J]. Biochim Biophys Acta,2012,1823( 10) : 1905 -1913.
[17]Xu J,Peng W,Sun Y,et al. Structural study of MCPIP1 N-terminalconserved domain reveals a PIN-like RNase nucleic acids research[J]. Nucleic Acids Res,2012,40( 14) : 6957 -6965.
[18]Wang M,Vikis HG,Wang Y,et al. Identification of a novel tumorsuppressor gene p34 on human chromosome 6q25. 1[J]. CancerRes,2007,67( 1) : 93 - 99.
[19] Minagawa K,Yamamoto K,Nishikawa S,et al. Deregulation of apossible tumour suppressor gene,ZC3H12D,by translocation of IGK@ in transformed follicular lymphoma with t( 2; 6) ( p12; q25) [J].Br J Haematol,2007,139( 1) :161 - 163.