����ѧ����


��������ά���Ǻ���ϵͳ������չ�����ڵĹ�ͬ�����ı�,��ɷι��ܲ����������,����Σ�����ཡ����Ŀǰ�о���Ϊ,����ά���Ļ����������̰������������Է��������ڴ�������ϸ������,�������ʽ�ԭ�Ľ����Ի���������ȡ�������ķ���֯�ṹ,����Ӱ��ε�ͨ���ͻ������ܡ� ��������ϸ����,��������Ƥϸ���ͳɼ�ϸ������,����άϸ������ֳ��������ڻ��ʽ�ԭ����ά���γɵ���Ҫ���ơ� TGF-β1 ����Ϊ���յ��γ���άϸ���������Ҫ��ϸ������,�����õķ����� β-catenin ;���߶���ء� ���о�ͨ���۲� β-catenin �����ó���TGF-β1 �յ��ķγ���άϸ������ֳ�����Ե�Ӱ��,̽�� β-catenin �����ó�������ά�����õ�ϸ������,Ϊ����ά���������ṩ�µ�˼·��
����1�� �����뷽��
����1.1 ��Ҫ�Լ�����Ʒ
��������γ���άϸ�� (CCC-REPF-1),pcDNA3.0����(Invitrogen),F-TrCP-Ecad ����(֣��ƽ���ڻ��� ),Lipofectamine2000 (Invitrogen),TGF-β1 (PeproTech),�� E-cadherin,�� α -SMA,�� Fn( SantaCruz),FITC ��־����С�� IgG(�人��ʿ�����﹤������˾),̥ţѪ��(�й������ļ���),���� DMEM ������(GIBCO),TRIzol �� RNA ��ȡ�Լ�(�人��ʿ�����﹤������˾),��ת¼�Լ���(Promega),ʵʱӫ�ⶨ���Լ���(�Ϻ��������﹤�̼�����������˾),α-SMA ���col�����col�����GAPDH ����(�Ϻ��������﹤�̼�����������˾)��
����1.2 �γ���άϸ������������������
���������������Ĵ���γ���άϸ��(CCC-REPF-1),�ú� 10%̥ţѪ�塢100 U/mL ��ù�ء�100 μg/mL ��ù�صĸ��� DMEM ������������ 5%�� CO2,37���˹���������,ϸ������������,���д���������ϸ������,��Ϊ����ϸ���顢TGF-β1 �̼��顢pcDNA3 תȾ�顢F-TrCP-Ecad תȾ�顣
����1.3 pcDNA3 �� F-TrCP-Ecad תȾ CCC-REPF-1ϸ��
������ȡ�������������� DNA, �� DNA Ũ�ȱ��á�
����ȡ������ϸ������,����ϸ����Ϊ 2×105cells/L,����12 �װ���,�۲�ϸ�������� 80%~90%ʱ,����Һ,����Ѫ����˫�������� 800 μL,��תȾ��100 μL ��˫����̥ţѪ����������м��� 4 ng ����,��Ϻ���5 min, �� 6 �� EP ��,3 ��Ϊ���� pcDNA3,3 ��Ϊ���� F-TrCP-Ecad,100 μL ��˫����̥ţѪ����������м��� 4 μL Lipofectamine2000 ��Ϻ��� 5 min,6 ��EP �� ,�������������Ϻ��� 20 min,�� 12 �� �� 6��������,����˫����̥ţѪ�����������ϴ 2 ��,����˫����̥ţѪ��������� 800 μL,������������ֱ���� 6 ����,ǰ����� 4~6 h �������������� תȾ 6 h ������������,����ֱ���� TGF-β1(5 ng/mL),���� 48 h��
����1.4 CCK-8 �����γ���άϸ����ֳ
����������ϸ������ϸ��Ũ���� 5000 ��/100 μL,��ÿ�� 100 μL ������ 96 �װ�,ÿ�鸴�� 8 ��,ÿ��ÿ���� CCK-8 10 μL,5%CO2�������ڼ������� 1h,�ֹ��ȼ��²ⶨ 450 nm ������ OD ֵ��
����1.5 Western ӡ�����ϸ���� E-cadherin��α-SMA��Fn�ı���
����ȡϸ���� PBS ϴ�Ӻ�ת�Ƶ� EP ��,���ĺ�������,���ռ�����ϸ����ϸ���ѽ�Һ�ѽ� 1 h ��,����30 min,ȡ����,BCA �����ʶ������ⶨ����Ũ��,ȡ�������� 30 μL �� SDS-PVDF ������Ӿ, תĤ��PVDF Ĥ,���� 4 �� 2~5 h,5%��֬ţ�̷�� 2 h�� ��4 ����С���� E ��𤵰�ס�α-SMA�� ��ά���ӵ�����¡����(ϡ��Ϊ 1��200)��Ĥ�Ӵ�������ҹ,��ϴĤ����Һ(TBST)����ɫҡ����ϴĤ 5 min×5 ��,����С�� IgG, �����·��� 2 h ��,TBST ϴĤ, ����ECL �� �����·�Ӧ 5 min �� ���� ,Taiphone ɨ ����,���� Gene Snap/Gene Tool ���������� GAPDH ����Ϊ�ڲζ������� E ��𤵰�ס�α-SMA����ά���ӵ���������ԻҶ�ֵ��
����1.6 �� RNA ���ἰʵʱӫ�ⶨ����ת¼-�ۺ�ø����Ӧ(Real-timePCR)
��������ϸ��ÿ���� 500 μL TRIzol Reagent,���Լ���˵����ȡ�� RNA ���� RT-PCR ��� α-SMA��col��col �����,GAPDH Ϊ�ڲΡ�
����GAPDH �� ������ : �������� 5 �@ - GGTGCT-GAGTATGTCGTGGAGT -3 �@ �� �� �� �� 5 �@ -CAGTCTTCTGAGTGGCAGTGAT -3,α -SMA �� ������ 5�@- AGCCAGTCGCCATCAGGAAC -3�@��������5�@- GGGAGCATCATCCCAGCAA-3�@,col��: �������� 5�@- GACATGTTCAGCTTTGTGTACCTC -3�@�������� 5�@- GGGACCCTTAGGCCATTGTGA -3�@col��:�������� 5�@- TTTGGCACAGCAGTCCAATGTA -3�@�������� 5�@-GACAGATCCCGAGTTCGCAGA -3�@��PCR �� Ӧ��ϵ���� : 95�� 20 s;95�� 3 s,60�� 30 s��
����40 ��ѭ���� ������������������ Ct ֵ�Ϳ�������
����1.7 ͳ��ѧ����
�����������ϲ��þ���±�����ʾ, ���ݴ�������SPSS 19.0 �������� t ��������,P <0.05 Ϊ������ͳ��ѧ���塣
����2�� ���
����2.1 �γ���άϸ���⾵�¹۲�
��������άϸ��������,�нϳ���ͻ��,���˳���Բ��,λ��ϸ������,�ɷ���״��դ��״���С� TGF-β1�̼���,ϸ�������������,��ͻ��ʧ,ϸ��������������,ϸ����϶��С��F-TrCP-Ecad תȾ���ϸ��������,�нϳ���ͻ��,ϸ���������Լ�϶��
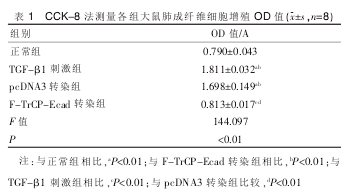
����2.2 CCK-8 ���۲����γ���άϸ����ֳ��������Ƚ�, TGF-β1 �̼��顢pcDNA3 תȾ���� OD ֵ��������,������Ƚ�,��ͳ��ѧ����;F-TrCP-Ecad תȾ�� OD ֵ�� TGF-β1 �̼������Լ���(P<0.01),��������Ƚ���ͳ��ѧ���졣 ���� 1��
��
����2.3 Western ӡ����������ϸ���� α-SMA��Fn��E-cadherin �ı���
������������Ƚ�, TGF-β1 �̼��顢pcDNA3 תȾ���� α -SMA��Fn ���ױ�����������, E-cadherin ������������ (P<0.01), ������Ƚ���ͳ��ѧ����;F-TrCP-Ecad תȾ��� TGF-β1 �� ���� α -SMA��Fn ���ױ�����������,E-cadherin ������������(P<0.01),��������Ƚ���ͳ��ѧ���졣 ���� 2��
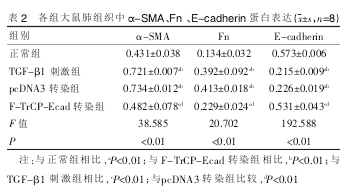
����2.4 ӫ��ʵʱ���� RT-PCR ��� α-SMA��col��col�����
������������Ƚ�, TGF-β1 �̼��顢pcDNA3 תȾ��β-catenin ;�� α -SMA,col��,col��mRNA ��������(P<0.01), ������Ƚ���ͳ��ѧ���� ,F-TrCP-EcadתȾ��� TGF-β1 �̼��� α-SMA��col��col����オ��(P<0.01),��������Ƚ���ͳ��ѧ���졣 ���� 3��
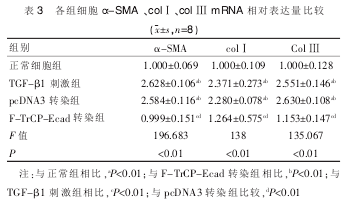
����3�� ����
�����γ���άϸ����ֳ�������ǿ�Ƿ���ά����������Ҫ���²�����,������Ƶ��о���Ѱ���µ����ưе���Ŀǰ�о��Ľ��㡣����ά���������� TGF-β1�̼���ʹ�γ���άϸ�����ϵ���ֳ��ת��Ϊ������άϸ�� (myofibroblast,MB)�� MB ��һ������ε�FB,�ܹ��������� ECM,���������� FB �� 4~5�� ,�������� α-SMA��Fn��I��III �ͽ�ԭ,������ά�����̡�
����TGF-β1 ���յ��γ���άϸ�������������Ҫ����������,TGF-β1 �� �����õ�;������ Smad��β-catenin �ͷ� Smad ͨ·, �� β-catenin ;���뼲������ά���γɵIJ������̸߶���ء� Vuga,et al���� WNT5A ����;���� IPF �ķγ���άϸ���������γ���άϸ���������ص�������, ͨ���Ǿ����WNT / β-catenin ; ��WNT5A �� ���������յ��γ���άϸ����ֳͬʱ����ϸ�������� Chilosi �ȱ��������ط��Է���ά��(IPF)��������֧���ܻ���ϸ�������˲�λ�γ���άϸ���пɼ� β-catenin ��ϸ������ת�ơ�����,��ʾ β-catenin �����ά�����γ���άϸ�����Ե������,�����������ͨ������ β-catenin���ܿ��ܶԷ���ά�������������á�
����F-TrCP-Ecad ������ Cong����Ƶ� β-catenin��������,ֻ����ϸ�����е� β-catenin ����,�����ƻ�ϸ��Ĥ�ϵ� β-catenin ����, �Ӷ���������ϸ�����ڵ�ת��,������ת¼���ԡ� ���о����ø�����תȾ�������άϸ��, �۲���� β-catenin ;���Էγ���άϸ����ֳ�����Ե�Ӱ�졣 �����ʾ:TGF-β1�̼� 48 h ��,����άϸ�������������,��ͻ��ʧ,ϸ��������������, ϸ����϶��С,F-TrCP-Ecad תȾ���ϸ��������,�нϳ���ͻ��,ϸ���������Լ�϶�� CCK-8 ����� F-TrCP-Ecad תȾ��γ���άϸ����ֳ�� TGF-β1 �̼������Խ��͡� F-TrCP-Ecad תȾ��γ���άϸ������ϸ����־�� α -SMA��Fn ���ױ��������Ե��� TGF-β1 �̼���,E-cadherin�������� TGF-β1 �̼������ߡ� F-TrCP-Ecad תȾ�����γ���άϸ�� β-catenin ;��ת¼���� α-SMA��col��col��������Խ���, ֤ʵ β-catenin �����ó�ͨ������ϸ�����е� β-catenin ����, ��������ϸ�����ڵ�ת������ TGF-β1 �յ��ķγ���άϸ����ֳ��������ǿ,������ά�����̡�
������������,TGF-β1 �Ǵٽ��γ���άϸ����ֳ�������Ҫϸ������,β-catenin ;���� TGF-β1 �������õ���Ҫ�ź�ͨ·,β-catenin �����ó�����(F-TrCP-Ecad ����)ͨ����ϸ��ź�;�� ,��ϳ���άϸ��ת��Ϊ�������ʽ������άϸ��,ͬʱ���Ƴ���άϸ���ͷŰ������,�����ά������,�Ӷ���Ϊ����ά�����������ṩ�µ�˼·��ʵ�����ݡ�

̽��Smad7��ActA/Smadsͨ·�ĵ��ػ���
�����أ����������������������������ת���������ӣ��������������������������������ԣǣƣ����������Ա֮һ������Ϊ����������ӣ������ź�ͨ·�ĵ�һ��ʹ��ͨ����������ϸ���ڣӣ���������Ӧ����ɾ���һ������ѧ��...
̽��IL-17A�ڷ���ά�����������е�����
�ط��Է���ά��(idiopathicpulmonaryfibrosis��IPF)����δ�����÷��ڳ��ˡ������ڷβ��������Խ�չ����Ѱ���ͼ����Է���Ϊ����������һ�ּ����Էμ�...
̽��IL-37�Ե��˾���ϸ������ĭ�������ü���
��������Ӳ�������ӣ���һ���ۼ�ȫ��Ѫ�ܵ����Բ��䡣���Ӳ�������֬�ʴ�л���ң�ͬʱҲ��һ��������֢�Լ�������������ϸ����ϸ�����Ӳ������䷢�����չ������ϸ��Ǩ�Ƶ���Ƥ��ת��Ϊ�����������õľ���ϸ����ͨ����������壨���������������...
̽��BK��TGF-��1�յ���PASMCs��ֳ������
�ζ�����ѹ(pulmonary��artery��hypertension��PAH)��һ�����صĽ�չ�Լ���������Ҫ�����Ƿ�Ѫ���������������ߣ�������չ�ɵ��»�������˥�߶�������1�ݡ���Ѫ���ع��Ƿ�Ѫ���������������ߵ���Ҫ���������������ζ���ƽ����ϸ��(pulmonaryarterialsmooth...
���ܸ��������ֶεĸĽ�,ȱѪ�����ಡ����Ŀǰȫ�������Լ�������Ҫ����ȱѪԤ����(IPC)������Ϊ��Ч���ļ�������ʽ,��IPC������ȱѪǰӦ��,����ٴ�Ӧ���ܵ����ơ��о���Ա����ȱѪ��������ͨ����СȱѪ�ļ��������,���Ѫ����Ƥϸ�����ܵ�;��������...
̽��ET-1�ܷ���������ļ����ɻ�����������
��Ƥ�أ�1(endothelin��1��ET��1)����Ѫ��ϵͳ��ET����Ҫ��ʽ��������Ƥϸ����Ѫ��ƽ����ϸ�����ļ�ϸ������ϸ�������ν���ϸ���Ⱥϳɡ�ET��1������ʧ������������˥�ߡ���������Ӳ���ȶ�����Ѫ�ܼ��������ж�������Ҫ���á������ɻ��ǻ����������ӵIJ���ȫ...
ȱѪ����(ischemicpostconditioning,IPO)��ͨ�����ļ����ٹ�ע���ھ����������ݵ�ȱѪ���ٹ�ע,�ܼ�������ʧ���ķ���,�����Ĺ���,��С�������,�����ļ�ϸ������[1]��Murry��[2]�����Ԥ������ͬ,IPO����ȱѪ���ٹ�ע֮ǰ���з������ݵ�ȱѪ���ٹ�ע��...
��������Ӳ��(atherosclerosis,As)��һ��������֢�Լ�...
̽��GDF-15��CRP�ڼ����Թ�����������Ѫ��ˮ
�����ֻ�����15��growthdifferentiationfactor-15,GDF-15����ת����������-��transforminggrowthfac-tor-��TGF-���������е�һԱ�����о���Ϊ��GDF-15����Ѫ�ܼ�����أ�GDF-15�Ƿ�ӳ��Ѫ�ܹ��ܺͼ����Ķ������ﻯѧ��־��[1].Ŀǰ�й���...
�ز�����X�������շ������Է��ķ�������̽
AIM2�ǽ�Щ�����·��ֵ�һ��ģʽʶ������(patternrecognitionreceptors,PRR),�ܹ�����DNA������ϸ����Ⱦʱ�ͷŵ�ϸ������dsD-NA[1],ͨ����Cĩ��HIN-200��ʶ��dsDNA��,��Nĩ�˵�pyrin����ASC��Nĩ�˵�pyrin�����,��ASC��Cĩ�˵�CARD��Caspase1ǰ���CARD��...