����ѧ����


�������ܶ���ı�̬���������ܼ�״�ټ��ص���,������ܶ���һֱ����Ϊ���о���״�ټ��ع��ܺ��ź�ͨ·��ģʽ����[1, 2].�������ǶԻ�����Ⱦ���״�ٸ������õ�������ע[3, 4], �������ܶ����ֱ����ڼ�״�ٸ��ŵ��о�[5-8], �����ͨ���ַ���צ�[9, 10]�����ô��ţ�ܺͱ���[11, 12]��ŷ������[13, 14]���ձ��Ĵ�Ƥ��[15, 16]��.����Ŀǰ�ӹ�����ڷ���צ����һЩ����, ��չ�ҹ��������ܶ����ֵ��о������ش�, ����Ϊ��״�ٸ��ż�����������о��ṩ�����������.
�����ڰ���(Rana nigromaculata)���ҹ�������һ�����ܶ�����, ����ѧ�߶Ժڰ��ܵ�����ѧ[17-20]�Ͷ���ѧ[21, 22]�й�һ���о�.Ȼ��Ŀǰ��û�й��ںڰ��ܱ�̬���������м�״�ٷ����ͼ�״�ټ��ط��ڵı���, Ҳû�кڰ��ܱ�̬������������̬������.��Щ��Ϣ�ǿ�չ�ڰ��ܼ�״�ٸ����о�������ѧ����.����ϵͳ�о��˺ڰ��ܱ�̬����������������̬����״����֯ѧ�Լ���״�ټ���ˮƽ�ı仯,���ƽ��ڰ����ڼ�״�ٸ����о��е�Ӧ��.
����1 �����뷽��
����1.1 �ڰ��ܺ���������
�����ڰ��ܳ��������ڶ��﷿ˮ½���ܵIJ�������,ÿ����ϴ, ˮ��(23±2) �� ,����������Ϊ 12h �� 12h.����ÿ��ιʳ���Ӻ��ܲ�������� 1 ��.����������ܸ�ע�� 15 μg �ٻ������ͷż��� A3(LHRH-A3)������ëĤ�����ټ���[���� 300 ���ʵ�λ(IU), ���� 200 IU], ���ּ��ؾ�����������Ϊ 0.6%��������ˮ����, �������ܶ����յ������豸��, �յ��䱧�Բ���.�ܾ��ѷ���ȥ��ˮ�з���, 4d ����������.
���������ת����������(100 cm×60 cm×30 cm) ����, ˮ�� 15 cm, ÿ��ιʳ����(�������)����.ÿ������ܶ�ÿ��ˮ�в����� 5 ֻ���(Gosner 30��[23]֮ǰ), 30��֮��ÿ��ˮ������1.5ֻ���.
�������������ÿ�ܻ�ˮ����, ˮ��������������: ����, pH 6.5-7.0, �ܽ������� 5 mg/L, Ӳ��(CaCO3)Լ 150 mg/L.�����ܶ��﷿��, �¶� (23±2)�桢������ �� �� Ϊ 12h �� 12h(��� �� ˮ �� �� ǿ �� 600-1000 mol/(m2s), ���ܸ�ǿ��100-300 mol/(m2s),�������� Gosner 26-46 ��[23]ȡ��.
����1.2 ��̬ѧ�۲�ͽ�ʶ��
��������Gosner[23]����βĿ���Ľλ���ϵͳ�Ժڰ��������ν��л���.ȡ 26-46 �������������̶��ڹ̶�Һ[�̶�Һ�䷽Ϊ��ζ��U��ȩ�U������(�����)=75�U25�U5]��, ֱ�����۹۲�����̬, ����֫������ʽ�����¹۲�(�Ŵ���Ϊ 1-4��).ÿ��������������,���ص�۲��Ȳ��仯.��������, ������̬ѧ�������Թ۲�����.
����1.3 ��״����֯ѧ���
������ 26-46 �������������, ȡ������̶�Һ��,ÿ��������ȡ��ֻ���.����֯�ӹ̶�Һ��ȡ����, ��ˮ24h; ���� 70%��80%��95%��100% �ƾ�Ũ���ݶ���ˮ�� ,���� 85%��95%�� 100%���ױ�����.��������������������Ƭ.��Ƭ��� 5 μm, Ȼ��������HE Ⱦɫ����Ƭ��,�ڹ�ѧ�����¹۲졢����,�Ŵ���Ϊ 10×Ŀ��, 10×�ᄉ.����������з���, �۲����ݡ�����ϸ����������̬����ɫ����С��.
����1.4 ��״�ټ�����ȡ���ⶨ
�������ڰ��� 32/33-46 ����������ȡ����, ���� 1.5 mL��ˮ�Ҵ���, �������� 1 �����������.���Ƚ�������ĥ2min.���ڳ������г��� 30s,Ȼ������15min, ��ȡ����Һ.
�����ڳ����м��� 0.3 mL ����ˮ�Ҵ�, ���� 30s ��, ���� 5min,�ϲ�����.����������Һ��37��ˮԡ������, ���� 0.5 mL PBS ����Һ, ����ܽ��, ��ȫ�Զ���ѧ�������߷�����(CLIA)�ⶨ T3����.
����2 ���
����2.1 �ڰ�����������̬ѧ�仯
�����ڰ������ 26-46 ���ε���̬ѧ�仯����(ͼ 1): 26-30 ����Ҫ����Ϊ��֫������, ��Ϊ��֫ѿ������: 26 ��Ϊ��֫ѿ�ոճ���, ���Ȳ�����ȵ�һ��; 27 �εĺ���ѿ���ȳ������ȵ�һ�뵫С�ڿ���; 28 ��ʱ����ѿ�����Գ�������;29/30 ��, ��֫ѿ���ȿɴ���ȵ� 1.5-2 ��.

����31-36 ��, Ϊֺ�ֻ���: 31 ��ʱ, ��֫ѿ�ʽ�״; 32/33 ��, ��֫��������ȱ��; 34��35 ��, ȱ������Ϊ�������ĸ�; ���� 36 ��ʱ, ǰ����ȱ������, �ֻ�����ֺ; 37 ��ʱ�ĸ�ȱ��������, �ֻ������Ե���ֺ.38-40 �γ�����ͻ�ؽ�����,38 ��, ��һֺ��������ͻ; ���� 40 ��, ��ֺ��������ֹؽ�����.40 �κ����������̬��,���Ӿ��ҵ��α俪ʼ.41 ��, ����ǰ֫���Ƥ������, йֳǻ����ʧ����ʧ; 42 ��, ǰ֫չ��, ����һ�ο�ʼ, �ڽǿ�ʼ���.43 �ο��Ѽ���, �Ӳ��濴�ڽdz����ǿ�, ��δ�ﵽ��,ͬʱ����β��ʼ����.44 �ο��ѽ�һ������, �ڽǴﵽ���в�ˮƽ, ����ȫ����.45 �οڽǴﵽ�ۺ�ˮƽ, βή�����ն�������ʧ, ֻ��һ��Сͻ��.46 ��β��ȫ����, ��֫����, �����߱���������Ƶ�ɫ��, ��ζ��������̬�������.
����2.2 �ڰ��������μ�״����֯ѧ�仯
������ͼ2��ʾ, �����ĵ�һ����(26��), ��״�������С, ���Ѿ������ݴ���, �������洦���Կ����ɵ�����Ƥϸ����ɵ� 1-2 �����ݽṹ,�м��н��ʴ���.�����ı�̬����������, ��27-37 ��ʱ, ��״�������������, ���ݸ���������, �������Ҳ�ڲ�������, ���ͬʱ, �����ڽ��ʺ���Ҳ�ڻ�������.�� 38 ��, ��״�����ͻȻ���, ������ĿҲѸ������, �����ڵĽ�����������, ���������Ӧ����.�������� 39-41 ��,��״�������������Ŀ�����������������.42 �μ�״�������������Ŀ�����������������, ��43 �μ�״�ٷ����ﵽ����, ����Ϊ��״��������������Ŀ��ࡢ����������.֮��� 44-46��, ��״������С, ������Ŀ���١������������, ������״��ò�����.
����
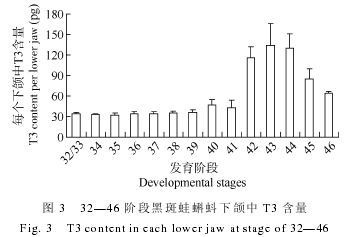
����2.3 �ڰ��ܷ��������� T3 ����ˮƽ�仯
�����ڰ��ܷ������������ T3 ��ˮƽ��ͼ 3 ��ʾ,����仯������: �ڱ�̬ǰ�ķ�������, T3 ������������, �����̬�ں�, T3 ����Ѹ������, �ڱ�̬�߷��ڴﵽ��ֵ, ��������½�.�������, ��32/33 ��, �ڰ��ܵ�������Ѿ����Լ��� T3,ÿ������к�����Լ�� 34 pg.�� 34-41 ��, ����� T3 ������������, ÿֻ����� T3 ������ 34 pg���ӵ� 45 pg ����.�� 42 �ο�ʼ, ����� T3 ����Ѹ������, ÿֻ����� T3 �������� 110 pg.�� 43��, T3 �����ﵽ��߷�, ÿֻ����� T3 �ķ�ֵˮƽ�� 130 pg ����.44 �μ�״�ټ��غϳɿ�ʼ����,ʹ��״���� T3 ����ˮƽ����.��̬��ɺ�(46��), ����� T3 ����ά����һ��ˮƽ.
����3 ����
����Gosner ��������̥�������εĻ���ϵͳ���㷺������βĿ������ε�ʶ��.���İ���Gosner ��ϵͳ, �Ժڰ��ܷ��������е���̬�仯������ϵͳ����.��������, �ڰ��ܵ���̬�仯��Gosner ϵͳ�е���̬��������һ��, ��������ϸ��ͬ: Gosner ������ѿ����Ϊ���ȵ� 1.5 ���� 2 ������Ϊ 29 ��; �����ȴﵽ���ȵ� 2 ��, ����Ϊ 30��; ����ѿ���ֽ�״��, ����Ϊ 31 ��.�������ǹ۲�ڰ��ܷ���, ����ѿ�ij�һ�㵽���˿��� 2��ʱ�ͻ���ֽ�״��, Ҳ���� 30 �β�����, �������ǽ��ڰ��ܵ� 29/30 �κϲ�.����, Gosner ����32 �� 33 �ηֱ�Ϊ��״���ϳ��� 1 ���� 2 ��ȱ��;���ڰ����� 2 ��ȱ������ͬʱ����, ��������Ҳ��32/33 �ϲ�.
�������ܶ���ı�̬������Ҫ�����ڼ�״�ٷ��ڵļ�״�ټ���.�����о��˺ڰ��ܴ�����һ���ε����һ�����ڼ��״�ٵķ���.�������, �ڰ��������ĵ�һ���μ� 26 �ξͳ��ְ������ʵļ�״������.֮ǰ��������ʾ, ����צ�ļ�״�������� Nieuwkoop and Faber (NF)�λ���ϵͳ�� 49-50 �βſ�ʼ����, �� 51 ��ʱ�������ʵ����������γ�[24].�ɴ˿���, �ڰ��ܵļ�״�����ݳ���ʱ��ȷ���צ�Ҫ��.�Ӽ��ط��ڵ�ʱ������, ������ 32/33 �εĺڰ�������м���� T3�Ĵ���.������������Ϊ, ����צ��� NF 54 ��ʱ��״�ٲſ�ʼ�ϳɷ��ڼ�״�ټ���[24].�����ڵ��о���Ϊ, ���ڷ���צ����ļ�״��Ҳ���кϳɼ�״�ټ��صĹ���, ���߷��������о�֮���Ա�������צ��״���� NF54 �ſ�ʼ���ڼ���, ��������Ϊ��ʱ�ļ�ⷽ��������������[25].�����צ�NF54 �ζ�Ӧ�ĺڰ��ܽ�Ϊ 34 ��, ��������� T3 ˮƽ��ǰһ�������û�����Ե���������, ��Ȼ 34 �εļ�״�����Ա� 32/33 �εļ�״�ٴ�, ��״����֯ѧ�ṹҲ���Է���.�� 38 ��ʱ�ڰ��ܵļ�״����֯ѧ�ṹ����һ�ν�Ϊ���Եķ���, ����Ϊ������Ӻ�������Ŀ����, ��Ӧ��״����֯�ķ���, T3 ��ˮƽҲ���ֻ�����������.��Ӧ��״�ٵĽ�һ������, T3 ˮƽ�� 40 �κ� 41 ����������.42 ��ʱ��״������Ѹ������ 20 ������, T3 ˮƽ��֮��������.�� 43 �� T3 ˮƽ�ﵽ����, ��ʱ�ļ�״����֯�ṹ��Ϊ����, ������Ŀ��� 30 ������.44 ��ʱ��״�ٿ�ʼ�˻�, ��ӦT3 ˮƽ����.֮�� 45 �κ� 46 ��, ��״�ټ����˻�, ��Ӧ T3 ˮƽѸ�ٻ���.������������������,�ڰ��ܼ�״�ټ���ˮƽ�ı仯���״����֯ѧ�ı仯��һ�µ�.
�������ݼ�״�ټ��صĿ�ʼ���ڡ�ˮƽ���ߡ������Լ���Ӧ����̬ѧ�仯, һ�㽫���ܶ���ı�̬��Ϊ����ʱ��: ��״�ټ��ز����ڻ��������ڵ�Ԥ��̬��(Pre-metamorphosis), ��Ӧ��ѿ�ķ���������; ��״�ټ��ط��ڵ��Ա���һ����ˮƽ״̬��ǰ��̬��(Pro-metamorphosis), ��Ӧֺ�ķֻ����ȵĿ��ٷ���; ��״�ټ���ˮƽ����ֱ����̬��ɵı�̬�߷���(Metamorphic climax), ��Ӧ����β�����ա��������ܵȾ��ҵı仯.�Ӽ���ˮƽ���о���������, �ڰ��ܷ��������в�û��һ��T3�����еĽ�, �� 34 �θ���Ҳû��һ����״�����������Ĺյ�, �������Dz��ܸ��ݼ�״�ټ��صı仯��ȷ��Ԥ��̬�ں�ǰ��̬�ڵĽ���.��Ϊ���о������п������������������������צ�Ƚ�, ���Dzο�����צ� 54 ����Ϊǰ��̬�ڵĵ�һ����, ������ȷ�����֮��Ӧ�ĺڰ��ܵ� 34 ����Ϊǰ��̬�ڵĵ�һ����.�� 42 ��, �ڰ��ܵ� T3 ˮƽ��������, ��Ӧ����̬�������ҵı仯, ��ǰ��չ�����ڽDZ��������ʼ���յ�, ��������̬�߷��ڵĿ�ʼ.���, �ڰ��ܱ�̬������ʱ�ڿ������»���: Ԥ��̬��Ϊ26-33��, ǰ��̬��Ϊ34-41��, ��̬�߷���Ϊ 42-46 ��.
�����ۺ����Ϲ���, ���Ľ�Ϊϵͳ���о��˺ڰ��ܱ�̬����������������̬�仯, �����˼�״����֯ѧ����ͼ��, �����˼�״�ټ���ˮƽ�ı仯, Ϊ���ڰ��ܼ�״�ټ��ع��ܺͼ�״�ٸ����о��춨����.
���������:
����[1] Yen P M. Physiological and molecular basis of thyroidhormone action [J]. Physiological Reviews, 2001, 81(3):1097-1142
����[2] Bernal J, Guada o-Ferraz A, Morte B. Perspectives in thestudy of thyroid hormone action on brain development andfunction [J]. Thyroid, 2003, 13(11): 1005-1012
����[3] Brucker-Davis F. Effects of environmental syntheticchemicals on thyroid function [J]. Thyroid, 1998, 8(9):827-856
����[4] Zoeller R T. Environmental chemicals impacting the thyroid:targets and consequences [J]. Thyroid, 2007, 17(9): 811-817
����[5] Tata J R. Amphibian metamorphosis as a model for thedevelopmental actions of thyroid hormone [J]. Molecularand Cellular Endocrinology, 2006, 246(1): 10-20
����[6] Heimeier R A, Das B, Buchholz D R, et al. Studies onXenopus laevis intestine reveal biological pathwaysunderlying vertebrate gut adaptation from embryo to adult [J].Genome Biology, 2010, 11(5): k55
����[7] Das B, Matsuda H, Fujimoto K, et al. Molecular and geneticstudies suggest that thyroid hormone receptor is bothnecessary and sufficient to mediate the developmental effectsof thyroid hormone [J]. General and ComparativeEndocrinology, 2010, 168(2): 174-180
����[8] Organisation for Economic Co-operation and Development(OECD). Test No. 231: Amphibian Metamorphosis Assay,OECD Guidelines for the Testing of Chemicals, Section 2[S]. Paris: OECD, 2009
����[9] Degitz S J, Holcombe G W, Flynn K M, et al. Progresstowards development of an amphibian-based thyroidscreening assay using Xenopus laevis Organismal and thyroi-dal responses to the model compounds 6-propylthiouracil,methimazole, and thyroxine [J]. Toxicological Sciences,2005, 87(2): 353-364
����[10] Heimeier R A, Das B, Buchholz D R, et al. Thexenoestrogen bisphenol A inhibits postembryonic vertebratedevelopment by antagonizing gene regulation by thyroidhormone [J]. Endocrinology, 2009, 150(6): 2964-2973
����[11] Helbing C C, Ovaska K, Ji L. Evaluation of the effect ofacetochlor on thyroid hormone receptor gene expression inthe brain and behavior of Rana catesbeiana tadpoles [J].Aquatic Toxicology, 2006, 80(1): 42-51
����[12] Croteau M C, Davidson M, Duarte-Guterman P, et al.Assessment of thyroid system disruption in Rana pipienstadpoles chronically exposed to UVB radiation and 4-tert-octylphenol [J]. Aquatic Toxicology, 2009, 95(2):81-92
����[13] Arukwe A, Jenssen B M. Differential organ expressionpatterns of thyroid hormone receptor isoform genes in p,p′-DDE-treated adult male common frog, Rana temporaria[J]. Environmental Toxicology and Pharmacology, 2005,20(3): 485-492
����[14] Gutleb A C, Appelman J, Bronkhorst M, et al. Effects of oralexposure to polychlorinated biphenyls (PCBs) on thedevelopment and metamorphosis of two amphibian species(Xenopus laevis and Rana temporaria ) [J]. Science of theTotal Environment, 2000, 262(1): 147-157
����[15] Hanada H, Katsu K, Kanno T, et al. Cyclosporin A inhibitsthyroid hormone-induced shortening of the tadpole tailthrough membrane permeability transition [J]. ComparativeBiochemistry and Physiology Part B: Biochemistry andMolecular Biology, 2003, 135(3): 473-483
����[16] Oka T, Miyahara M, Yamamoto J, et al. Application ofmetamorphosis assay to a native Japanese amphibian species,Rana rugosa, for assessing effects of thyroid systemaffecting chemicals [J]. Ecotoxicology and EnvironmentalSafety, 2009, 72(5): 1400-1405
����[17] Park S, Park S H, Ahn H C, et al. Structural study of novelantimicrobial peptides, nigrocins, isolated from Rananigromaculata [J]. FEBS Letters, 2001, 507(1): 95-100
����[18] Sumida M, Kanamori Y, Kaneda H, et al. Complete nucleotidesequence and gene rearrangement of the mitochondrialgenome of the Japanese pond frog Rana nigromaculata [J].Genes & Genetic Systems, 2001, 76(5): 311-325
����[19] Sumida M, Kato Y, Kurabayashi A. Sequencing and analysisof the internal transcribed spacers (ITSs) and coding regionsin the EcoR I fragment of the ribosomal DNA of theJapanese pond frog Rana nigromaculata [J]. Genes &Genetic Systems, 2004, 79(2): 105-118
����[20] Takase M, Ukena K, Tsutsui K. Expression and localizationof cytochrome P45011β, aldo mRNA in the frog brain [J].Brain Research, 2002, 950(1): 288-296
����[21] Li X Y, Zhou J, Yu M, et al. Toxic effects of 1-methyl-3-octylimidazolium bromide on the early embryonic develo-pment of the frog Rana nigromaculata [J]. Ecotoxicologyand Environmental Safety, 2009, 72(2): 552-556
����[22] Wang M Z, Jia X Y. Low levels of lead exposure induceoxidative damage and DNA damage in the testes of the frogRana nigromaculata [J]. Ecotoxicology, 2009, 18(1): 94-99
����[23] Gosner K L. A simplified table for staging anuran embryosand larvae with notes on identification [J]. Herpetologica,1960, 16(3): 183-190
����[24] Nieuwkoop P D, Faber J. Normal Table of Xenopus laevis(Daudin). A Systematical and Chronological Survey of theDevelopment from the Fertilized Egg till the End ofMetamorphosis [M]. Garland Publishing Inc. 1956, 22
����[25] Marsh-Armstrong N, Cai L, Brown D D. Thyroidhormone controls the development of connectionsbetween the spinal cord and limbs during Xenopus laevismetamorphosis [J]. Proceedings of the National Academyof Sciences, 2004, 101(1): 165-170

��(fluorene)���������һ���������(USEPA)���ȿ��Ƶ�16�ֶ����(polycyclicaromatichydrocar-bons��PAHs)֮һ����һ�ֽ���2����3��֮��Ķ��������Ҫ��Դ��ʯ�͡�ľ�ġ�ú̿�ȵ�ȼ�պͺ���ʯ�Ϳ��������е�й©���̵ȶ�����ᱻˮ���е�...

�о����鶯���������߲�ķ������������߲��
�����γɹ�����, ��̥������ֻ�Ϊ�������߲����ϸ����ϸ�������߷�����, �������߲��ַֻ�Ϊ��������ͱ������㡣�������߲�ķֻ��Բ��鶯��������̥����������Ҫ��...
��̥�����ڲ��ָ���ʼ�ڷ��ҹ�����Ĥ����γ����ںϣ������ҹָܷ�Ϊ���ҷ��ҿף�����Ĥ��֯�����γɷ��Ұ�[1,2].���⣬���ҹ�����Ĥ��Ҳ�����ķ������ҵķָ������ҹ�����Ĥ�����ķ�ԭ�������ı�������ͻ���ں��쳣�ɵ��·����ȱ���Ҽ��Ĥ��ȱ���...
����ļ����(Intermuscularbone)���ɼ��������֯����ͬԴ�ǻ�������Ĥ��,ͨ�������֯��������������,���ż�ǿ���ڼ���ϵ�����á��ҹ���ˮ��Ҫ��ֳ����,������(Carassiuscarassius)������(Ctenopharyngodonidella)����(Hypophthalmichthysmolitrix)...
��S3�����ڹ�Ӭ���������е�����̽��
�����嵰�ײ����ں�������װ�͵��ϳ�������Ҫ����,��ϸ�����ڡ�������DNA�������ȹ�����ͬ��������һ�����á�һЩ�����嵰���쳣���������༲���ķ���������ء�S3�Ǻ�����С�ǻ���һ����Ա,�к�������ø�Ļ���,�ǵ���ǰ�嵰��,������NF-B�������е�...
�ȵ������������ӵ��ڹ������������������û�
�ȵ������������Ӣ�IGF-����һ�ֽṹ���ȵ���ԭͬԴ�ĵ������ģ����зdz����ӵ�����ѧ���ܣ���ϸ���ķֻ�����ֳ����������������о�����Ҫ�ٽ����á��о�������IGF-��Թ���������������������Ҫ�������á�����ķ������ֱ�ӹ�ϵ����ֳ����ľ���...