��������ѧ����

����ժ Ҫ����ֲ���Ƥë��ֲ���Ƥϸ���γɵ�һ������ë״��,��ֲ������������������á�Ŀǰֲ���Ƥë�������Ͻ�(Arabidopsis thaliana)������(Lycopersicon esculentum)��ˮ��(Oryza sativa)���̲�(Nicotiana tabacum)������(Artemisia carvifolia)�ȶ��־���������ҩ��ֲ����չ���о������Ͻ��Ƥëϸ���ṹ��,Ŀǰ�����о���Ϊ�����ȫ��,Ϊ��������ֲ����о��ṩ��ָ������Ȼ����,��Ƥë��ֲ���ڲ����ⲿ�������һ����Ȼ����,��Ҫͨ�����������ͷ��ڻ�ѧ�������ֲ��Բ��������Ŀ��ԡ���ǰ���ڱ�Ƥë�о��Ѵ�ϸ�����淢չ�����ӻ��Ʋ��档���о���ֲ���Ƥë�����͡����ܡ����������ص㡢���ӵ����о�4����������ܽ�,Ϊ���ֲ���Ƥë���о��ṩ���ۻ���,ͬʱΪֲ������������ṩָ����
�����ؼ��ʣ���ֲ��; ��Ƥë; ��������; ���ӵ���;
����Abstract����Plant trichome is a special trichomes form by plant epidermal cells.It protected and regulated plants themselves.At present,the research on the trichomes of plants was widely distributed in various economic crops and medicinal plants such as Arabidopsis thaliana,Lycopersicon esculentum,Oryza sativa,Nicotiana tabacum,and Artemisia carvifolia etc.Trichomes of Arabidopsis had a simple structure.Therefore,its research was the most in-depth and comprehensive,and it also provided guidance for the subsequent research of other plants.In nature,trichome was a natural barrier between the interior of the plant and the external environment,which enhanced plant resistance to adverse environments mainly through physical defense and secretion of chemicals.The current research on trichome had evolved from the cellular level to the molecular mechanism.This review summarized the four aspects of the type,function,physiological and biochemical characteristics and molecular regulation of plant trichomes,and provided a theoretical basis for the study of plant trichomes in the future,and provided guidance for plant improvement and breeding.
����Keyword����plant; trichomes; develpment mechanism; molecular regulation;
����ֲ���Ƥë(trichome)����ֲ���Ƥϸ�������γɵ�һ�־�������ṹ��ë״������[1]����ͬ���͵ı�Ƥë��ֲ�����������������ӵ����ò�����ͬ,��ֲ������ͷ����вο���ֵ[2]����Ƥë�����ֲ��Բ��������������ֺ��ͻ�е���˵Ŀ��ԡ����Ƥë���Զ�����ֲ������ˮ�ֺ�������ɢ���봢��,Ӧ���¶����,�赲�������,����ֲ��ı������ʼ�������ֲ�����ϲ�����ͣ��ʱ��,ͬʱ��һ���̶�����ǿ��ֲ�������Ļָ������ȡ�����,�����ͱ�Ƥë�ɷ��ڳ�һЩ��ѧ��������ࡢ�����,���ֵ����P������в��,ͬʱ��Щ�����ﻹ��������ҩ�빤ҵԭ��,����һ���ľ��ü�ֵ[3,4]��ֲ���Ƥë��������ˮƽ���о�,���������ڽ���ϸ���ֻ������ۻ���,���Ҷ�ũҵ��������һ����ָ������[5]����ǰ��ģʽֲ�����Ͻ��Ƥë����ʼ���쳤����֧�ȹ��̵ķ��ӻ����Լ�ֲ�D���벻ͬ����Эͬ���ر�Ƥë�ķ�����ʽ���������̽����������о�[6],����ģʽֲ�������ǵ���Ҷֲ���Ƥë�������̵ķ��ӻ����Բ���ȷ�����Ķ�ֲ���Ƥë�����͡����ܡ����������ص�������ܽ�,��ģʽֲ�����Ͻ��Ƥë�������ӵ��ػ�������ģʽֲ���Ƥë�����о���̬���в�������,Ϊ���ֲ���Ƥë���о��ṩ���ۻ�����

����1 ����Ƥë������
����ֲ���Ƥë���ͷḻ,�ڲ�ͬ����,��ֲͬ�����Ϸ����Ų�ͬ�����á���Ƥëͨ���ɷ�Ϊ����ë״�������ë״��[7]��һ����ͷϸ������ϸ���ͻ�ϸ�����,��ëͷϸ���ͱ�ϸ����ϸ�����������з�ʽ��ͬ,�������а�״����״����Ƭ״����״����֦�Ͳ���֦��,����ë״��־��з��ڹ���,����ë״���ͷϸ�������з��ڹ���[8]��ֲ���Ƥë��Ϊ��ϸ��ë״��,ģʽֲ�����Ͻ��Ƥë�����翪ʼ�о��ĵ�ϸ����֧ë״��,ͬʱ����ֲ�O������״��һ�塣ˮ�����ڳ�ë��ë����ë3�ֱ�Ƥë,��ë�ֲ���ά�����Ĺ���ϸ����,ë����ë���ֲ����������˶�ϸ����[9]���ƹ�(Cucumis sativus)����6�ַ���ë,����4���Լ�Ķ��˽���,����2�ִ�����ͻ������,û�м�Ķ���,������һ���з�֧[10]������(Solanum)ֲ����ë��ͷϸ������4����������,��һ�־���1��2��ͷϸ��,�ڶ��־��ж��ͷϸ��,�����־��ж�С�ľ����Ľ�ϸ����ͷϸ��,���һ�־��м�С�Ķ�ϸ����ͷλ��һ����С�ĵ�ϸ������[11]����Ȼ����,��Լ90%��ëֲ��ë״�����һ�����ϸ��,ʣ��10%�����־��ж����֧��ë״��[12]����ֲͬ��������Ƥë�����Ͷ��ֶ���,��ֲ���������������з��ӵĹ���Ҳ������ͬ,����ԭ��,����ֲ������Ȼ�糤�ڵ��������������ʽ������ء�
����2�� ��Ƥë�Ĺ���
������Ƥëһ���̶���������ֲ���Ƥ�ĺ��,�Դ����ֵ�����������������ֺ�[13]����ë����Ҫ�����Dz������к������ý������õ����������ӷ����͡���֬�����༰����Ȼ�����,�������������������ж�������;��һ����,��ë�Ĵμ���л����Ҳ����һ���Ĺ�ҵ��ҩ�ü�ֵ���һ����Ե���ֲ�����������źŷ���[14]������ë��û�з��ڴ�л����,�����ڵֿ����˻������յ����ۡ���ֹ������䡢�ֿ��ɺ�����Ӧ���Ρ������ؽ����Լ�Ԥ�������ֺ��ͻ�е���˵ȷ��淢����ͬ����Ҫ�����á�����֮��,ֲ���Ƥë���ܶȡ���״�������������ؼ��������صIJ�ͬ,����ֲ��Ե�ǿ����ͬ[15]���̲ݵ���ë�����ų��̲ݱ��������еIJ�����,���ٶ�������˺�,ͬʱ��Щ��л�����������ɱ��������͡�ʳƷ���Ӽ���[16]���Ž�����[17]�о�����,��Ҷ����ë����ά��Ƥ�ĵ����������Ը�����Ҷ����ë����ά��Ƥ�ϵĺ���,�����ֲ���ũҵ���漰�����Ŀ��ԡ�������ư��(Coelophora inaequalis)���ھ��и��ܶȱ�Ƥë�ķ�����,�䷱ֳ�������˶�����ԶԶ�������ڵ��ܶȱ�Ƥë�ķ�����[18]�����Ͻ�ı�Ƥëϸ���ڴ��ϵ����䱡,���ܵ�������P�������ֺ�ʱ,������ǿ�������Կ�Ѹ����������[19]��������в�ȼ���,����(Nepeta cataria)��Ƥë�ĸ���������ҶƬ������ë�����ܶ�����,��������в�ȵ�Ӱ��[20]���Է���������(Saintpaulia ionantha)���е���в��,ͬ���������ޱ�Ƥë��ֲ����֯����������Ũ�ȸ����б�Ƥë��ֲ����֯[21]��
����3 ����Ƥë�����������ص�
������ֲ���ܵ����������������в��ʱ,ͨ���ı�Ҷ���ء���ԭ���ǡ��������ӵȵĺ����Լ�����ø�Ļ���,�糬������ø(CAT)�����������绯ø(SOD)����������ø(POD)����ά��������������ת[22]����Ƥë����һ���̶���Ӱ�����������ʵĺ�������ԡ����ٵ�[23]��ˮ���о��б�����ëˮ���Ĺ�����á��������á��������ʼ��������ڹ�ˮ���������ɵ�[16]��2�ֱ�Ƥë�����̲ݽ��е�������в��ʱ,�������̲�Ҷ������Ӻ������Զ��ڷǷ������̲�,��2���̲�����в������ë�ܶȾ������ӡ������Ͻ������ѧ����,��ϸ���ڸ�����Ũ�ȷ����仯,pH�������ؼ��������⺬������[24]���κ���[25]��4��Ʒ�ֵķ���ͬʱ���иɺ�в��,���Ѷ�ëƷ�ֵ���Ժ�ˮ����Ҷ���غ�����������ͨƷ��,Ҷ����ij��ṹ��ʧ�̶ȵ�����ͨƷ��;�ڿ����Բ�����,��ø�������������ȶ�ʱ,��ë����POD��SOD���Ը�����ͨ���ѡ�ֲ���Ƥë�ķ��ڹ�������ϸ���ڸ�ϸ�������������������ܲ��ɷ�[26]������ǿ��л���Եı�Ƥëϸ���д����Ŷ���ϸ����,�ȱ�֤��ֲ����������,����ǿ�˶Ի�е���˺Ͳ��������Ŀ���[27]��������ȫ���̲ݶ���ë���д���������,��ϸ����,��ϸ����,������״�������������,��Щ����ϸ������ǿ��л���Եĵ�������[28]��ͬʱ,��������Ҳ���������տ�(Helianthus annuus)����ĸ��(Leonurus artemisia)������(Mentha haplocalyx)��ֲ������ڵ���ëϸ����[29,30]�����̲ݡ����ѷ�������ë��ϸ���ʺ�Һ���д��ڴ�������ﰿ���������������,������Щ�������ʾ����µ���ʽ�ų�����[25,31]��ͨ��C14ͬλ���ٷ����̲ݳ�����ë�з���Ҷ��������˷�����ĺϳ�,���̱���ë��û��Ҷ����,�������ĺϳɳ����в���ȷ[32]��˯��(Withania kansuensis)��ë״��ϸ���е���������������ͷ������ĺϳɡ����ۺͷ��ڹ���[33]�����ο�(Labiatae)ֲ����ں���,ϸ������״�����ı�,��������������ճ��,˵��ϸ���˿���Ҳ�����˷�����ĺϳ�[34]����ë��ֲ��μ���л��������ϳɺ���й���ض�����,���ڱ�Ƥë����������ڼ�,��ϸ�����Ĺ��ܺ��Ƿ���������ĺϳ����ͷŵȷ��滹�������
����4�� ��Ƥë�ķ��ӵ����о�
������Ƥë����ʼ������������,����λ�ü���̬�ص����Ϊ�о�ϸ���ֻ������˾�������̬�������Ƶ�ģʽ���ϡ���ֲ�����ٷ���ʱ,��Ƥë�Ѿ���ʼ����,����������������ֲ����ֻ�����������ˮƽ�ʹ�������Эͬ�����������[6]��
����4.1�� ���Ͻ��Ƥë��ʼ�����ķ��ӵ��ػ���
������Ƥë�ķ���λ��,��С����״��������һϵ�л������,ͨ�����ڵı�Ƥϸ������ͬʱ������ë״�塣������Ϊ��ʼÿ��ϸ�������б�Ƥë�ٽ����Ӻ���������,�ٽ����ӱ�����ͬʱ����ϸ����Ƥë��������Ҳ������,�ٽ����Ӷ��ϸ�������γɱ�Ƥë,ͬʱ������������ȥ����ϸ��,���������Ƥë�ֻ�[35]��Ŀǰ�����Ͻ��Ƥë�����ķ��ӵ��ػ������2��ģ��,��һ��Ϊ��������ģ��:��Ƥë������ص�������MYB-BHLH-WD40����R2R3MYB����������TRY(TRIPTYCHON)��CPC(CAPRICE)��ʹ��������ϸ���ƶ�,����MYB��������ת¼������GL1(GLABRA 1)/MYB23-GL3(GLA-BRA 3),�����γ����Եĸ�����TRY/CPC-GL3/EGL3(ENHANCER OF GLABRA 3)-TTG1(TRANSPARENT TESTA GLABRA 1),��������ر�Ƥë���γ�[36]���ڶ���Ϊ��������ģ��:�Ա�Ƥë��������˫�����õ�WD40���嵰��TTG1��ϸ���������ƶ�,��BHLH�͵���GL3/EGL3��MYB�͵���GL1/MYB23����γ�GL1/MYB23-GL3/EGL3-TTG1�������Ա�Ƥë�������������,��ȱʧ��WD40�͵���TTG1�����ɷDZ�Ƥëϸ��[37](ͼ1)��
����ͼ1 ���Ͻ��Ƥë���ӵ��ػ���ģ��
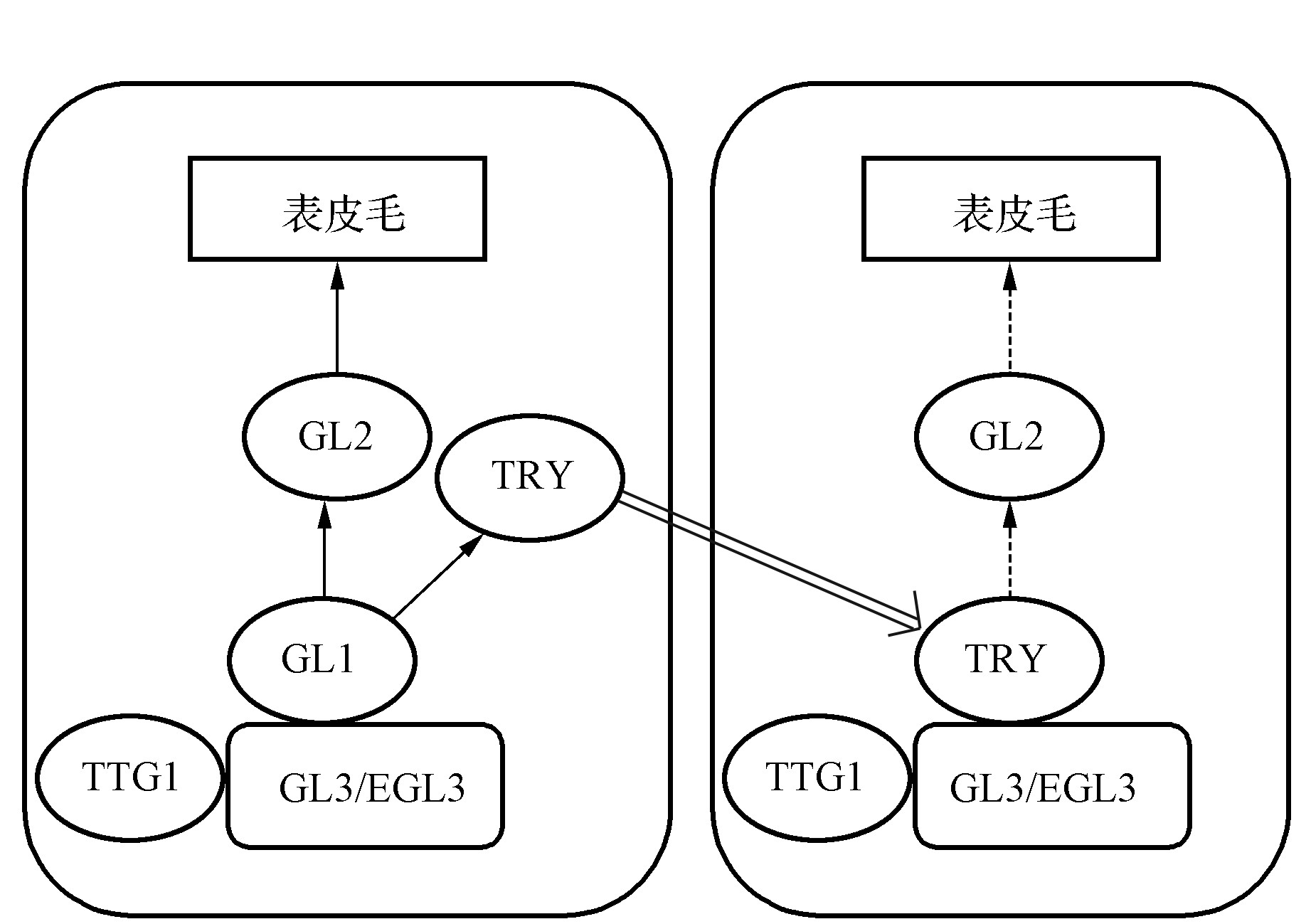
����Fig.1 Model of molecular regulation of trichomes in Arabidops
����ע::�������;:��������ϸ������˶�;:������ء�
����Note::Positive regulation;:Intercellular movement of protein;:Negative regulation.
����4.2 �����Ͻ��Ƥë�������̵ķ��ӵ���
����ģʽֲ�����Ͻ��Ƥë����������������Ҫ����3��ʱ��:1)��Ƥë��ʼ����,�����������������ӹ�ͬ���������ط��������е�һGL3����ͻ��,���Ͻ�ë״���������м���,�����������߶�ͬԴ����EGL3ͬʱ���÷���˫ͻ��ʱ,���ֹ�������,������ȫ��ëֲ�ꡣҰ�������Ͻ�GL2,CPC-TRY˫ͻ���干ͬ���õ�ֲ����,��Ƥë�ʹ������ֲ�,��CPC-TPY��˫ͻ����ֲ����,��Ƥë���������ҳʴ�״����[38]��2)��Ƥë����DNA���Ƽ���֧����,DNA����Խ��,ë״��Խ��,��֧����Խ��,��Ƥë�ķ�֧���ܵĵ���,�ܵ����з�������ű�Ƥë�������������֧����[5]����Ƥë����DNAҪ����4�θ��Ʋ����γɷ�֧�ṹ,�ڴ˹�����,GL3��TRY�ֱ��ǿ��ƺ���DNA���Ƶ�������������[39],SIM�������Ʊ�Ƥëϸ������˿����ת����ڸ���,�������Ʊ�Ƥë��֧���γ�,���Ͻ�SIMͻ������н��ٵķ�֧ë״��[40]������,��ù��(GA)�ĸ���������SPY(SPINDLY)�Ա�Ƥëϸ������DNA�ĸ���Ҳ��һ������������[41]��3)��Ƥë���쳤����,˿��ϸ���쳤�Ļ���,ϸ����������˿��ɢ,����״,�����ۼ���һ��,����ϸ�������쳤����,ֱ��ϸ������[42]��
����4.3 ��ֲ�D�ضԱ�Ƥë������Ӱ��
������Ƥë������ֲ�����ٷ���ʱ���Ѿ���ʼ,ֲ�D�ص�����ϳ�;�����źŴ���;�������˱�Ƥë�����ĸ�����[43]���Իƹ�ͬʱʹ��ϸ��������(CK)�ͳ�ù��(GA),������ë�Ĵ�С�ޱ仯,���ż���Ũ������,��Ƥë��������[11]����ͬŨ�ȵ����������(MeJA)��GA��������(IAA)�������Ѻ�,����Դ��ͬ���غ��������仯,����2����ë�ܶȴ������[44]����С��[45]����������в�ͬ���ش���,����JA��GA3���ܴٽ���άϸ����ʼ�ֻ�,���Ҷ��߾���Эͬ�ٽ����á���0.1 mmol·L-1��1.0 mmol·L-1��ˮ����(SA)�ֱ�Ӧ�������Ͻ��2��Ұ������ϵ,��Ƥë�ܶȾ�����,֤��SA�Ա�Ƥë�ķ���������������[46]�����Ӳ�����,�����Ͻ�Ҷ��ϸ���о�����,TEM(TEMPRANILLO)ת¼���Ӽ���ϸ����GA�Ļ��ۺ�ת�˵��ı����TEM�ĵ�����,Ҷ��ϸ��ѡ���Ե����ܱ�ϸ������GA,����Ӱ���Ƥϸ���ķ���ģʽ[37]��JA�ɵ�������ά�ķ������쳤,GhJAZ2��������GhMYB25-like��GhMYC2��ת¼��������ò����������,��������������ά����[47]����ɫ���ᵥ����ø����(IAAM)�����Ͻ��Ƥë�������GL2���������������ת�����Ͻ漰�̲���,���Ͻ����̲ݵı�Ƥë�ܶȾ���������,֤����GL2����ˮƽ��IAA��Ӱ��[48]�������о�˵��,��Ƥë�ķ�����ʽ������������ֲ�D���벻ͬ�����Լ���ͬ���ؼ��Эͬ����,���дִٽ�ֲ�������༤�ص���Դ������һ��Ũ�ȷ�Χ�������˱�Ƥë���ܶȼ�����(ͼ2)��
����ͼ2 ֲ�D�ضԱ�Ƥë�γɵķ��ӵ���ģʽ
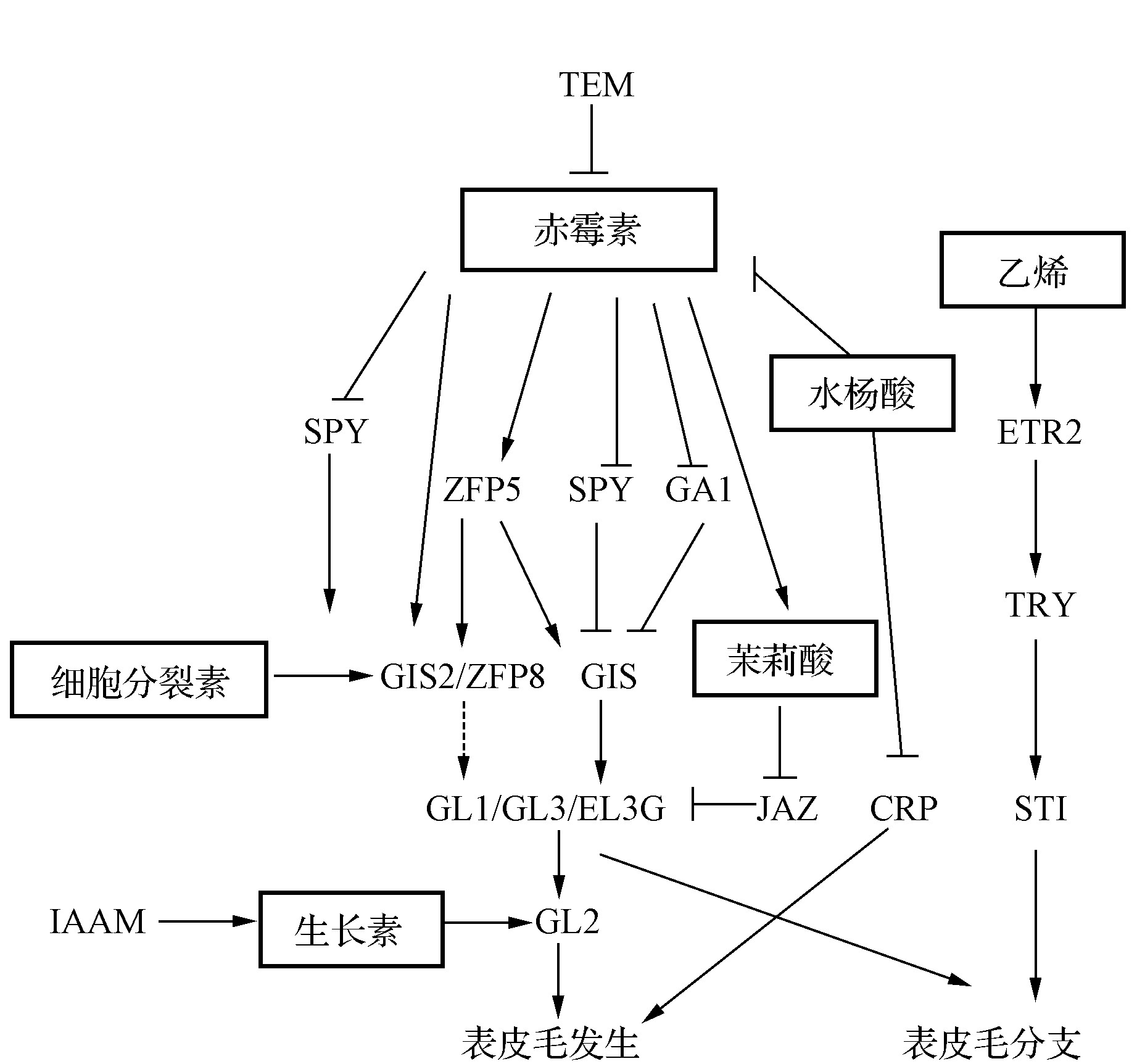
����Fig.2 Molecular regulation patterns of plant hormones on trichome formation
����ע::�ϵ���ٽ�;:�µ�������;:���Ʒ�����
����Note::Up or promote;:Down or inhibit;:Inhibition occurs.
����4.4 ����ģʽֲ���Ƥë�����о���չ
������ǰ�Ա�Ƥë�ķֻ����̼����ӵ��ػ����о��ཨ�������Ͻ浥ϸ����Ƥë�о�������,�������ɶ�ϸ�����ɵ�ֲ���Ƥë���ߵ��γɻ���������֮ͬ��,�����߸�Ϊ���ӡ����ز�ͬ���ͱ�Ƥë�����Ĺؼ�����ͬ,ͬһ�����ڲ�ֲͬ���Ƥë���������е�����Ҳ���ڲ���[49]���о�����,�������(Antirrhinum majus)��R2R3����ת¼���ӵĵ��ػ���MIXTA��MIXTA LIKE1ת���̲���,�̲�ҶƬ�;��ϵı�Ƥë�����������,���������Ͻ�ͻ�����й����ﲢû���������Ʊ仯[50]�����������CotMYB��MIXTA����һ��ͬԴ��,�������Ͻ��й����ﲻ�ı����Ͻ�ı���,�������̲ݱ�Ƥë����,����TRY�����̲�ҶƬ��Ƥë�ķ����������Ͻ���Ӱ��[51]�����Ͻ��Ƥë��������CPC�Է��ѱ�Ƥë��״��Ӱ��,������TRY��ͬԴ����SLTRY,�ٽ����ѵĵ��ϱ�Ƥë����,ת�����Ͻ��,ȴ�������Ͻ���ϱ�Ƥë�ķֻ�,�ٽ����¸�ë����[52]��
����Ŀǰ,�Է�ģʽֲ���Ƥë�����ķ��ӻ��Ƶ��о���������,����ˮ��(Oryza sativa)���ƹ�(Cucumis sativus)������(Lycopersicon esculentum)����(Gossypium hirsutum)���ƻ���(Artemisia annua)�Ⱦ��������ҩ��ֲ���ѿ�չ��ط��ӻ����о�����WO(WOLLY)����Ӱ�췬�ѱ�Ƥë�ķ���,SIMYC1��SIEOT1������Ʒ��ѱ�Ƥë�������ʵĺϳ�[53]���ƹ��е�CSGL1��������ҶƬ��������ʵ��ë״����γ�,���Ը�ë���γ�������Ӱ��[54]��Mhl1����ʹ���ױ�Ƥ�γɾ�ë,���䷢��ͻ���ֱ�ӵ��������о�ë���γ�[55]��HL6��OsWOX3B����Эͬ����ˮ����Ƥë���γ����쳤,HL1(HAIRY LEAF-1)��HL2(HAIRY LEAF-2)����Ӱ���仨����֧�ı�Ƥë����,HG(HAIRY GLUME)����Ӱ��ҶƬ��ԵС��֦�ı�Ƥë����[51]���ƻ�����ë������ϳ��ܵ�ADH2��������һ�ִ�����ø����,Ӱ�쵥��ͪ������ϳ�[55]������ǰ����Щ����ķ��ӻ������˽����١�
����5�� ������չ��
����ֲ���Ƥë��Ϊֲ�������Ӵ��ĵ�һ������,��״����,�Ҿ��ж��ֹ���,��ֲ������������������������Ҫ�ĵ�������,��Ƥë�����ֲ�������Ŀ���,ʹ���ڶԿ��ɺ����亦���溦�������ȷ������������ޱ�Ƥë��ֲ��,һ���̶��ϼ�����ũҩʹ�öԻ�����ɵ���Ⱦ,��Լ��ˮ��Դ��ͬʱ��Ƥë���������,�ؽ�����,��е���˵���ǿ��Ӧ����������,Ϊ��������,ɳ���̻��ṩ�˱�ѡֲ����з��ڹ��ܵ���ë���������������������,��л����Ҳ����Ϊ�����Ļ�ѧ���Ϻ�ҩ�ò��ϡ����Ӳ�����,����֤�ݱ���˫��Ҷֲ��������ƵĿ��Ʊ�Ƥë�γɻ���,����ϸ�����ϸ��ֲ���Ƥë�������ƴ��ڲ���,�����ڵ���Ҷֲ����,��Ƥë�ķ������û����Բ��Ǻ�������Զ���ֲ��ı�Ƥë�о�Ҳ����,ͬһ�������ڲ�ͬ��ֲ���еı��﷽ʽ������ͬ,ֲ�����յı���Ҳ������ͬ����ǰ��ֲ���Ƥë�ķ��ӵ����о��༯����ģʽֲ�����Ͻ���,��ģʽֲ��ı�Ƥë���ط�ʽ������������Ķ�ֲ���Ƥë�����͡����ܡ����������ص㼰���ӵ��ػ��ƽ������ܽ����,����Ϊ���ֲ���Ƥë������о������ֹ����ṩ�ο����ݡ�
���������
����[1]BALKUNDE R,PESCH M,HLSKAMP M.Trichome patterning in Arabidopsis thaliana from genetic to molecular models[J].Current Topics in Developmental Biology,2010,91(91):299-321��
����[2]ATTALA Z,CELEP F,BARA F,et al.Systematic significance of anatomy and trichome morphology in Lamium (Lamioideae;Lamiaceae)[J].Flora,2016,225:60-75��
����[3]YAN A,PAN J B,AN L Z,et al.The responses of trichome mutants to enhanced ultraviolet-B radiation in Arabidopsis thaliana[J].Journal of Photochemistry and Photobiology B:Biology,2012,113(1):29-35��
����[4]ANDRADE M C,SILVA A A D,NEIVA I P,et al.Inheritance of type IV glandular trichome density and its association with whitefly resistance from Solanum galapagense accession LA1401[J].Euphytica,2017,213(2):52��
����[5]SCHULMAN S,H�aLSKAMP M.Epidermal differentiation:Trichomes in Arabidopsis as a model system[J].International Journal of Developmental Biology,2005,49(5-6):579-584��
����[6]KHOSLA A,PAPER J M,BOEHLER A P,et al.HD-Zip proteins GL2 and HDG11 have redundant functions in Arabidopsis trichomes,and GL2 activates a positive feedback loop via MYB23[J].Plant Cell,2014,26(5):2184-2200��
����[7]�����ã�����飬Фǿ���ȣ�ľܽ�ز�ͬʱ�ڻ��Ľṹ�仯�о�[J]�������գ�2016(7):50-54��
����[8]����ƽ���ܶ��࣬ţ��־���ȣ���ͬ�̲�������ë�ܶȼ�������[J]���й��̲ݿ�ѧ��2018,39(4):7-17��
����[9]WANG Y P,CHEN W L,QIN P,et al.Characterization and fine mapping of GLABROUS RICE 2in rice[J].Journal of Genetics&Genomics,2013,40(11):579-582��
����[10]XUE S D,DONG M M,LIU X W,et al.Classification of fruit trichomes in cucumber and effects of plant hormones on type II fruit trichome development[J].Planta,2019,249(2),407-416��
����[11]KANG J H,FENG S,JONES A D,et al.Distortion of trichome morphology by the hairless mutation of tomato affects leaf surface chemistry[J].Journal of Experimental Botany,2010,61(4):1053-1064��
����[12]NOGUEIRA A,OTTRA J H,GUIMARAES E,et al.Trichome structure and evolution in Neotropical lianas[J].Annals of Botany,2013,112:1331-1350��
����[13]�ع�ԣ���骣��Ƶ��ǣ��ຣ���ϲ���Ҫ��������Դ�ֲ���������������ֲ��[J]���ຣ��ѧѧ����2017,35(4):50-55��
����[14]RIDDICK E W,SIMMONS A M.Do plant trichomes cause more harm than good to predatory insects[J].Pest Management Science,2015,70(11):1655-1665��
����[15]RIDDICK E W,SIMMONS A M.Plant trichomes have mixed impacts on predatory insects[J].Pest Management Sciene,2015,70(11):1668��
����[16]����ɣ������㣬�����ᣬ�ȣ������ͺͷǷ������̲�Ҷ����ë����в�ȵ�Ӧ��Ӧ�о�[J]���й�ũҵ�Ƽ�������2019,21(6):55-60��
����[17]�Ž��������壬���Ʒ����ȣ�����ͬ��ë��״���Ȼ���ϵ��ѡ�����������������о�[J]������ũѧ����2017,32(3):137-142��
����[18]HEINZ K M,ZALOM F G.Performance of the predator Delphastus pusillus on Bemisiaresistant and susceptible tomato lines[J].Entomologia Experimentalis Et Applicata,2012,81(3):345-352��
����[19]LIU H,ZHOU L H,JIAO J,et al.Gradient mechanical properties facilitate Arabidopsis trichome as mechanosensor[J].Acs Applied Materials&Interfaces,2016,8(15):9755-9761��
����[20]ZHOU Y,TANG N Y,HUANG L J,et al.Effects of salt stress on plant growth,antioxidant capacity,glandular trichome density,and volatile exudates of Schizonepeta tenuifolia briq[J].International Journal of Molecular Sciences,2019,19(1):252��
����[21]SALTVEIT M.E,HEPLER P K.Effect of heat shock on the chilling sensitivity of trichomes and petioles of African violet(Saintpaulia ionantha)[J].Physiologia Plantarum,2010,121(1):35-43��
����[22]�����ԣ�����ƽ����־¼���ȣ���ͬƷ�ֹ�������Ե��µ�������Ӧ���ͺ�������[J]������ũҵ��ѧѧ����2018,39(2):40-46��
����[23]���٣��。������ǣ��ȣ�ˮ����Ҷ��״����Ƥë�����о���չ[J]����ũѧ����2015,29(11):2110-2116��
����[24]����ǿ����־�ޣ�����ޣ��ȣ�ֲ���������̽������Ƥë����ѧ�̼���Ƥë�շ��ķ�����Ӧ[C]���Ͼ����й�ֲ�ﲡ��ѧ��2016��ѧ����ᣬ2016��
����[25]�κ��ۣ����ѱ�Ƥë���������ڿ�в���еĹ����о�[D]��������������ũҵ��ѧ��2017��
����[26]STRATMANN J W,BEQUETTE C J.Hairless but no longer clueless:Understanding glandular trichome development[J].Journal of Experimental Botany,2016,67(18):5285-5287��
����[27]���������������Բ���ҶƬ���ϵͳ����������л�����ṹ��Ӱ��[D]���Ϸʣ�����ũҵ��ѧ��2015��
����[28]UZELAC B,JANOEVIC'D,STOJICIC'D,et al.Morphogenesis and developmental ultrastructure of Nicotiana tabacum short glandular trichomes[J].Microscopy Research&Technique,2017,80(7):779-786��
����[29]TURNER G W,GERSHENZON J,CROTEAU R B.Development of peltate glandular trichomes of peppermint[J].Plant Physiology,2000,124(2):665-679��
����[30]AMEREN E,HELLER A,SPRING O.Capitate glandular trichomes of Helianthus annuus(Asteraceae):Ultrastructure and cytological development[J].Protoplasma,2014,251(1):161-167��
����[31]�Ż����̲���ë��̬���ṹ������������о�[D]��֣�ݣ�����ũҵ��ѧ��2008��
����[32]KEENE C K,WAGNER G J.Direct demonstration of duvatrienediol biosynthesis in glandular heads of tobacco trichomes[J].Plant Physiology,1985,79(4):1026-1032��
����[33]MUNIEN P,NAIDOO Y,NAIDOO G,et al.Micromorphology,histochemistry and ultrastructure of the foliar trichomes of Withania somnifera (L.)Dunal(Solanaceae)[J].Planta,2015,242(5):1107-1122��
����[34]�����ܣ����࣮���ο�ֲ����ë��������ë���ڹ��ܵ��о���չ[J]�������գ�2012(10):194-196��
����[35]MOOSE S P,NICK L,CARLSON S R.The maize macrohairless1locus specifically promotes leaf blade macrohair initiation and responds to factors regulating leaf identity[J].Genetics,2004,166(3):1451��
����[36]YANG C X,YE Z B.Trichomes as models for studying plant cell differentiation[J].Cellular&Molecular Life Sciences,2012,70(11):1937-1948��
����[37]REINHARDT H,GIERER A.Applications of a theory of biological pattern formation based on lateral inhibition[J].Journal of Cell Science,1974,15(2):321-346��
����[38]ZHANG F,GONZALEZ A M,PAYNE C T,et al.A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis[J].Development,2003,130:4859-4869��
����[39]����ϼ����ǫ����꣬�ȣ�ֲ���Ƥë�������ӵ����о���չ[J]��ֲ������ѧ����2019(6):711-717��
����[40]GREBE M.The patterning of epidermal hairs in Arabidopsis--updated[J].Current Opinion in Plant Biology,2012,15(1):31-37��
����[41]LI J A,ZHONG J Z,SHA S,et al.GLABROUS INFLO-RESCENCE STEMS (GIS)is required for trichome branching through gibberellic acid signaling in Arabidopsis[J].Plant&Cell Physiology,2012,53(2):457-469��
����[42]SUO B X,SEIFERT S,KIRIK V.Arabidopsis GLASSYHAIR genes promote trichome development[J].Journal of Experimental Botany,2013,64(16):4981-4991��
����[43]PATTANAIK S,PATRA B,SINGH S K,et al.An overview of the gene regulatory network controlling trichome development in the model plant,Arabidopsis[J].Frontiers in Plant Science,2014,5(5):259��
����[44]������¿��������飬�ȣ���ʩGA��Me��JA��IAA��SA��KT�Է��ѱ�Ƥë����������[J]����ѧ����2016,43(11):2151-2160��
����[45] ��С�ԣ�����Gossypium hirsutum)GhJAZ3��GhGAI1��JA��GA�鵼��ת�������Ͻ��Ƥë�ֻ����쳤�еĹ����о�[D]���人������ʦ����ѧ��2016��
����[46]TRAN M B,BERGELSON N.Interactive effects of jasmonic acid,salicylic acid,and gibberellin on induction of in Arabidopsis[J].Plant Physiology,2003,133(3):1367-1375��
����[47]HU H Y,HE X,TU L L,et al.GhJAZ2 negatively regulates cotton fiber initiation by interacting with the R2R3-MYBtranscription factor GhMYB25-like[J].Plant Journal,2016,88(6):921-935��
����[48]л���ۣ��˼��������࣬�ȣ��̲ݺ����Ͻ��Ƥë���������iaaM����Ŵ�ЧӦ[J]������ũҵ��ѧѧ����2018,246(3):59-64��
����[49]ZHANG X L,YAN F,TANG Y W,et al.Auxin response gene SlARF3 plays multiple roles in tomato development and is involved in the formation of epidermal cells and trichomes[J].Plant&Cell Physiology,2015,56(11):2110��
����[50]MARIA P R,JAFFE F W,EUGENIO B,et al.Development of three different cell types is associated with the activity of a specific MYB transcription factor in the ventral petal of Antirrhinum majus flowers[J].Development,2005,132(2):359-370��
����[51]WU X,LI Y E,SHANGGUAN X X.Progress in studies on transcription factors related to cotton fiber development[J].Cotton Science,2013,25(3):269-277��
����[52]RUMI T W,YUKA N,SHUSEI S,et al.Control of plant trichome and root-hair development by a tomato(Solanum lycopersicum)R3 MYB transcription factor[J].PloS One,2013,8(1):e54019��
����[53]SPYROPOULOU E A,HARING M A,SCHUURINK RC.RNA sequencing on Solanum lycopersicumtrichomes identifies transcription factors that activate terpene synthase promoters[J].BMC Genomics,2014,15(1):402��
����[54]LI Q,CAO C J,ZHANG C J,et al.The identification of Cucumis sativus Glabrous 1 (CsGL1)required for the formation of trichomes uncovers a novel function for the homeodomain-leucine zipper 1gene[J].Journal of Experimental Botany,2015,66(9):2515-2526��
����[55]POLICHUK D R,ZHANG Y,REED D W,et al.A glandular trichome-specific monoterpene alcohol dehydrogenase from Artemisia annua[J].Phytochemistry,2010,71(11-12):1264-1269��

miRNA��ֲ�↑��ʱ�估�����ٷ���������
����ֲ������Ҫ�ľ������٣����ڡ����ͼ�����ķ�֦����Ҫ��״���ǻ����������ڱ���.���������о�Ҳ�Ƿ�������ѧ�ͷ�������ѧ�о������Ծ������ֲ������ѧ���о�����������ֲ�ᆳ��һ����Ӫ�������Σ�����Ӫ���ͷ�����֯�������̬.��ʱ�������˵����...

ֲ��������UV-B���յ����ü����������
̫�����в���Ϊ100��400 nm�ĵ�ŷ����Ϊ������(UV)���������ײ������Խ����Ϊ����:����������(UV-A)��������Χ400��320 nm;�в�������(UV-B)��������Χ320��280 nm;�̲�������(UV-C)��������Χ280��180 nm��...
���ص���ֲ�������������о��ɹ�����
��������һ��ֲ�D����Ҫ���������ᡢ������������������������ᡢ12-��-ֲ���ϩ��Ȼ���ͪ�����������1962�꣬�����������Ϊһ�����ϴ���ܰ���з��������������70������о�����������ʶ�������ص�����ѧ���ܡ��ϳ�;�������źŴ���;����...
����ֲ����̬�ṹ����ӵ�����֮һ��Ҳʹ�ñ���ֲ���ֲ�����������½��ֲ�����Բ�ͬ��������Alvarez-Buylla��2010����ˮ����ũ���ﻨ���ٵķ�����Ϊ�����ṩ����Ҫ����ʳ��Դ��1��ѧ���ȵ��о�����֮һ��ˮ����ԭ����С�������֯����������С��...

Ŀǰ�����ڴ������о����֣����»��۰�����Ҫ��������ĺ��������쳣��Ĥ֬�����������ʴ�л���Һ�СRNA�����쳣���������еĹ�����...

�ʺ�����ֲ�����о��ж����������ܣ���Ҫͨ����ֲ�����ø���Ե����ߣ����ͻ������������ﺬ����������������ɻ����������������ˣ����ֲ��Ŀ����ԡ�...
��������һ�������о����ص��ػ���������ƽ̨���������Ļ�������Դ��ԭ��Ƥ�ض�ϸ��ѡ�����������;����Ϊ�������֯ĸϸ����mothercellofmeristemoid,MCM��������ϸ���������ȷ��ѣ�����С���������֯ϸ����meristemoidcell,M...
���ާ��ֲ���������ڸ������ε���̬�ṹ��
���ާ��(Athyriaceae)Ϊ��С�͡�����Ϊ��������ާ��ֲ�Լ��20��500�֣��㲼ȫ�����ȴ������´����أ������ȴ������ȴ�ɽ��Ϊ�ࡣ�ҹ�����������Լ400�֡�ާ��ֲ�������ȷ��������巢�������Կ���Ϊ����ѧ���ֵ��о��ṩ��Ч�����ݡ�������������...
���DZ���ֲ�����е��ڷ�ֳ�����в�������ֳ���٣���ֲ����֯����Ϊ���ӵĽṹ��Ҳ�DZ���ֲ������������½��ֲ�����Ҫ�����������Ǹߵ�ֲ����巢�������Ļ��ڣ�Ҳ��ֲ���Ӫ������ת����ֳ���������ܺ���Ĺؼ����衣��˫��Ҷֲ�����Ͻ�Ϊ������Ӫ������ʱ...
���Ը�ϵ��ֲ�ﳤ����Ӧ½���������γɵ���Ҫ���٣���ֲ���������������з����Ź̶�ֲ�ꡢ����ˮ�ʡ��ϳ������л������ʵ��ڶ��[1-4],��ӵص����ŵ��ϲ������Ҫ�����羥��Ҷ��������ʵ��������������[5-6].��ˣ��о�ֲ���ϵ���������ĵ��ػ�������...