病理学论文


呼 吸 道 合 胞 病 毒 (respiratory syncytial virus,RSV)是引起 5 岁以下婴幼儿下呼吸道感染的主要病原[1]. 婴幼儿时期 RSV 感染与儿童持续喘息和气道高反应性(airway hyper-responsiveness,AHR)密切相关[2]. Thorburn 等[3]报道超过 40%的需要收入 ICU 的重症 RSV 感染患儿合并下呼吸道细菌感染, 且以能产生脂多糖(lipopolysaccharide,LPS)的革兰阴性菌为主,提示合并细菌感染,特别是革兰阴性菌,可能加重 RSV 感染. LPS 作为革兰阴性菌主要成分,在其致病性方面发挥重要作用.本课题组前期结果显示 LPS可通过 TLR4 (Toll-like receptor 4,TLR4) 上调 RSV感染后上皮细胞 IL-6 表达水平[4],但在体内 LPS 对RSV 感染的影响还不清楚. 本研究拟建立 RSV 感染后 LPS 再刺激的小鼠动物模型, 观察其气道炎症及AHR,并探讨气道炎症及 AHR 产生机制,为进一步寻找 RSV 感染后继发革兰阴性菌感染的治疗靶点奠定理论基础.
1 材料与方法
1.1 病毒准备 呼吸道合胞病毒 A2 标准株来源于北京儿童医院病毒实验室, 将其接种于 Hep-2 单层细胞,37 ℃条件下培养于含 10%胎牛血清的 DMEM 培养基中,细胞出现 90%以上病变时刮起贴壁细胞,收集混悬液,12 000 r/min 4 ℃离心 10 min, 收集上清,分装病毒液,-80 ℃保存备用, 空斑实验测定病毒滴度. 未接种病毒的细胞培养液为病毒对照液.
1.2 实验动物及分组 SPF 级 6~8 周龄雌性 Balb/c 小鼠由重庆医科大学实验动物中心提供, 饲养于 IVC级环境中,独立呼吸通道,灭菌饲料及饮水喂养,所有动物实验操作获得重庆医科大学伦理委员会同意,遵守《实验动物管理条例》. 将小鼠随机分为 4 组. 对照组: 病毒对照液 100 μl 滴鼻后,50 μl PBS 连续 3 d滴鼻;LPS 组:病毒对照液 100 μl 滴鼻后,10 μg LPS(Escherichia coli 055:B5,Sigma)/50 μl PBS 连续 3 d滴鼻;RSV 组:病毒液 100 μl 滴鼻,相当于滴入病毒7×106PFU,50 μl PBS 连续 3 d 滴鼻;RSV+LPS 组:病毒液 100 μl 滴鼻后,连续 3 d 滴鼻给 10 μg LPS/50 μlPBS. 在首次处理后第 7 天收取肺组织、肺泡灌洗液标本.
1.3 病毒拷贝数检测 按 BioTake 快速全 RNA 试剂盒说明书提取小鼠肺组织总 RNA,紫外分光光度计定量后,TaKaRa PrimeScript RT reagent Kit 逆转录试剂盒,按说明进行逆转录. 取 2 μl cDNA 用探针法检测病毒拷贝数. 采用 TaqMan Universal PCR MasterMix(Applied Biosystems), 程序为:50 ℃ 2 min,95 ℃10 min, 95 ℃ 15 s,60 ℃ 1 min,39 个循环. 引物、探针、质粒参考文献设计[5],均由华大基因合成(表 1).

1.4 肺 泡 灌 洗 液 (bronchoalveolar lavage fluid,BALF) 中炎症细胞总数及分类计数 成功气管插管后,4 ℃预冷 PBS 3 ml 灌洗肺脏,0.5 ml/次,共 6 次,回收率大于 90%. 灌洗液于 4 ℃ 2 500 r/min 离心 5 min后,收集上清,-80 ℃保存备用. 1 ml PBS 重悬细胞沉淀,显微镜下计数,剩余液体再次离心后将细胞沉淀涂片,快速瑞姬氏染液(南京建成科技有限公司)染色后,光学显微镜下分类计数. 计数时为实验者随机选取组别及小鼠编号后从 1 开始编号, 编号完成后,与另一实验者分别计数,2 人计数平均值为所得细胞计数及分类计数结果.
1.5 肺组织病理切片 将小鼠麻醉处死后,取出肺组织,置 4%甲醛固定 24 h 后,行冲水、脱水、组织透明、浸蜡、包埋.组织蜡块切片后行 HE 染色,光镜下观察肺组织受损及炎症浸润情况,根据文献[6]分别进行细支气管周围炎症、血管周围炎症及肺泡炎症病理评分. 评分时为实验者随机选取组别及小鼠编号后从 1开始编号,编号完成后,与另一实验者分别评分,2 人评分平均值为所得病理评分结果.
1.6 气道高反应性测定 小鼠气道高反应性在清醒状态下检测, 通过体积描述法检测小鼠自主呼吸.Baseline 为小鼠在未进行雾化给药前,体积描记仪器(EMKA)记录气道阻力记录的 Penh 值. 待检测的小鼠用生理盐水或不同质量浓度 (3.125、6.25、12.5、25及50 mg/ml)的乙酰甲胆碱(Sigma)雾化 3 min,休息 2 min,然后记录 Penh 值,连续记录读数 5 min.气道高反应性曲线表示为在不同质量浓度乙酰甲胆碱雾化后的气道阻力(enhanced pause, Penh),Penh=PEP/PIF×Pause.
1.7 肺 组 织 TRIF、 MyD88 蛋 白 表 达 检 测 采 用Western blot 方法检测肺部 TRIF、 MyD88 蛋白表达.提取肺组织总蛋白,按凯基全蛋白抽提试剂盒说明书操作,BCA 法蛋白定量. 80 μg 蛋白上样, 经 10%SDS -PAGE 凝胶电泳 , 转膜 , 室温封闭 1 h, 一抗(TRIF-1 ∶500,MyD88-1 ∶200,β-action-1 ∶10 000)(TRIF -Abcam,MyD88 -Santa Cruz Biotechnology,β -action-Epitomics)4 ℃孵育过夜 ,二抗 (1∶10 000)(联科生物)室温孵育 1 h,ECL 发光液(Bio-rad)进行蛋白信号检测.
1.8 细胞因子水平检测 ELISA 方法检测小鼠 BALF中 KC、IFN-γ、IL-1β、IL-6 细胞因子水平, 操作按试剂盒(KC--R&D,IFN-γ、IL-1β、IL-6-四正柏)说明书进行.
1.9 统计学分析 统计学分析采用 Graphpad Prim5.03 进行数据统计分析,采用单因素方差分析(One-way ANOVA), 先进行方差齐性 Bartlett's test 检验,方差齐者组间差异使用 Newman-keuls 检验 ,方差不齐者组间差异使用 Kruskal-Wallis test 检验;P<0.05为差异有统计学意义.
2 结果
2.1 病 毒 拷 贝 数 RSV +LPS 组 (1.36 ×105±6.13 ×104copies RSV-RNA/ml)与 RSV 组 (1.06×105±5.38×104copies RSV-RNA/ml) 之间病毒拷贝数差异无统计学意义.
2.2 LPS 对 RSV 感染小鼠肺部炎症的影响2.2.1 BALF 中细胞总数及分类计数 LPS 组 、RSV组、RSV+LPS 组 BALF 中细胞总数较对照组均明显增多(P 均<0.05),且 RSV+LPS 组细胞总数增多最明显. LPS 组以中性粒细胞为主,RSV 组以淋巴细胞为主,RSV+LPS 组以中性粒细胞为主(图 1).
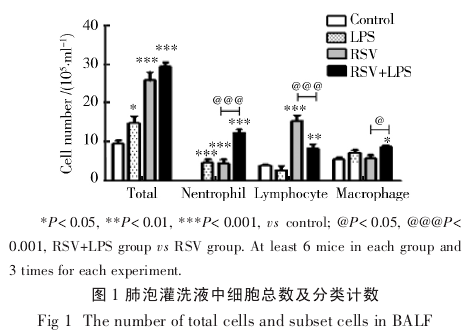
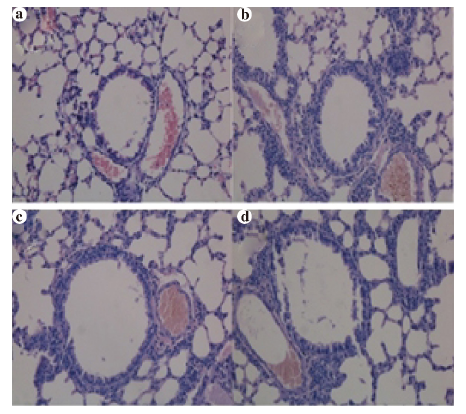
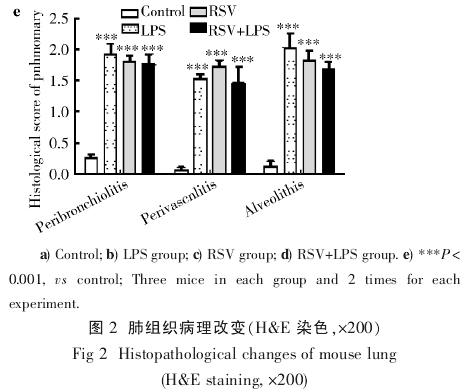
2.2.2 肺组织病理改变 对照组小鼠肺泡结构清楚,壁薄无充血,肺间质无炎症细胞浸润.LPS 组、RSV 组和 RSV+LPS 组均存在弥漫性肺泡间隔增厚与肺泡壁破坏,气管周围、血管周围不同程度炎症细胞浸润,3组气管周围、血管周围、肺泡间隔病理评分差异无统计学意义(图 2).
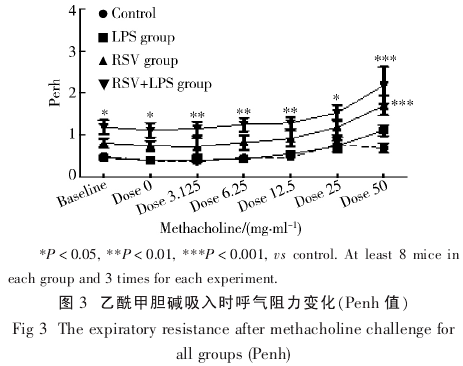
2.3 气道高反应性(Penh 值) LPS 组较对照组无气道高反应性,RSV 组、RSV+LPS 组气道高反应性较对照组明显增高(P 均<0.001). RSV+LPS 组气道高反应性较 RSV 组增高,但差异无统计学意义(图 3).
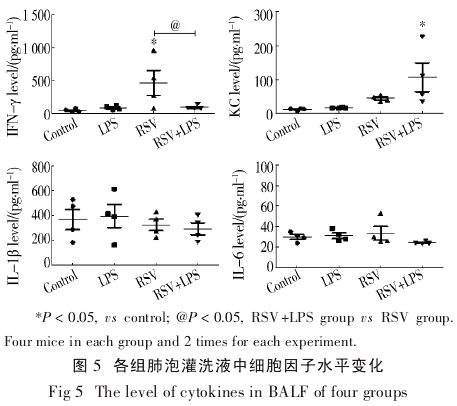
2.4 TRIF 与 MyD88 蛋白表达 LPS 组肺组织中TRIF 蛋白表达较对照组变化不明显;RSV 组中 TRIF点蛋白表达较对照组稍增高,差异无统计学意义;RSV+LPS 组 TRIF 蛋白表达较对照组明显增高 (P <0.01);RSV+LPS 组较 LPS 组 、RSV 组 TRIF 表达升高,差异无统计学意义.各组肺组织中 MyD88 蛋白表达无差异(图 4).
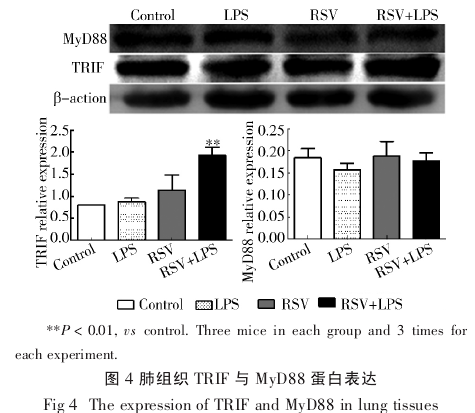
2.5 细胞因子检测 RSV 组小鼠 BALF 中以 IFN-γ升高为主(P<0.01),RSV+LPS 组以 KC 升高为主(P<0.05). 各组 BALF 中 IL-6、IL-1β 表达无明显变化(图 5).
3 讨论
全球范围内每年约有 3 380 万婴幼儿因RSV 感染而致下呼吸道感染, 其中 340 万患儿需要住院治疗,6.6~19.9 万患儿死亡[7]. 40%~50%的因感染 RSV而住院的患儿会发展成为持续性喘息[8],特别是重症RSV 肺炎患儿, 发展为反复喘息及成为儿童哮喘的风险更高[9]. Thorburn 等[3]报道超过 40%的需要收入ICU 的重症 RSV 感染患儿都伴随下呼吸道细菌感染. RSV 感染伴随继发性细菌感染因其对气道炎症的影响可能会加重喘息的严重性[10]. 本研究中,LPS刺激 RSV 感染小鼠后,气道炎症与 AHR 均增加,与课题组前期的体外研究结果[4]及临床现象一致.
病毒进入宿主,利用宿主细胞进行复制、扩增,对宿主造成直接损伤.Zhao 等[11]报道博卡病毒可以引起下呼吸道感染,但是只有博卡病毒高拷贝复制的患者才出现临床表现,提示高拷贝病毒复制和疾病严重性存在相关性. 且本课题组前期研究也发现 RSV 病毒复制可能与感染后气道炎症及 AHR 相关[12]. 本研究中 RSV+LPS 组较 RSV 组病毒拷贝数无增高,提示病毒高拷贝可能不是 RSV+LPS 组气道炎症及 AHR 较RSV 组加重的原因.
中性粒细胞是最先抵达炎症部位的天然免疫细胞,进行病原清除的同时也可引起炎症反应[13]. 本研究中,LPS 刺激 RSV 感染小鼠后使其 BALF 中炎症细胞由淋巴细胞为主转变为中性粒细胞为主. 同时,RSV+LPS 组小鼠 AHR 也较 RSV 组有增高, 提示中性粒细胞可加重 AHR,与研究报道 RSV 感染重症患儿气道以中性粒细胞为主[14]相符.
KC 是小鼠中性细胞最重要的化学趋化因子之一,它的功能相当于人类的 IL-8. 作为中性粒细胞强效趋化因子及化学激活剂,IL-8 可引起中性粒细胞向呼吸道迁移、活化[15],是重症 RSV 肺炎的危险因素之一[16]. 本研究中,RSV 组小鼠 BALF 中 IFN-γ 表达明显增加,与课题组前期报道 IFN-γ 表达可增加RSV 感染小鼠气道炎症及 AHR 相符[17]. RSV+LPS 组BALF 中以 KC 为主,提示 LPS 改变了单纯 RSV 感染BALF 中以 IFN-γ 为主的细胞因子表达,增加了 KC表达,BALF 中细胞因子水平与细胞计数及分类计数结果一致. 其可能机制是:LPS 做为革兰阴性菌的致病成分可促进中性粒细胞趋化因子 KC 在肺组织中的表达,从而肺泡灌洗液中中性粒细胞明显增多. 过多的中性粒细胞所引起的炎症反应可以引起组织损伤[13],从而使 RSV+LPS 组小鼠气道炎症及 AHR 较RSV 组增强.
Toll 样受体家族 (Toll-like receptor,TLRs)作为天然免疫的重要组成部分,通过识别多种病原微生物的病原分子相关模式 (pathogen molecular associatedpattern,PMAP)在急性炎症反应和信号转导方面起重要作用[18]. RSV 感染 9HTEo 气道上皮细胞后 TLR1-10 mRNA 水平均上调[4]. TLRs 通过下游髓样分化因子 88(MyD88)依赖途径和 MyD88 非依赖途径(主要是 TRIF)进一步发挥作用.RSV 的免疫反应特别是天然免疫与病毒成分 F 蛋白、G 蛋白关系密切, 而目前研究较清楚的是 F 蛋白通过 TLR4 发挥作用.文献报道正常气道上皮细胞对 LPS 暴露无反应, 当 RSV 感染该气道上皮细胞后,TLR4 出现膜表达增高而胞内表达降低的重新分布现象[19-20].因此,在本研究中可能由于 RSV 感染后上皮细胞膜 TLR4 表达升高对 LPS的耐受机制被破坏,TRIF-KC 通路活化,下游炎症因子水平上调. 另一方面,LPS 或 RSV 干预后 MyD88水平存在动态变化, 本研究仅对感染后第 7 天进行研究,未检测到 MyD88 升高.以上可能是 LPS 组、RSV+LPS 组肺组织中 TRIF 存在表达差异而 MyD88 变化不明显的原因.
综上所述,RSV+LPS 组小鼠气道炎症及 AHR 较RSV 组增加可能是通过 TRIF-KC 途径募集中性粒细胞,造成肺组织损伤所致. 在环磷酰胺处理的免疫低下的小鼠中,RSV 通过 TRIF-IFN-γ 引起气道炎症及 AHR[17],而在 LPS 刺激 RSV 感染小鼠后,也是通过增强 TRIF 来增加气道炎症及 AHR,提示 TRIF 在RSV 导致的炎症中扮演重要角色, 但是它是如何引起下游不同细胞因子的表达, 从而加重气道炎症及AHR 需要进一步研究.
【参考文献】
[1] Escobar GJ, Masaquel AS, Li SX, et al. Persistent recurringwheezing in the fifth year of life after laboratory-confirmed,medically attended respiratory syncytial virus infection ininfancy[J]. BMC Pediatr, 2013, 13(1): 97.
[2] Blanken MO, Rovers MM, Molenaar JM, et al. Respiratorysyncytial virus and recurrent wheeze in healthy preterminfants[J]. N Engl J Med, 2013, 368(19): 1791-1799.
[3] Thorburn K, Harigopal S, Reddy V, et al. High incidence ofpulmonary bacterial co -infection in children with severerespiratory syncytial virus (RSV) bronchiolitis[J]. Thorax, 2006,61(7): 611-615.
[4] Xie X, Law H, Wang L, et al. Lipopolysaccharide inducesIL -6 production in respiratory syncytial virus -infectedairway epithelial cells through the toll -like receptor 4signaling pathway[J]. Pediatr Res, 2009, 65(2): 156-162.
[5] Hu A, Colella M, Tam JS, et al. Simultaneous detection,subgrouping, and quantitation of respiratory syncytial virusA and B by real-time PCR[J]. J Clin Microbiol, 2003, 41(1):149-154.
[6] Peebles RS, Sheller JR, Collins RD, et al. Respiratorysyncytial virus infection does not increase allergen-inducedtype 2 cytokine production, yet increases airwayhyperresponsiveness in mice[J]. J Med Virol, 2001, 63(2):178-188.
[7] Nair H, Nokes DJ, Gessner BD, et al. Global burden ofacute lower respiratory infections due to respiratorysyncytial virus in young children: a systematic review andmeta-analysis[J]. Lancet, 2010, 375(9725): 1545-1555.
[8] Mandelberg A, Tal G, Naugolny L, et al.Lipopolysaccharide hyporesponsiveness as a risk factor forintensive care unit hospitalization in infants with respiratorysyncitial virus bronchiolitis [J]. Clin Exp Immunol, 2006,144(1): 48-52.
[9] Bacharier LB, Cohen R, Schweiger T, et al. Determinants ofasthma after severe respiratory syncytial virusbronchiolitis[J]. J Allergy Clin Immunol, 2012, 130(1): 91-100.
[10] Deng Y, Liu W, Luo ZX, et al. Impact of bacterialcolonization on the severity, and accompanying airwayinflammation, of virus -induced wheezing in children [J].Clin Microbiol Infect, 2010, 16(9): 1399-1404.
[11] Zhao B, Yu X, Wang C, et al. High human bocavirus viralload is associated with disease severity in children underfive years of age[J]. PLoS One, 2013, 8(4): e62318.
[12] 臧 娜, 谢晓虹, 邓 昱, 等. 白藜芦醇对呼吸道合胞病毒感染小鼠肺部病毒滴度及其气道炎症的影响[J]. 第三军医大学学报, 2011, 33(18): 1924-1927.
[13] Amulic B, Cazalet C, Hayes GL, et al. Neutrophil function:from mechanisms to disease [J]. Annu Rev Immunol, 2012,30: 459-489.
[14] Collins PL, Melero JA. Progress in understanding andcontrolling respiratory syncytial virus: still crazy after allthese years[J]. Virus Res, 2011, 162(1): 80-99.
[15] 吴大玮, 姚海燕. 慢性阻塞性肺疾病患者诱导痰 IL-8、TNF-α 水平和炎症细胞的分布[J]. 中国老年学杂志, 2007,27(14): 1366-1369.
[16] Wu P, Hartert TV. Evidence for a causal relationshipbetween respiratory syncytial virus infection and asthma[J].Expert Rev Anti Infect Ther, 2011, 9(9): 731-745.
[17] Zang N, Xie XH, Deng Y, et al. Resveratrol -mediatedgamma interferon reduction prevents airway inflammationand airway hyperresponsiveness in respiratory syncytialvirus-infected immunocompromised mice[J]. J Virol, 2011,85(24): 13061-13068.
[18] Klouwenberg PK, Tan L, Werkman W, et al. The role ofToll -like receptors in regulating the immune responseagainst respiratory syncytial virus [J]. Crit Rev Immunol,2009, 29(6): 531-550.
[19] Monick MM, Yarovinsky TO, Powers LS, et al. Respiratorysyncytial virus up -regulates TLR4 and sensitizes airwayepithelial cells to endotoxin[J]. J Biol Chem, 2003, 278(52):53035-53044.
[20] 谢晓虹, 刘恩梅, 杨锡强, 等. 呼吸道合胞病毒感染后气道上皮细胞 Toll 样受体 4 的表达及其功能[J]. 中华结核和呼吸杂志, 2008, 31(3): 213-217.

激活素A(ActivinA,ActA)是转化生长因子(transforminggrowthfactor-,TGF-)超家族成员之一,其作为ActA/Smads信号通路的第一信使,通过触发下游细胞内Smads级联反应,完成具有一定生物学功...
特发性肺纤维化(idiopathicpulmonaryfibrosis,IPF)病因未明、好发于成人、局限于肺部、进行性进展,以寻常型间质性肺炎为病理特征的一种间质性肺疾...
动脉粥样硬化(AS)是一种累及全身血管的慢性病变。AS不仅仅是脂质代谢紊乱,同时也是一种慢性炎症性疾病,多种免疫细胞及细胞因子参与了其发生与进展。单核细胞迁移到内皮下转化为具有吞噬作用的巨噬细胞,通过清道夫受体(scav-engerre...
肺动脉高压(pulmonary、artery、hypertension,PAH)是一类严重的进展性疾病,其主要特征是肺血管阻力进行性升高,持续发展可导致患者右心衰竭而死亡[1]。肺血管重构是肺血管阻力进行性升高的重要病理生理基础,肺动脉平滑肌细胞(pulmonaryarterialsmooth...
尽管各种治疗手段的改进,缺血性心脏病仍是目前全球致死性疾病的首要病因。缺血预处理(IPC)被公认为有效的心肌保护方式,但IPC必须在缺血前应用,因此临床应用受到限制。研究人员发现缺血后处理可以通过减小缺血心肌梗死面积,提高血管内皮细胞功能等途径发挥心...
内皮素-1(endothelin-1,ET-1)是心血管系统中ET的主要形式,可由内皮细胞、血管平滑肌细胞、心肌细胞、肝细胞和星形胶质细胞等合成。ET-1在心律失常、慢性心力衰竭、动脉粥样硬化等多种心血管疾病发病中都起着重要作用。氧自由基是机体内氧分子的不完全...
缺血后处理(ischemicpostconditioning,IPO)即通过在心肌的再灌注早期经过反复短暂的缺血与再灌注,能减少心律失常的发生,改善心功能,缩小梗死面积,减少心肌细胞凋亡[1]。Murry等[2]提出的预处理不同,IPO是在缺血后、再灌注之前进行反复短暂的缺血与再灌注。...
动脉粥样硬化(atherosclerosis,As)是一个慢性炎症性疾...
生长分化因子15(growthdifferentiationfactor-15,GDF-15)是转化生长因子-(transforminggrowthfac-tor-,TGF-)超家族中的一员。有研究认为,GDF-15与心血管疾病相关,GDF-15是反映心血管功能和疾病的独立生物化学标志物[1].目前中国关...
AIM2是近些年来新发现的一种模式识别受体(patternrecognitionreceptors,PRR),能够感受DNA病毒或细菌感染时释放到细胞浆的dsD-NA[1],通过其C末端HIN-200区识别dsDNA后,其N末端的pyrin区与ASC的N末端的pyrin区结合,而ASC的C末端的CARD与Caspase1前体的CARD区...