昆虫学论文


花绒寄甲( Dastarcus helophoroides Fairmaire) 属鞘翅目( Coleoptera) 穴甲科( Bothrideridae) 昆虫。最早称该虫为花绒坚甲[1],后来又将其改名为花绒穴甲,归入穴甲科[2],最终又依据该虫的寄生性生物学特性将其定名为花绒寄甲[3]。花绒寄甲成虫的食性较杂,取食枯朽的树皮、木块以及天牛幼虫和其它昆虫的干尸体[4]; 幼虫为闯蚴型,孵化后行动十分活跃,能依靠其发达的胸足快速爬入天牛蛀蚀的坑道寻找寄主。花绒寄甲是多种害虫的寄生性天敌,其主要寄主不仅有严重危害经济与生态林木的光肩星天牛( Anoplophora glabripennis Motschulsky) 、桑天牛( Apriona germari Hope) 、云斑白条天牛( Batoceralineolata Chevrolat) 、松褐天牛( Monochamus alternatusHope) 等[5 -6],还有与天牛类害虫同宿主的吉丁虫科的六星吉丁( Chrysobothris succudanea) 、十斑吉丁( Melanophila decastigma Fabricius) 等。因此,利用花绒寄甲建立天牛类害虫治理技术体系,无论对于天牛类害虫的有效防治,还是避免过度使用农药给生态环境带来的危害,都具有十分重要的意义。
已有研究者对花绒寄甲的生物学特性[5]、分类地位[2]、分布和寄生范围[6 -7]、行为学特征[2,8 -9]、与天牛互作的化学生态学[10 -14]、耐寒性[15]、人工繁育方法[16 -20]、林间释放防治天牛的方法[21 -24]等进行了较为系统的研究,特别是近年来花绒寄甲在天牛类害虫的生物防治上广泛应用并取得了重大成果。
本文在对所有关于花绒寄甲的研究文献信息进行分析的基础上,重点对花绒寄甲的分布情况、感觉系统和行为学研究及其在天牛科害虫生物防治应用等方面的最新研究进展进行综述,以期为建立我国林业蛀干害虫综合治理策略提供参考,为今后深入开展植物-天牛-花绒寄甲三级营养系统的互作机制研究提供科学借鉴。
1 花绒寄甲的研究文献分析
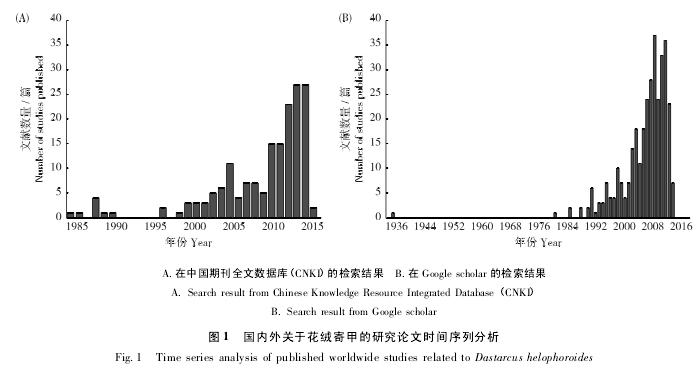
通过在中国期刊全文数据库( CNKI) 搜索“花绒寄甲”或“花绒坚甲”、“花绒穴甲”关键词的文献,共检索到相关文献 147 篇。147 篇文献的时间序列分析表明我国对花绒寄甲的研究在 2000 年以前呈零星分布状态,2000 年以后的研究呈现上升趋势( 图 1-A) 。20 世纪 80 年代以后,蛀干害虫对我国林业生产的危害日趋严重,害虫的生物防治也逐渐受到重视,这也许是 2000 年之后有关花绒寄甲的研究论文逐渐增多的原因之一。
在 Google scholar 搜索“Dastarcus helophoroides”和“Dastarcus longulus”关键词,共查到相关文献 337篇。对文献的时间序列分析表明,国际上关于花绒寄甲的研究始于 20 世纪 80 年代,90 年代起因光肩星天牛传入北美和欧洲成为重要的检疫性害虫,花绒寄甲相关的研究论文数量呈现显着上升趋势( 图1-B) 。
以 CNKI 中 147 篇花绒寄甲相关文献关键词出现频次进行分析: 以松褐天牛检索的文献出现 33次,以生物防治检索的文献出现 26 次,以光肩星天牛检索的文献出现 11 次( 表 1) 。由此可以看出,花绒寄甲对松褐天牛、光肩星天牛等蛀干害虫具有较好的防治效果。因此,进一步对花绒寄甲进行细致深入的研究,可以为林业柱干害虫生物防控技术提供理论基础和技术支持。

2 花绒寄甲的地理分布
早期的研究认为花绒寄甲分布于东经 105° ~135°、北纬 30° ~ 40°的地域范围内,为我国和日本所特有,但2012 年有报道称在韩国也发现了该虫的踪迹。花绒寄甲在日本分布于本州( 冈山和广岛) 、九州[25]和大阪[2],主要寄主为松褐天牛。分布在韩国的花绒寄甲其主要寄主为松褐天牛和云杉花墨天牛( Monochamus saltuarius Gebler)[26]。从动物区系的观点来看,花绒寄甲的分布范围跨越古北区与东洋区两大区系,具有较为广泛的生态适应性[7]。
花绒寄甲在我国的分布范围广泛,南至广东省深圳( 北纬22°38') ,北到吉林省梅河口( 北纬 41°41',东经125°54) 均有该虫分布[6],主要分布的省( 区) 有广东、江苏、安徽、河北、河南、山西、山东、宁夏、陕西、北京、吉林、辽宁等。该虫在我国的越冬北界为从吉林省梅河口到辽宁省凤城,经北京、山海关到陕西省延安,再北上至宁夏回族自治区银川。之后魏建荣等[7]的调查发现,该虫广泛分布于北纬22°26' ~42°32',在我国西北半干旱地区的行道树或防护林( 杨树) ,南至广州的亚热带马尾松林,北至吉林省( 中温带) 的天然蒙古栎、辽东栎的阔叶次生林中都有该虫的踪迹,其分布范围进一步扩大。此外,适合花绒寄甲生长的海拔高度为3. 3 ~1 184. 9 m[6]。
基于 GIS 平台的农林作物病虫害监测预警管理系统可以满足时空数据处理的要求,有效分析森林病虫害发生和分布的时空性[27 -29]。该平台不仅可以预测病虫害发生和分布,还可以全面展示害虫天敌的分布区、扩散范围等。基于 GIS 平台可进一步研究花绒寄甲的分布格局、扩散和适生范围等。
3 花绒寄甲的感觉系统及行为学研究
3. 1 花绒寄甲的感觉系统
昆虫的感受器分为嗅觉感受器、视觉感受器、触觉感受器、听觉感受器等[30],接受外界信号的最主要器官是触角和复眼,对觅食、求偶、定位等具有重要作用[31 -35]。对昆虫复眼和触角外部及内部显微与超微结构的研究,是了解其行为学的基础。花绒寄甲寄主( 天牛幼虫等) 的生活场所较隐蔽,花绒寄甲成虫在产卵场所的选择上就需要有精确的定位和识别[36],因此推测花绒寄甲拥有独特的复眼和触角。
目前对花绒寄甲感觉器官的研究还相对较少。Ren 等[37]利用扫描电子显微镜和透射电子显微镜对花绒寄甲成虫触角进行了详细研究: 成虫的触角呈锤状( capitate) ,长约937 μm ±59. 6 μm,由柄节、梗节及鞭节( 9 节) 组成; 雌雄成虫的触角没有明显差异。但是该项研究中并没有对雌雄成虫触角的长度、直径等进行差异显着性分析,这也许是没找出差异的原因之一,不过也有可能不存在雌雄二型现象,因此目前尚未将触角作为花绒寄甲雌雄性别辨别的依据[38]。花绒寄甲具有弱趋光性[8],但是对该虫视觉系统及视觉蛋白受体等的相关研究尚未见报道。
3. 2 花绒寄甲的行为学研究
由于对花绒寄甲的感觉器官研究报道较少,因而对该虫的行为学研究也相对薄弱。观察花绒寄甲成虫的行为发现: 在弱红光下,该虫的视觉系统对寄主定位的影响较小,但还不清楚其他单色光刺激是否会对该虫的行为产生影响; 成虫的飞行活动能力较强,交尾成一字形等[8]。另有研究者对植物-天牛-天敌的三级营养系统进行了初步研究。调查分布于陕西、吉林、广东 3 省的花绒寄甲成虫对不同天牛虫粪选择性和对天牛的寄生率,结果表明所有的花绒寄甲种群均选择各自本源的寄主,对不同寄主的寄生率也有差异[12]。利用光肩星天牛幼虫的虫粪对花绒寄甲进行嗅觉信号刺激,发现花绒寄甲对取食不同植物光肩星天牛幼虫虫粪的反应具有显着差异[11]; 使用锈色粒肩天牛( Apriona swainso-ni) 等天牛幼虫虫粪对花绒寄甲进行刺激时,也发现对花绒寄甲成虫具有吸引作用[13 -14]。鉴定锈色粒肩天牛[取食槐树( Sophora japonica) ]和光肩星天牛[取食垂柳( Salix babylonica) ]幼虫虫粪挥发物的主要成分为单倍烯,利用 Y 型嗅觉仪测试花绒寄甲对单倍烯的反应,其中对 α-蒎烯和 β-蒎烯的反应较灵敏[10]。
综上所述,对花绒寄甲的感觉系统及行为学的研究,主要集中在嗅觉及其化学生态学方面。然而,昆虫对寄主、配偶、产卵场所的定位和识别行为不仅仅只是依靠嗅觉作用,视觉、味觉、触觉等感受系统也是导致其行为活动的重要因素[39 -41]。因此,深入全面地开展该虫感觉系统的研究,有利于阐释该虫的行为活动规律,从而为更好地应用该虫构建林业蛀干害虫的生物防治技术体系提供科学借鉴。
4 花绒寄甲在天牛科害虫生物防治的应用
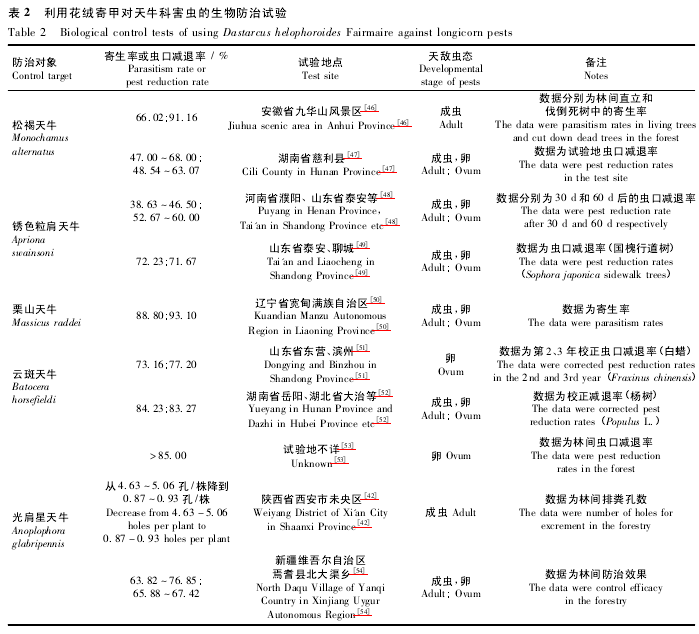
我国的林业长期遭受天牛类昆虫的危害,已造成重大的经济损失和较严重的生态损失[42]。例如: 光肩星天牛等曾给我国的“三北防护林”造成了毁灭性的危害[43 -45]; 松褐天牛还是传播松材线虫病的主要媒介,造成病虫害对林业的协同危害。花绒寄甲作为防治天牛类蛀干害虫最有效的天敌,对松褐天牛、光肩星天牛、云斑天牛( Batocera horsefieldi) 、栗山天牛( Massicusraddei) 等大中型天牛均有较好的寄生效果[5]。大量试验数据( 表2) 表明: 花绒寄甲对天牛类蛀干害虫的寄生率在66%以上; 用于防治锈色粒肩天牛的虫口减退率在38. 63% ~60. 00%,防治松褐天牛的虫口减退率在 47. 00% ~68. 00%,防治其它天牛的虫口减退率也均在 72% 以上。虽然释放花绒寄甲成虫对天牛类害虫的防治效果较好,但是花绒寄甲成虫的饲养成本较高。根据长时间的林间试验,采用花绒寄甲成虫和卵对天牛进行生物防治的效果并无显着差异[55],因此从降低防治成本考虑,当前仍是通过在林间释放花绒寄甲卵卡对天牛类害虫进行生物防治。花绒寄甲在天牛类害虫生物防治中的应用,为我国的经济与生态林木生产做出了重大贡献。
以植食性昆虫为中心的三级营养关系及其相互作用一直是昆虫学研究的热点之一[56],也是合理调控害虫及其天敌种群结构,分析昆虫与植物协同进化关系的重要理论基础。在明确植物对害虫的抗性强弱、植物如何吸引天敌等基础上,可以合理搭配植物结构提高整个生态系统的稳定性,从而更好地发挥天敌对害虫的生物防治效果。但是,目前对于寄主植物-天牛-花绒寄甲三级营养系统的研究还不够完善,现有的研究主要是针对天牛-寄主植物二者之间的相互作用机制[57 -59]以及天敌对天牛的生物防治效果[60]等,今后需要就寄主植物-天牛-花绒寄甲的三级营养系统及互作机制开展深入系统的研究。
5 展望
由于全球性气候变暖,甲虫类害虫的危害将越来越严重,尤其是天牛类害虫常常给林业生态系统造成毁灭性危害[5]。光肩星天牛在我国以及北美、欧洲国家的部分林区的危害严重,据不完全统计,到 2008 年害虫已分别给北美和欧洲造成约 40 亿美元、60 万欧元的经济损失[45]。其他天牛科害虫如桑天牛不仅危害桑树,而且取食苹果树等经济林木[61]; 松褐天牛是检疫性病害松材线虫病的重要传播媒介[62]; 云斑天牛和栗山天牛等大中型天牛均是林业生产的危险性害虫[63]。由于天牛蛀干危害,生活场所较隐蔽,对其进行化学防治不仅效果不好,而且对其它天敌昆虫、花粉媒介昆虫的伤害大[64],同时还存在农药污染及防治成本过高的问题。因此,利用有效的天敌昆虫对天牛类害虫进行生物防治,不仅能够减少化学农药对环境的污染,而且对于维持生态系统的稳定性和生物的多样性具有重要意义。
植食性昆虫及其天敌对寄主、配偶、产卵场所等的定位和识别是通过视觉、嗅觉、味觉等感觉系统综合作用产生的,每一种行为反应的每一个环节均有一系列复杂的神经活动,是外部环境条件的刺激和内部生理状态综合作用的结果[65],而不是嗅觉、视觉或味觉系统的单一作用。王广利等[66]的研究认为天牛的交配行为多数由短距离或接触性信息素启动,虫体颜色、形状、大小以及虫体体表化合物和视觉均可影响其交配成功率; 李为争等[67]对天敌昆虫无翅桃蚜的研究表明其不仅可以识别寄主龟纹瓢虫的挥发物,而且可以识别龟纹瓢虫的体色。
了解天牛及其天敌的嗜好和排斥颜色、气味等,可为调控害虫及其天敌的种群结构,综合管理害虫及天敌打下理论基础。例如,在行道树或混交林中搭配不同颜色、气味的树种或者树干涂刷不同颜色[68],利用具有害虫排斥的颜色、气味的树种,干扰害虫的正常取食、交尾等行为活动,或者利用具有能够对天敌产生吸引的颜色和气味,聚集天敌昆虫等等[69]。总之深入开展花绒寄甲的感觉系统调控机制研究,了解各种感觉信号在花绒寄甲对寄主定位和识别机制中的关键作用,不仅可以丰富和完善该虫的感觉系统理论知识,而且对于今后调节林业生态系统中天牛及其天敌的种群结构有着重大的理论指导意义,同时也可以为研制与开发环境兼容性好、不杀伤天敌的害虫诱捕装置提供理论基础。
参考文献 ( References)
[1] 秦锡祥,高瑞桐. 花绒坚甲生物学特性及应用研究[J]. 昆虫知识,1988,25( 2) : 109 -112
[2] 王希蒙,任国栋,马峰. 花绒穴甲的分类地位及应用前景[J].西北农业学报,1996,5( 2) : 76 -79
[3] 杨忠岐. 利用天敌昆虫控制我国重大林木害虫研究进展[J].中国生物防治,2004,20( 4) : 221 -227
[4] 雷琼,李孟楼,杨忠岐. 花绒坚甲的生物学特性研究[J]. 西北农林科技大学学报: 自然科学版,2003,31( 2) : 62 -66
[5] 魏建荣,杨忠岐,马建海,等. 花绒寄甲研究进展[J]. 中国森林病虫,2007,26( 3) : 23 -25
[6] 黄大庄,杨忠岐,贝蓓,等. 花绒寄甲在中国的地理分布区[J].林业科学,2008,44( 6) : 171 -175
[7] 魏建荣,杨忠岐,牛艳玲,等. 花绒寄甲的分布与生态学习性补充调查[J]. 中国森林病虫,2009,28( 1) : 16 -18
[8] 魏建荣,杨忠岐,唐桦,等. 花绒寄甲成虫的行为观察[J]. 林业科学,2008,44( 7) : 50 -55
[9] 王亚红,来燕学,岑定浩,等. 花绒寄甲产卵行为和产卵量及对松褐天牛幼虫的寄生作用[J]. 环境昆虫学报,2011,33( 4) :517 - 522

天牛科 (Cerambycidae) 是鞘翅目 (Coleoptera) 中最大的科之一, 全世界约有45000种, 是鞘翅目 (Coleoptera) 昆虫中形态变异具多样性的类群之一。...

红缘天牛Asiashalodendri(Pallas)属鞘翅目Coleoptera天牛科Cerambycidae天牛亚科Cerambycinae亚天牛属AsiasSemenov,是一种危害严重,寄主广泛的蛀干害虫。有文献记载的红缘天牛的寄主有23种之多(孙逢海等,1994;王大洲和王金华,200...
竹林已成为我国南方重要经济植物,随着面积的不断扩大,害虫种类和数量也不断增多,特别是介壳虫的危害已经严重影响了竹类生长和经济收益,日前,竹农主要采取化学防治方法防治介壳虫,在一定程度上破坏了竹林生态平衡,关于竹林介壳虫及其天敌的相关报道...
高粱蚜[Melanaphissacchari(Zehntner)]别名高粱黄蚜、甘蔗黄蚜,属半翅目(Hemiptera)、蚜科(Aphididae),其主要寄主为高粱(Sorghumbicolor)、玉米(Zeamays)、甘蔗(sugarcane)和荻草(Triarrhercasacchariflora).高粱蚜发生世代短,繁殖快,每年...