生物化学论文


TRAF( TNF receptor associated factor) 家 族蛋白是一类具有相同 C 末端保守结构域的细胞内接头蛋白, 最早被发现能够介导肿瘤坏死因子受体( TNFR)家族的细胞内信号转导并因此得名,但后来的研究发现 TRAF 蛋白还能够与包括 Toll 样受体( TLR)家族、IL-1 受体家族以及视黄酸诱导基因蛋白 I( RIG-I) 样受体家族在内的多种受体蛋白相互作用并传递信号, 目前已经在哺乳动物中发现了 7 种 TRAF 蛋白。 TRAF 家族蛋白通过对细胞生存、增殖、分化和凋亡的调控,广泛参与包括免疫、胚胎发育、应激反应等多种生物学过程并在其中发挥着重要的作用[1]. TRAF4 是 TRAF家族蛋白中比较特殊的一个,本文对 TRAF4 的结构和功能做一综述。
1 TRAF4 蛋白的结构和一般性状
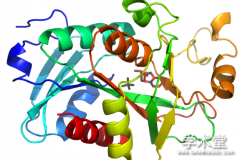
人类 TRAF4 基因编码一个 470 aa 组成、相对分子质量 54 kD 的衔接蛋白,在胞浆、细胞核以及细胞膜上均有定位。 TRAF4 蛋白主要包含 3 个结构域, 从 N 端到 C 端依次为一个 RING-finger结构域、7 个连续的锌指结构域、1 个 TRAF 结构域。 其中 TRAF 结构域是 TRAF 家族蛋白的特征性结构,除了 TRAF7,所有的 TRAF 蛋白家族成员都包含一个 C 末端 TRAF 结构域。 按照结构和功能的不同,TRAF 结构域又可以被分为 TRAF-N 和 TRAF-C 两部分,其中 TRAF-N 部分由多个α 螺旋组成, 在不同 TRAF 家族成员中保守性较差[2];而 TRAF-C 部分序列高度保守,折叠形成一个 8 链 β-三明治结构, 又被称作 MATH( meprinand TRAF-homology) 结 构域 ,主要介导 TRAF 蛋白的三聚化以及作为与信号分子直接或间接结合的位点( 图 1)。 三聚化的 TRAF 蛋白形成蘑菇样结构,3 段 TRAF-N 结构域的 α-螺旋平行排列在一起形成的卷曲螺旋结构,构成蘑菇样结构的柄,TRAF-C 构成蘑菇样结构的伞盖( 图 2) . 相比于其他 TRAF 家族成员,TRAF4 的 TRAF-N 结构域要更短,仅由 3 个 7 肽重复区构成,而其他 TRAF家族成员至少由 10 个以上 7 肽重复区构成。这就造成 TRAF4 蛋白与其他 TRAF 蛋白的结合能力很弱[3~5].除 TRAF1 蛋白外,所有的 TRAF 家族成员都包含一个位于 N 末端的 RING-finger 结构域以及几个连续的锌指结构域。 TRAF4 的 RING-finger结构域为两个锌指结构组成的 C3HC3D 型模体。

RING-finger 结构域在泛素途径中具有重要的作用, 能够将泛素酶与其底物蛋白相连, 发挥类似E3 连接酶的作用[6]. 3 个富含半胱氨酸的 CART结构域 ( 与 RING-finger 结构域和 TRAF 结构域相关的富含半胱氨酸结构域(cysteine rich domainassociated with RING and TRAF domain) 位 于RING-finger 结构域和 TRAF 结构域之间,每一个CART 结 构域均由一个外显子编码 , 包含两个HC3HC3 锌 指 模 体 . TRAF4 是 唯 一 具 有 3 个CART 结 构域的 TRAF 家 族成员 ,TRAF4 也 因此结构域曾被命名为 CART1. 后来的研究显示,这段富含半胱氨酸区域共由 7 个锌指模体构成,其中前 6 个锌指模体为 C2HC 型,而第 7 个为 C2H2型。 在 N 端第一个 CART 结构域中存在 1 个核定位信号,此外,在 RING-finger 结构域 N 端的上游也存在 1 个核定位信号,TRAF4 也是 TRAF 家族中唯一具有核定位信号的成员[3].
与 TRAF 蛋白家族的其他成员不同,TRAF4与包括 TNFR1、TNFR2、Fas、CD40 在内的大多数TNFR 家族成员不存在直接的相互作用, 与淋巴毒素 β 受体( LTβR)和神经生长因子受体( NGFR)仅存在微弱的相互作用, 这种作用能够产生怎样的生理作用仍然未知,并且 TRAF4 的高表达并不能像 TRAF2、TRAF5、TRAF6 那样激活 NF-κB[7].
TRAF4 主 要通过间接作用或形成复合体的方式在胞浆、胞膜和细胞核中迁移而发挥作用,其作用发挥涉及 JNK、ERK、PI3K/Akt、Wnt 等多条信号通路[8~11],截至目前 ,JNK 途 径被认为是其中最重要的一条,TRAF4 的大部分生物学效应伴随 JNK途径的激活, 但 TRAF4 单独激活 JNK 途径的能力很弱, 必须与其他蛋白如 MEKK4 等结合之后才具有显著激活 JNK 途径的能力[12].
进化分析显示,TRAF1、2、3、5 是在脊椎动物的进化过程中出现的, 与 TNFR 家族形成于同一阶段, 这一结果说明此 4 种 TRAF 家族蛋白的功能与 TNFR 家族蛋白的功能是相关的。 TRAF4 和TRAF6 在进化过程中出现较早,在低等的腔肠动物中即已发现 TRAF4 的同源类似物,并且无脊椎动物和脊椎动物的 TRAF4 同源蛋白在胚胎发育过程中都起着相似的作用[13],据此推断 TRAF4 可能是 TRAF 家族中古老的成员之一, 并且其发挥功能是 TNFR 非依赖的。
2 TRAF4 与肿瘤
TRAF4 最早在乳腺癌患者的转移淋巴结中被鉴定出来[14]. 之后的研究显示,除了乳腺癌,在包括肺癌、子宫癌、前列腺癌、结肠癌在内的多种肿瘤组织中都存在 TRAF4 蛋白的高表达,阳性率接近 1/2, 在肺癌中更是高达 60%. 并且 TRAF4高表达只局限在肿瘤细胞中, 周围的基质细胞则表达正常[15]. 一项基于 40 个独立的基因芯片筛查实验的整合分析显示,67 个基因的高表达可以作为肿瘤的特征性标记物,TRAF4 即为其中之一[16].
并且 TRAF4 高表达的乳腺癌患者对化疗药物的敏感性降低,预后较差[17]. 肿瘤中 TRAF4 蛋白的高表达常常伴随着亚细胞定位的异常。 虽然TRAF4 蛋白序列中存在两个可能的核定位信号,但内源性 TRAF4 的核定位目前仅在肿瘤细胞中发现。 在大多数正常细胞中,TRAF4 主要以膜相关的形式定位, 而在侵袭性肿瘤细胞中,TRAF4主要定位在胞浆中, 此外, 还有大约 20%的TRAF4 阳性肿瘤定位于细胞核中[15]. TRAF4 的高表达和定位异常有可能成为肿瘤诊断的一个标志物,并且这种异常表达有可能参与肿瘤的发生。
3 TRAF4 的生物学功能
3.1 TRAF4 的生理表达及其在发育中的作用
在成人几乎所有组织上皮的基底细胞层中都存在 TRAF4 蛋白的基础表达,并且各组织间表达水平差异不大[18]. 在成年小鼠的各组织中也检测到了基础水平 TRAF4 的表达,但其中表达水平最高的部位是中枢神经系统的海马、 嗅球以及小脑的浦肯野细胞[19].
在小鼠的胚胎中,TRAF4 广泛表达在各个发育中的器官,其中中枢神经系统、外周神经系统和感觉器官的神经组织表达水平最高。 TRAF4 缺陷的纯合子小鼠显示出明显的上呼吸道缺陷, 并导致约 30%的小鼠在胚胎期死亡,幸存的 TRAF4 缺陷小鼠表现出典型的共济失调症状, 并发生多种低渗透性改变,包括气管和中轴骨骼( 胸骨、肋骨、尾骨)的严重畸形,以及导致轻度脊柱裂表型产生的神经管闭合缺陷[19~21]. TRAF4 表达在少突胶质细胞分化的每一个阶段,TRAF4 敲除导致严重的中枢神经系统髓鞘不稳定和缺失。 此外, TRAF4能够通过 NgR/p75NTR/RhoA 信号途径抑制肌动蛋白细胞骨架重排从而促进轴突再生和抑制神经细胞的凋亡,对神经元具有保护作用[22].TRAF4 在小鼠中枢神经系统中的表达是受到调控的, 在小鼠出生后 4 周内,TRAF4 的表达明显下调。 在包括爪蟾、 斑马鱼、 果蝇在内的多种模式生物中,TRAF4 均表现出类似小鼠的表达模式[23~26].
综上所述,TRAF4 在大多数生物体内各器官广泛表达,但高表达于神经系统,对各系统发育,尤其是神经系统的发育起着不可或缺的作用。
TRAF4 调控发育的机制目前尚未明了,有研究显示,TRAF4 能够正向调控 TGF-β/BMP 和 TGF-β/nodal/activin 信号途径,此两个途径均与胚胎的发育密切相关[24]. 此外,还可能与 TRAF4 对细胞极性和凋亡的调控有关。
3.2 TRAF4 与细胞极性
TRAF4 有 多种不同的亚细胞定位 , 包括胞浆、细胞膜和细胞核。 外源表达的 TRAF4 主要存在于细胞间接触点部位的细胞膜上。 在体外培养的极化上皮细胞和正常人乳腺标本中,TRAF4 与紧密连接的标志性蛋白共定位于细胞与细胞连接点上。荧光漂白恢复实验显示 TRAF4 与紧密连接的相关性是高度动态的,TRAF4 在紧密连接和细胞浆间不断移位。这一结果说明 TRAF4 可能是正常上皮细胞间连接的信号传感器。与之相反的是,在分化差的上皮细胞肿瘤中,TRAF4 或者弥漫性分布在胞浆中, 或者在胞浆或胞核中呈现局灶性分布, 提示这种细胞间信号转导功能可能被削弱[8,9].
分节极性蛋白 Armadillo 是细胞间粘附的重要调节因子, 也是 Wnt 途径重要的下游信号分子。 在果蝇胚胎发育过程中,Armadillo 与 TRAF4发生直接的相互作用,以 TNFR 和 JNK 非依赖的方式精细调节中胚层细胞间粘合连接的形成[25].
在人类细胞中, 可能存在类似的相互作用。 β-catenin 是果蝇 Armadillo 在脊椎动物中的同源蛋白,最近的研究发现,TRAF4 可以与 β-catenin 结合并影响 β-catenin 的亚细胞定位, 高表达的TRAF4 可 以使 β-catenin 由 胞浆进入细胞核 ,进而通过激活 Wnt 途径促进乳腺癌细胞的增殖[11].TRAF4 和 β-catenin 在脊椎动物胚胎发育中的作用机制目前尚无报道。
TGF-β 信 号途径是细胞极性的关键调节因素,参与上皮-间充质细胞的转化和肿瘤的发生,被看作是肿瘤治疗的一个潜在靶点。 研究显示TRAF4 在 TGF-β 诱导的 SMAD 和非 SMAD 途径中起着关键的作用。 当 TGF-β 受体受到刺激的时候,TRAF4 被活化的 TGF-β 受体复合物募集,拮抗 E3 泛素连接酶 SMURF 的作用 ( TRAF4 作为SMURF2 的主要 E3 连接酶, 同时 TRAF4 本身可以作为 SMURF1 和 SMURF2 两种 E3 连接酶的底物)[24,27, 28],促进去泛素酶 USP15 向 TGF-βI 型受体( TβRI) 的募集。 这两个过程都能够促进 TβRI 在胞浆膜上的稳定性进而促进 TGF-β 信号途径。 此外,TGF-β-TRAF4 的相互作用可以激活 Lys63 相关的 TRAF4 多聚泛素化并激活 TGF-β 活化激酶( TAK1) 及其下游的 p38 MAP 激酶和 NF-κB 级联反应。 敲除 TRAF4 可以引起 E-cadherin 的高表达并抑制细胞的迁移[29]. TRAF4 对于 TGF-β 诱导的迁移、 上皮间叶转化和乳腺癌转移起着关键的作用。
最近的研究显示,TRAF4 分子具有一个独特的磷脂酰肌醇结合结构域, 通过与磷脂酰肌醇的相互作用,TRAF4 负向调节紧密连接并促进细胞的迁移。 TRAF4 的高表达可能通过破坏紧密连接的稳定性和促进细胞迁移来促进乳腺癌的进程[30].
目前看来,对细胞极性的调控是 TRAF4 最为重要的生物学功能,明确 TRAF4 在细胞极性调控中的作用机制对于理解 TRAF4 在发育和肿瘤发生中的作用具有重要的意义。
3.3 TRAF4 与凋亡
TRAF4 在凋亡中的作用存在两种截然相反的结论。 例如,敲除 TRAF4 可以诱导浦肯野细胞发生凋亡[21];TRAF4 高表达可以使 HEK293 细胞抵抗 anti-Fas 抗体诱导的凋亡[31];TRAF4 敲除的MCF-7 细胞被 TNF-α 刺激后, 早期凋亡细胞比例明显增高[32]. 其抗凋亡机制可能是 TRAF4 依赖其 TRAF 结构域与 p53 竞争性结合去泛素化酶HAUSP,从而抑制了 p53 的去泛素化 ,降低其稳定性[17]. 另外的一些研究却显示 TRAF4 具有促进凋亡的能力。 例如,在果蝇中,TRAF4 的同源蛋白DTRAF1 可 以 被 果 蝇 TNF/TNFR 同 源 物 Eiger/Wengen 的相互作用所募集,继而通过凋亡信号调节激酶 1( ASK1) 和 JNK 途径诱导果蝇细胞发生凋亡[33,34];在体外培养的 293T 细胞中 ,TRAF4 与神经生长因子受体 p75 NTR 直接作用,抑制 NF-κB 的激活,诱导细胞发生凋亡[35];在 头颈部肿瘤中,TRAF4 可以被 p53 家族成员( p63,p73,p53)正向调控[36,37];放射线诱导淋巴细胞凋亡和噪声诱导耳蜗细胞凋亡过程中都会出现 TRAF4 的高表达[38,39];在多种不同来源的细胞系中,TRAF4 的高表达可以诱导凋亡并且抑制集落形成, 并且与p53 的 状态无 关 . 这些看似 矛 盾 的 结 果 说 明TRAF4 在细胞生存、死亡的调控上具有细胞类型特异性。 TRAF4 对凋亡的调控很可能参与胚胎的发育和肿瘤的发生。
3.4 TRAF4 与活性氧(ROS)的生成
ROS 是正常氧代谢的副产物,具有多种重要的生物学活性,包括参与杀菌、炎症反应以及内皮细胞的激活 等 . NADPH 氧 化 酶 2 ( NOX2) 是NADPH 氧化酶中分布最广的一个亚型,通过质膜传递电子给分子氧产生超氧化物,参与杀菌、炎症和上皮细胞的激活。 P47phox 是 NOX2 的一个胞浆内亚基,也是目前已知所有 TRAF4 结合蛋白中与 TRAF4 作用的机制最为明确的一个。 研究证实,TNF-α 能够诱导 p47phox 发生 PKC 依赖的磷酸化,TRAF4 与磷酸化 p47phox 的 C 末端结合并携带 p47phox 向细胞膜转位, 这种转位能够诱导JNK 的 激活 , 并引起 NOX2 的 激活和 ROS 的 产生[40].
GPIb-IX-V 和 GPVI 是两种血小板糖蛋白受体,分别与血管性血友病因子( VWF)和胶原蛋白结合起始凝血过程。 在血小板活化过程中都会产生大量的 ROS,而 ROS 又会反过来调控血小板的活化,其机制未知。 研究显示,TRAF4 和 p47phox可 以 作 为 GPIb-IX-V 和 GPVI 的 结 合 伴 侣 ,GPIb-IX-V 和 GPVI 的胞浆段与 TRAF4 的 TRAF结构域、p47phox 的 SH3 结构域以及 Hic-5 形成一个复合体,从而使 NOX2 活化[41]. 这一发现为解释氧化还原信号通路与血小板活化间的关系提供了一个合理的依据。
3.5 TRAF4 与免疫
TRAF 家族是固有免疫信号通路中重要的接头蛋白, 通过与 TNFR 和 Toll/IL1 受体家族成员作用, 经 NF-κB 和 MAPK 的激活诱导免疫和炎症反应。但 TRAF4 似乎是一个例外, TRAF4 缺陷的小鼠 T/B 淋巴细胞、粒细胞、巨噬细胞、树突细胞分化和功能均正常, 仅表现为树突细胞的迁移能力降低[42]. TRAF4 不能与大多数 TNFR 和 TLR直 接 作 用 ,TRAF4 与 不 同 的 TLR 成 员 共 转 染293T 细 胞会对 TLR 介 导的 NF-κB 和 IFN-β 激活产生剂量依赖性抑制作用[43],提示其对固有免疫发挥的是抑制作用。 目前看来,TRAF4 发挥免疫抑制作用主要与 NOD2 途径及其与 TRAF6 的相互作用相关。
NOD2 (nucleotide-binding oligomerization do-main containing 2)是一种细胞内模式识别受体,在免疫平衡中起着重要的作用。 NOD2 在包括克罗恩氏病( 节段性结肠炎)、Blau 综合征等炎症性疾病的发病中起着重要的作用。有研究显示,TRAF4可能是 NOD2 途径的一个关键负调控子。 TRAF4是 IκB 激酶 α (IKKα) 的底物,其 Ser426 可以被IKKα 磷酸化, 磷酸化的 TRAF4 能够与 NOD2 蛋白结合并抑制 NOD2 诱导的 NF-κB 激活及杀菌作用。Ser426 位于 TRAF4 分子特有的一个膨大的β 凸起中,其他的 TRAF 家族蛋白均无此结构,这也解释了在所有的 TRAF 家族蛋白中,仅 TRAF4是对固有免疫起抑制作用[44,45].
作为 TRAF 家族中另一位古老的成员, 不同于 TRAF4,TRAF6 能够介导多种 TNFR 家族的信号转导,在固有免疫中发挥激活作用。 实验显示,TRAF4 可 与 TRAF6 发 生直接或间接的相互作用,拮抗 TRAF6 的生物学功能。 早期的研究即已发现,TRAF4 能够与 p47phox 协同, 通过对抗TRAF6 和 β 干扰素 TIR 结构域衔接蛋白( TRIF)的作用而抑制 TLR 介导的信号转导[43]. 新近的研究显示,TRAF4 与 TRAF6 的对抗作用还体现在IL-17 信号的转导中。 IL-17 是 Th17 细胞分泌的标志性细胞因子,IL-17 与 IL-17R 结合后募集IL-17 途径中两个关键信号分子肌动蛋白相关基因 1( Act1)和 TRAF6,形成 IL-17R-Act1-TRAF6信号复合物,进而激活 NF-κB 和 MAPK 途径,促进炎症性细胞因子分泌, 在多种炎症反应和自身免疫疾病的发病中起着重要作用。 TRAF4 和TRAF6 在 Act1 上 具有相同的结合位点 , 因而TRAF4 可以与 TRAF6 竞争结合 Act1, 从而抑制IL-17 信号转导。 将野生型 Th17 细胞转入 TRAF4缺陷小鼠能够在极短的时间内诱导自身免疫性疾病的发生[46].

此外,TRAF4 在免疫中的作用还与糖皮质激素诱导 TNF 受体( GITR)有关。 GITR 主要表达在T 细 胞 、B 细 胞和巨噬细胞表面 ,TRAF4 能 够与GITR 胞浆段发生间接作用, 通过 JNK 途径增强GITR 对 NF-κB 激活, 拮抗抑制性 T 细胞的功能而促进辅助性 T 细胞的激活[47]. 看起来 TRAF4 似乎对适应性免疫有一定的促进作用, 但这一结论并未得到体内实验的支持。
4 总结与展望
作为 TRAF 家族最古老的成员之一, 有关TRAF4 的研究却相对较少,TRAF4 蛋白功能的认知仍然模糊。 仅就目前的研究来看, 大多数情况下,TRAF4 与 TNFR 家族其他成员没有明显的直接作用或者作用很弱, 主要通过间接作用或形成复合体的方式在胞浆、 胞膜和细胞核中迁移而发挥作用, 其作用发挥涉及 JNK、ERK、PI3K/Akt、Wnt 等多条信号通路, 但目前尚无法准确归纳出TRAF4 在信号通路中的确切位置 . 多项有关TRAF4 的研究出现了诸多矛盾的研究结果,例如对凋亡的影响、对 NF-κB 以及对某些信号途径的作用等,这充分说明 TRAF4 的生物学功能复杂而多样。
作为一种衔接蛋白, 鉴定其相互作用成分及作用机制将是揭示 TRAF4 神秘作用的关键。 目前,已经鉴定出的 TRAF4 结合蛋白多达 20 余种,涉及膜受体、蛋白激酶、胞浆衔接蛋白、泛素途径相关酶、凋亡相关蛋白等( 表 1),其中很大一部分蛋白与 TRAF4 的互作机制尚未完全清楚,仍有待进一步研究。 同时有理由相信,与 TRAF4 相互作用的成分并非仅仅以上 20 余种蛋白,以目前的技术水平,一些短暂的、低水平表达蛋白以及修饰蛋白相互作用及间接相互作用的鉴定仍然存在一定的困难,并且与 TRAF4 作用的成分很可能并不局限于蛋白质。类似于转录因子的拼版理论,不同细胞内包含 TRAF4 作用成分的不同组合, 导致TRAF4 产生细胞特异的多样的生物学效应,这是造成 TRAF4 生物学效应复杂多样的最可能原因。分子间相互作用鉴定和定量技术的不断完善将会为 TRAF4 作用机制的研究提供有力支持。
参考文献(References):
[1] DEMPSEY P W, DOYLE S E, HE J Q, et al. The signalingadaptors and pathways activated by TNF superfamily[J]. Cy-tokine & Growth Factor Reviews, 2003, 14(3-4):193-209.
[2] ROTHE M, WONG S C, HENZEL W J, et al. A novel familyof putative signal transducers associated with the cytoplasmicdomain of the 75 kDa tumor necrosis factor receptor[J]. Cell,1994, 78(4):681-692.

研究发现,在哺乳动物基因组中,只有不到2%的转录产物具有蛋白编码功能,其余98%均为非编码RNA(noncodingRNA,ncRNA)[1,2].根据核苷酸序列的长度,ncRNA可分为短链ncRNA(short/smallncRNA)和长链LncRNA(longncRNA),二者之间并没有特...

蛋白质组学研究的主要内容包括在蛋白质水平上大规模地分析组织细胞的蛋白质表达水平、翻译后修饰、蛋白质间的相互作用等,从而揭示蛋白质的功能,已在疫苗筛选、指导治疗、临床药物开发及预后判断等领域发挥了重要作用。...
在自然环境下,两栖动物受到紫外线、病原微生物的伤害及捕食者的威胁.面对恶劣的自然环境,两栖动物体表进化出许多特殊的腺体,这些腺体能够分泌丰富多样的生物活性肽,如抗氧化肽、抗菌肽、抗病毒多肽等,其中的抗菌肽研究已取得诸多进展.有研究者仅从...

由于蛋白质控制体内重要的生理活动, 人们一直在研究蛋白质的生物作用机理。随着科学技术的发展, 蛋白质的研究越来越深入, 一些特殊蛋白质的生物学有了新的发展。而且一些动植物蛋白开始应用到人类的生产、医疗等方面。...
GAF结构域是以最早被发现含有这个结构域的蛋白质来命名的,由特异性的cGMP磷酸二酯酶 (cGMP-binding phosphodiesterases) 、腺苷酸环化酶 (adenylyl cyclases, AC) 和细菌转录因子FhlA (formate hydrogenlyase transcription activator) 的首字母构成。...

鱼类精巢原料量有限, 限制了鱼精蛋白防腐剂的广泛应用。目前人们正在尝试通过基因工程在微生物中以融合蛋白质的形式表达抗菌肽基因, 但仍存在表达产物少的问题。因此, 提高防腐抗菌肽类的产量、降低成本, 是目前必须解决的问题之一。...
膜蛋白是生物膜功能的主要承担者,根据膜蛋白的分布,膜蛋白可以分为三类:外周膜蛋白、整合膜蛋白和脂锚定蛋白。膜蛋白的功能具有多方面性,有些膜蛋白可以作为载体而将物质转运出细胞;有些膜蛋白是激素或者其他化学物质的专一受体;有些膜蛋白作为专一的...
硫化物是H2S,HS-和S2-的统称。由于硫化物与细胞色素aa3血红素卟啉环铁离子的可逆性结合进而阻止氧气与其结合[1];此外高浓度的硫化物可以导致细胞色素氧化酶构象的不稳定和亚基的降解[2];因此,硫化物能够破坏线粒体氧化呼吸链中电子传递,抑制ATP的产生。所...
1肌球蛋白的结构与功能肌球蛋白主要存在于平滑肌中,它是肌原纤维粗丝的组成单位。其分子形状如豆芽状,由多条重链与多条轻链组成。肌球蛋白的家族较大,目前发现的肌球蛋白有24种,但依据其来源又可分为传统的肌球蛋白和非传统的肌球蛋白,如传统的...
所有生物细胞的表面都覆盖着种类繁多的糖类化合物,例如O连N-乙酰葡糖胺,鞘糖脂,粘多糖、磷脂锚和脂-寡聚糖等,这些糖-蛋白质或糖-脂类复合物决定细胞的类型和调控细胞的生长,在环境发生变化时调节细胞做出应答反应。唾液酸就是众多糖类化合物...