生物技术论文


淀粉是由脱水葡萄糖单位构成的生物高聚物,大部分是以淀粉颗粒的形态存在,不论来自何种植物或组织,都包含2种主要的多糖,即直链淀粉和支链淀粉。直链淀粉通常被定义为线性长链分子,是由脱水葡萄糖单位经过α-1,4糖苷键连接而成的,而支链淀粉是由葡萄糖经过α-1,4糖苷键和α-1,6糖苷键连接而成,即由许多短链在还原端通过α-1,6糖苷键连接在一起,使支链淀粉具有高度的分支。由于直链淀粉和支链淀粉的结构不同,且在淀粉中的含量不同,所以决定各类淀粉的结构和性质的差异。
淀粉颗粒是一种半结晶的聚合物,其包含有结晶区和无定形区。淀粉颗粒中形成结晶区的主要为支链淀粉,支链淀粉分子相互聚集形成的双螺旋结构是结晶的基础。而直链淀粉是组成淀粉颗粒无定形区的主要成分。然而在高直链淀粉品种下,淀粉分子是如何影响淀粉颗粒的结构的,目前仍不是十分清楚。
这主要是由两方面的原因造成的:一是缺乏更为有效的技术;二是淀粉降解模型的设计上需要更科学化。
虽然已有许多通过酶降解体系来研究淀粉结构,例如分析降解前后淀粉颗粒形态和分子结构的变化等,然而采用不同的方法和技术分析淀粉的结构仍显得十分必要。本试验采用复合微生物发酵的方法结合色谱检测技术,试图从不同的角度来研究淀粉的精细结构。
1 试验材料和方法
1.1 试验材料
普通玉米淀粉,购于Sigma公司(美国);高直链玉米淀粉,来源于国民淀粉公司(澳大利亚);其他试剂都为化学分析纯度。
1.2 试验方法
1.2.1 淀粉蒸煮
玉米淀粉的蒸煮处理:将淀粉与蒸馏水相混合制成水分含量为60%的淀粉乳,置于高压锅中于121℃处理30 min,冷却后重复高压处理两次。取出后冷却,冷冻干燥,将干燥后得到的固体经粉碎机粉碎后过0.5 mm的筛网。
1.2.2 微生物发酵处理
利用微生物发酵法降解普通玉米淀粉和高直链玉米淀粉,对其降解前后的分子结构进行分析。具体的试验过程为:利用磷酸缓冲液将来源于健康人体粪便制备成固形物10 g/mL的肠道微生物混合菌液,用于体系发酵。取200 mg淀粉样品底物加入到含有9 mL发酵介质的发酵瓶中并水合1 h(温度控制在4 ℃)。发酵介质包含以下物质(每1 L蒸馏水):2.5 g胰蛋白酶、125 μL矿物质溶液(每1 L蒸馏水中132 g CaCl2·2H2O,100 g MnCl2·4H2O,10g CoCl2·6H2O,80 g FeCl3·6H2O)、250 mL缓冲溶液(每1 L蒸馏水中4 g (NH4)HCO3、35 g NaHCO3)、250 mL矿物质溶液(每1 L蒸馏水中5.7 g Na2HPO4,0.6g MgSO4·7H2O)和1.25 mL刃天青溶液0.1 g/mL。取33.5 mL灭菌后的还原性溶液(每1 L蒸馏水中6.25 g半胱氨酸盐酸盐,6.25 g Na2S·9H2O,40 mL NaOH)添加到1 L发酵介质中,发酵介质的pH调整到7.2。每个发酵瓶接种1 mL的微生物混合菌液,每毫升混合菌液中含有1×1010活菌,最终保证达到每个发酵瓶中含有1×109活菌。每个试验瓶中用氮气喷雾,并用石蜡密封作为防止泄露的预防措施。试验瓶在37 ℃振荡中培养24 h,发酵结束后立即冷冻贮藏。
1.2.3 利用HPLC分析2种淀粉的分子组成取不同发酵时间段的发酵液冷冻干燥,取固形物50 mg放入25 mL离心管中,向其中加入8 mL二甲亚砜(DMSO)和水的混合液(DMSO与H2O的体积比为90∶10)。每只离心管封盖,置于沸水浴中加热30min,冷却后加入20 mL无水乙醇,封盖后均匀震荡3min,静置5 min后于3 000 r/min条件下离心,弃去上清液,加入20 mL无水乙醇再次震荡离心。再次用乙醇洗涤重复操作得淀粉沉淀物。
取15 mg该淀粉沉淀物,用0.5 mL 0.2 mol/L的NaOH溶液溶解该淀粉,震荡混合10 s。向所得溶液中加入0.5 mL 0.05 mol/L的乙酸钠缓冲液(pH 4.0)进行中和,然后加入离子交换树脂(0.20 g,BioRad AG?
501-X8,Hercules,USA),并于50 ℃水浴锅中保持1.5h。于10 000 r/min 离心10 min后,得上清液,进行HPLC分析(LC 1150,GBC Instruments, Vic, 澳大利亚)。色谱柱为UltrahydrogelTM250柱和保护柱(7.8mm×300 mm,Waters,日本),检测器为蒸发光散射检测器(ELSD)(ALLTech,Deerfield, 美国)。流动相为0.05 mol/L的醋酸铵缓冲溶液(pH 5.2)。
1.2.4 扫描电子显微镜(SEM)观测淀粉颗粒的形态。取不同发酵阶段的发酵液,经冷冻干燥、粉碎后,均匀地撒在贴有双面胶的样品面盘上,用Hitachi1B-3离子涂布器喷金固定,然后利用扫描电子显微镜(SEM)观察并记录颗粒形态。发酵前的淀粉样品直接用SEM观察并与前者进行比较。每种淀粉样品至少记录3次颗粒形态。
2 结果与讨论
2.1 淀粉分子的色谱图
2.1.1 发酵前2种淀粉分子色谱图的比较
对于普通玉米淀粉,其分子主要由2个组分组成,即大分子的支链淀粉和相对分子量较低的直链淀粉,其中支链淀粉占较大比例;而对于高直链玉米淀粉,液相色谱图(图1b)显示其是由3个组分所组成,分别为大分子的支链淀粉、相对分子量较低的直链淀粉以及更低分子量的直链淀粉。为了便于理解,本试验将直链淀粉的这两个组分分别称为高分子直链淀粉分子和低分子直链淀粉分子。这种分子结构是现阶段本试验研究的最新发现,特别是目前人们对于低分子直链淀粉的分离、结构测定等方面研究还不充分,对于该组分又是如何影响淀粉的结构和降解特征的研究也有待于更深一步进行。
2.1.2 原淀粉(未蒸煮)在发酵过程中淀粉分子色谱图的变化
对于普通玉米淀粉而言,在发酵过程的前半段(即从0~12 h),直链淀粉和支链淀粉比例似乎变化不大。这可能说明微生物同时利用直链淀粉和支链淀粉2个组份,即同时利用淀粉颗粒的结晶区和无定形区。而随着发酵过程的进一步进行,直链淀粉表现出较强的易降解性,这个结果可能提供两个方面信息:
一是直链淀粉在淀粉颗粒中的分布状况;二是随着发酵的进行,微生物产生了大量的分解酶,酶对淀粉表面及内部侵蚀,这就为发酵的后半阶段(12~24 h)微生物有效利用非结晶区提供了前提。这也就是为什么在发酵的前半段淀粉颗粒中支链淀粉和直链淀粉的比例变化不大,而在发酵的后半段直链淀粉则被快速地利用。
而对于高直链玉米淀粉的发酵,则呈现出极其特异的现象。从色谱图(图1b)所呈现出的变化来看,高直链淀粉分子在发酵的前半段就呈现出了极强的被利用状况,相反低分子的直链淀粉则表现出了与支链淀粉相类似的低发酵特性。这一点为重新认识淀粉的结构特征特别是对新型淀粉品种提供了非常有用的信息。众所周知直链淀粉是形成非结晶区的主要成分,其被降解酶快速有效的利用已经达成共识。而低分子直链淀粉的存在以及其对淀粉颗粒结构的影响可能使人们对淀粉的结构有重新认识,也就是说低分子直链淀粉在淀粉颗粒中的分布状态、分子排列情况等性质都需要做深入的研究。
2.1.3 蒸煮后淀粉在发酵过程中淀粉分子色谱图的变化
蒸煮过程不仅可以破坏淀粉颗粒表面的形貌,还破坏淀粉颗粒的内部结构,主要体现在两个方面:一是淀粉内部的结晶区被破坏,支链淀粉的分枝双螺旋的有序排列被打乱,在水溶液状态中形成大分子的缠绕状态;另一个变化是无定形区中直链淀粉的重排,直链淀粉在回升过程中形成更为有序的排布状态,这种变化会在其被微生物的利用过程中体现出来。对于普通玉米淀粉,被打乱有序排列结构的支链淀粉表现出被快速利用的现象,图谱(图1c)显示24 h发酵后基本上未检测出支链淀粉分子,这说明大分子的支链淀粉即使发生了在回升过程中的分子重排,但仍快速地被微生物利用。其图谱中所检测出的小分子物质可能是由高分子直链淀粉被微生物利用后所产生的低分子直链淀粉。而对于蒸煮后的高直链玉米淀粉而言,除了高分子直链淀粉所呈现出的高被利用状况外,与普通玉米淀粉一样,其所含的少量的支链淀粉也被微生物消化利用,而低分子直链淀粉分子则呈现出在发酵后残留淀粉颗粒中相对积累现象。这说明蒸煮过程中相对于高分子直链淀粉分子,低分子直链淀粉更易趋于排列成更为有序的状态,从而体现出了其慢速发酵性。【图.略】
2.2 发酵过程中淀粉颗粒形态变化
2.2.1 原淀粉颗粒在发酵过程中的形态变化
通过SEM可以很清楚的显示淀粉颗粒被微生物利用的过程。由图2可见2种原淀粉颗粒发酵过程中形态的变化大不相同。对于普通玉米淀粉而言,微生物的利用是从表面和内部同时进行的,这说明这种淀粉的内部结构比较疏松。也有些报道称这种淀粉表面及其内部有多孔的通道,在微生物发酵过程中可分泌多种酶系,在淀粉酶系作用下淀粉颗粒表面变得粗糙且多孔,这就为微生物在其表面吸附和进入内部提供条件。这也为微生物进一步发酵提供新的起点,即在前面所述的微生物及其分泌的降解酶特别是淀粉酶系在此阶段开始快速利用淀粉颗粒中的无定形区,这和本试验的排阻液相色谱所显示在发酵的后期(12~24h)直链淀粉分子被快速利用的结构相吻合。
而对于高直链玉米淀粉而言,其微生物的利用途径则完全不同。电镜图(图2b)显示淀粉颗粒被微生物利用的过程是逐步由表面向内扩散,这种利用方式可使淀粉颗粒的尺寸逐步变小,这种降解方式可能与其比较紧密的表面和内部结构有关联,同时这种利用模式和用淀粉酶系测试结果相类似(结果将在近期发表)。因为高直链玉米淀粉中绝大部分是由直链淀粉所组成的,由本试验色谱图(图2b)结果可知,高分子直链淀粉是逐步被降解消化的,结合相对应的SEM图可能说明高分子直链淀粉主要是排布在淀粉颗粒的表面。但是各类淀粉分子是如何精确地分布在淀粉颗粒中仍要进一步的研究。
2.2.2 蒸煮后的淀粉颗粒在发酵过程中的形态变化
蒸煮导致淀粉颗粒的破碎,在发酵过程中对于普通玉米淀粉而言,在发酵前期形成片状骨架结构(图3a)。这可能是由于蒸煮过程中发生了淀粉分子的迁移以及相分离,也就是说容易被微生物所利用的淀粉分子凝聚集中于某些区域,从而被微生物降解后所剩余的淀粉形成了特定的形态,发酵后期微生物进一步利用残留骨架,最后形成不规则的形态。
而对于高直链玉米淀粉而言,在发酵前期不形成这种骨架结构,但是其表面呈圆滑形态(图3b)。这可能是因为其结构紧实,微生物及其产生的酶系由表面逐步利用这类淀粉,从发酵初期到发酵结束似乎均始终以此种由表及里的发酵模型进行。
3 结论
采用复合微生物发酵方法来研究淀粉的精细结构,这有区别于传统的单酶或双酶作用体系来研究淀粉的结构,前者具有独到之处,这是因为淀粉颗粒虽然绝大部分是淀粉,但它仍存在少部分的脂肪、蛋白质、非淀粉质多糖与其它杂质,这些杂质的存在也会对淀粉的降解产生外部因素的影响。
本试验非常有效地揭示出不同淀粉分子发酵的动力学,其中已经证实高分子直链淀粉分子(无论存在于普通玉米淀粉还是存在于高直链玉米淀粉)容易被微生物所利用。排阻液相色谱结果显示高直链玉米淀粉中的低分子直链淀粉分子具有很强的抗降解性,这就为研究淀粉中各种淀粉分子在淀粉颗粒中的排布情况提供了有价值的信息。
参考文献:
[1] FRENCH D. Fine structure of starch and its relationshipto the organization of starch granules[J]. Journal of theJapanese Society of Starch Science, 1972(19): 8-25.
[2] HIZUKURI S, KANEKO T, TAKEDA Y. Measurement ofthe chain length of amylopectin and its relevance to the originof crystalline polymorphism of starch granules[J]. Biochimicaet Biophysica Acta, 1983(760): 188-191.
[3] HAN X Z, HAMAKER B R. Amylopectin fine structureand rice starch paste breakdown[J]. Journal of Cereal Science,2001, 34(3): 279-284.

水体富营养化主要是由于人为原因导致水体中氮、磷等营养元素远远超过水体的自净能力,从而在水体中富集,水体中的藻类因为有了丰富的营养物质得以急剧增长,细菌也大量繁殖。...

污泥堆肥化技术是20世纪60年代后期迅速发展起来的一项生物技术。它通过借助各种微生物在特定的环境中对有机物进行分解,将污泥改良成稳定的腐殖质,用于改良土壤或肥田,在实际应用中可以达到无害化、减量化、资源化的效果,经济、实用、不需要外加能源...

为了能够检测出食品中的微生物,相关工作人员将越来越多的新技术引入其中,以便能够做好食品检测工作,保障食品的安全性。...
酶是一种可以高效控制特定化学反应的一种通用的生物催化剂,在实际应用中通常将酶作固定化处理。固定化酶由于其高稳定性、可重用性、易分离等性质被广泛地应用于化学、生物、农业和医药等领域[1-3].酶固定化方法主要包括物理吸附法、共价结合法、包埋...
随着污水处理厂的大规模增加,污水得到妥善的处理,而集中了大部分污染物的脱水后污泥的处置成了一个巫待解决的难题。传统的处理方式一般是填埋或焚烧,填埋势必造成长期的二次污染,使得污水处理的努力化为乌有,而焚烧会带来一些始料未及的环境污染。经过研究人...
广式高盐稀态酿造酱油工艺有以下3个特点:(1)采用晒露发酵,依赖于环境温度和光照;(2)酿造过程中不添加风味菌种,而是通过酿造环境中多年积累并形成体系的多种产香酵母和细菌自然接种发酵;(3)自然出油,不使用压榨设备。这些特点赋予了广式酱油成本较...
近年来,组学技术(基因组学、转录组学、蛋白质组学和代谢组学)和基于组学的数学模型在生命科学领域变得越来越流行,而基于组学分析的系统发酵优化和遗传改造策略在产油微生物中也得到广泛应用。...
微生物群落是地球上生命基本元素(C、N和S等)进行生物地球化学循环的主要驱动力,与人类健康、环境保护以及工农业生产等密切相关。近十年来,随着高通量测序的广泛应用,微生物组学成为新兴概念和热点。微生物组与不同的生存环境结合,诞生人体微生物组,宿...
微生物能够产生多种具有生物活性的次级代谢产物,其中大部分活性次级代谢产物来源于放线菌[1],这些物质具有抗菌、抗肿瘤、杀虫、免疫抑制和免疫激活等功效,被广泛地应用于医药业、农业、兽业、食品工业等领域。近年来,随着抗生素耐药菌的不断出现,深入研...
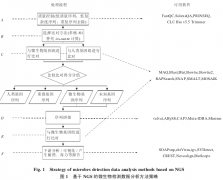
除了计算技术方面的优化,生物技术的创新也能给微生物检测方法带来革新。未来生物技术的发展使测序技术向着高通量、低成本、长读取长度的方向发展,目前已接近实用的第三代测序技术具有超长读长的特点,一旦应用将会极大地改善微生物检测方法的流程。对微生物...