生物技术论文


代谢组学研究生物体内源代谢物的种类、数量及其在内外因素作用下的变化规律,是系统生物学的重要组成部分,也是继基因组学、转录组学和蛋白质组学之后迅速发展起来的新兴学科。代谢组学定义有20世纪90年代末Nicholson提出的"metabonomics"[1]和 21 世纪初 Fiehn 提出的"metabolomics"[2]两种,两者在拼写及侧重点稍有不同,但本质上都是指从整体上研究生物体的代谢产物。目前,对于代谢组学的定义为"对某一生物、组织或细胞中所有低分子量(通常指分子量<1000)代谢产物进行定性和定量分析的一门科学。代谢产物作为生物体在内外因素作用下基因转录和蛋白表达的最终结果,是生物体表型的物质基础。同时代谢产物又能影响或调节基因的转录和蛋白的表达和活性。代谢组与基因组、转录组、蛋白组相比更接近生物体的表型,基因组和蛋白组的微小变化可以在代谢组层面得以体现和放大[3].因此,代谢组学研究越来越受到广泛关注,代谢组学手段在解析生物系统及基因功能等方面也发挥着越来越重要的作用。
在整个代谢组学的研究中,植物代谢组的研究占有重要的地位。作为一类固着性生物,植物可以产生种类繁多的代谢产物,总数在20万~100万种之间[4].植物代谢物大体可分为初生代谢物和次生代谢物两大类。初生代谢物为维持植物生命活动和生长发育所必须[5],次生代谢物则更多的参与植物抗病、抗逆等环境应答[6,7].植物代谢物种类繁多,结构各异、含量差异很大,这一方面使植物成为了研究代谢物生物合成及其调控的理想材料,同时植物代谢物的复杂性也对植物代谢组学研究提出了严峻的挑战[8,9].近年来,随着代谢组学分析技术的发展,特别是基于质谱及核磁共振的代谢谱分析的发展,代谢组学研究的内容不断扩展[10,11].另外,通过代谢组学与其他组学技术(如转录组学、基因组学)的整合[12],植物代谢组学在功能基因鉴定、代谢途径解析及自然变异的遗传分析等方面都取得了较大的进展[13~17].本文综述植物代谢组学技术及其研究现状和前景。
1 植物代谢组学技术
与其他组学技术只需分析相同或相似的特定类型对象不同,代谢组学特别是植物代谢组学所要分析的对象种类繁多、理化性质各异、浓度范围分布极广,依靠单一的分析手段难以对全部植物代谢物进行无偏检测。目前,基于色谱(气相、液相、毛细管电泳)、质谱(mass spectrometry,MS)、核磁共振(nuclear magnetic resonance,NMR)、傅里叶变换-红外光谱(Fourier transform-infrared spectrometer,FT-IR)等分析平台在植物代谢组学研究中都得到了广泛应用,其中研究最深入、应用最广的是核磁共振和色谱-质谱联用两大技术平台。
1.1 基于核磁共振平台的代谢组学技术
核磁共振技术是代谢组学研究中应用最早、最为常见的技术之一[18,19].核磁共振是一种基于具有自旋性质的原子核在核外磁场作用下吸收射频辐射而产生能级跃迁的谱学技术,常用的有氢谱(1H-NMR)、碳谱(13C-NMR)、磷谱(31P-NMR),其中1H-NMR应用最为广泛。核磁共振技术样品用量小、几乎不需要进行样品前处理,而且能够提供代谢物准确的结构信息。核磁共振技术的不足在于其检测灵敏度低、分辨率不高,难以检测丰度较低的代谢物。近年来,随着950 MHz高场强核磁共振及超低温探头技术(cryoprobe technology)等的发展,检测分辨率和灵敏度得到了较大的提高[20],加之其无损伤性、分析速度快、代谢物检测的无偏性等特点,核磁共振技术在植物代谢组学研究中应用将更加广泛。
1.2 基于色谱-质谱平台的代谢组学技术
基于气相色谱-质谱(GC-MS)技术的代谢组学平台主要用于分析热稳定、易挥发、能气化的小分子代谢物,分离效率高,具有较高的分辨率和灵敏度[21,22].另外,气相色谱-质谱检测结果可通过与标准谱图库比对获得代谢物结构信息,易于对代谢物进行定性。利用多维色谱技术发展的全二维色谱质谱联用技术,如全二维气相色谱-飞行时间质谱(GC×GC-TOF/MS)能够进一步提高色谱分离效果和峰容量,同时通过提高灵敏度和聚焦效果增加代谢物的检出量,展示了对于复杂生物学样品分析的优越性能。气相色谱-质谱发展较为成熟,已经成为植物代谢组研究中广泛应用的分析方法,是目前复杂样品中代谢物分析的主要定性和定量手段之一[23].
基于液相色谱-质谱(LC-MS)的代谢物检测方法检测限宽、灵敏度高,对待测组分的挥发性和热稳定性没有要求,通过选择不同类型色谱柱可以实现从极性到非极性的各种代谢物特别是各种次生代谢物的检测,一般也不需要衍生化的复杂的前处理,因此在植物代谢物检测技术的发展中越来越受到重视[24].传统的液相色谱-质谱方法有靶向代谢组学(targeted metabolomics)[25,26]和非靶向(non-targeted/untargetedmetabolomics)[27,28]两大类。靶向代谢组学方法只能对少数(一般少于100种)已知代谢物进行定性和定量检测,但其具有灵敏度高、定量准确的特点。与之相反,非靶向代谢组学方法能够同时检测数百乃至数千种代谢物(包括已知和未知代谢物),但其灵敏度较之前者减低1~2个数量级,定量准确性也相对较差[29].针对上述两种方法的优缺点,近年来相继发展出能够同时定性、定量数百种已知代谢物和定量近千种已知及未知代谢物的广泛靶向代谢组学分析方法(widely-targeted metabolomics)。
笔者开发的一种新的广泛靶向代谢组学分析方法,能够在30 min内定量超过800种已知和未知代谢物[28].该方法与靶向方法具有相同的灵敏度和定量准确性,同时结合了非靶向方法代谢物种类覆盖广泛、检测数量大的优点,在一定程度上代表了代谢组学检测手段的发展趋势,特别适合对大量试样的高通量、广覆盖的代谢组学分析,具有非常广泛的应用前景。在液相色谱技术方面,超高效液相色谱(ultra performance liquid chromatography,UPLC)及超高压系统(压力大于105 kPa)等新型液相色谱技术的应用能显着改善色谱分离度,与质谱联用时基质干扰减小,在提高检测灵敏度的同时大大缩短了分析周期。因此,超高效液相色谱-质谱替代传统液相色谱-质谱特别适用于微量复杂混合物的分离和高通量代谢组学分析,在植物代谢组学研究中得到了快速而广泛的应用。
毛细管电泳技术是基于带电分子在电场中的泳动速度差异而实现代谢物分离分析的技术。毛细管电泳技术与其他色谱技术互补,特别适于利用广泛靶向代谢组学手段对离子型代谢物(如核苷酸、有机酸、氨基酸、磷酸化的寡糖等植物初生代谢物)的分离和分析[30,31].毛细管电泳质谱(CE-MS)将毛细管电泳快速、高效、分辨率高、重复性好等特点和质谱分析灵敏度高、速度快等优点相结合,在强极性代谢物,特别是带电代谢物的分离分析中具有广泛的应用前景[32,33].
1.3 代谢物的注释
除了代谢组学分析平台方面的发展以外,代谢物的结构鉴定或注释也是目前代谢组学分析中的一个重点和难点问题。到目前为止,代谢组学主要集中在对已知代谢物的研究方面。值得注意的是,随着代谢组学技术的发展,特别是非靶向和广泛靶向技术的发展,代谢组学分析过程中获得了大量"未知代谢物"的定量信息。这些"未知代谢物"能够在代谢组学分析中被重复、稳定的定量检测,但是其化学结构未知。虽然它们可能是生理、病理及逆境响应中的重要生物标志物,或是直接在上述过程中发挥作用,但是由于缺乏相应的结构信息,很难对这些未知代谢物进行深入研究,进一步的应用也受到很大限制。由于代谢物结构的解析需要对代谢物进行分离、纯化,并获得其核磁共振、光谱等综合特征,难以达到组学对分析通量的要求,目前代谢组学中未知代谢物的解析工作主要集中在其结构注释方面。
利用飞行时间质谱、傅里叶变换质谱及轨道阱质(orbitrap)谱等可以提供代谢物的精确质量,多重质谱(MSn)能够提供代谢物完整的碎片离子信息。通过对这些信息的分析和处理,能够得到代谢物的相对分子质量、元素组成及官能团等结构信息,并进而利用生物信息学手段进行代谢物结构注释。基于此,近年建立了一系列公共代谢组学数据库,如MassBank[34]、KNApSAcK Core Database[35]、Plant MetabolomeDatabase(PMD)[36]、Metlin[37]、Golm Metabolome Database(GMD)[38]、Platform for RIKE Metabolomics(PRIMe)[39]、MeltDB[40]及 Madison Metabolomics Consortium Database(MMCD)[41]等。部分公司也提供商业化的代谢组学数据库,如Agilent Fiehn GC-MS Metabolomics Library、Agilent MetlinPersonal Metabolite Database、Wiley Registry/NIST Mass Spectral Library等。同时,也开发出一些用于解析质谱和核磁共振数据及通过整合代谢途径信息进行代谢物注释的工具。
近年来还发展了一种利用系统生物学手段进行未知代谢物结构鉴定及注释的功能代谢组方法。该方法利用正向遗传学手段获得未知代谢物与已知功能基因的关联信息,结合代谢物之间的相关性及代谢途径/网络分析,对未知代谢物进行结构注释,或进一步通过与标准品的比对进行结构确证[42].笔者将上述方法与质谱获得的结构信息整合,注释水稻中包括类黄酮、维生素、萜类等超过160个未知代谢物,并鉴定了其中部分代谢物的结构[13],取得了良好的效果。
2 植物代谢组学研究进展
植物代谢物是植物生物体生理状态在代谢水平的体现。任何生长发育及环境因子等内、外因素对植物体的扰动都会引起植物代谢物浓度或代谢流的变化,且大多数情况下这种变化不是表现为一个或少数几个代谢物的变化,而是多个代谢物甚至是多个代谢途径的改变。由于代谢组学从整体上检测代谢产物的变化,因此被越来越广泛地应用于植物生物学及相关领域的研究中。限于篇幅,本文仅对代谢组学在植物生物学基础研究中的进展进行简要介绍(图1)。
2.1 用于植物代谢物积累模式研究
通过代谢组对植物代谢物积累模式的研究不仅可以了解植物代谢物的不同时空积累模式,而且还可以研究其成分及含量在不同环境条件下的变化,为进一步解析其生物合成及调控奠定基础。代谢物的时空积累模式以研究具有生理、生物活性的次生代谢物最具代表性。类黄酮(flavonoids)是植物中普遍存在的一类次生代谢物,广泛参与植物发育、生理等过程,对人体也有明显保健作用,具有广泛生理活性[43].
笔者所在的研究组通过靶向代谢组学手段检测了水稻中超过90种类黄酮物质的时空积累模式,结果表明其中绝大多数在营养生长阶段的叶中的含量最高。进一步研究表明,不同类型的类黄酮在水稻籼粳两个亚种间的积累模式也存在着明显的差异[44].酚胺(phenolamides)是广泛存在于植物中另一类参与植物生长及胁迫响应的次生代谢物[45].笔者所在的研究组发现,与类黄酮不同,酚胺在根中高量积累,同时其在籼粳亚种间的积累模式也与类黄酮正好相反[13,46].由于上述两类物质均在植物逆境响应过程中发挥作用,推测它们在水稻不同组织、不同亚种间的特异积累可能与水稻不同组织、不同亚种所受胁迫类型的不同有关。硫代糖苷(glucosinolates)是一类十字花科植物中特异的含硫代谢物,在抵抗生物胁迫过程中发挥重要作用[47].对模式植物拟南芥中硫代糖苷的研究不仅表明其积累具有明显的时空特异性,还受到外界环境因子的调控,同时与不同生态型的地域适应性密切相关[16,48].上述代谢组学研究结果从一个侧面说明了植物代谢物,特别是次生代谢物在植物的进化、亚种分化等方面的作用。
除此之外,通过对胁迫条件下植物体内各代谢产物进行定性及定量分析,从而发现植物逆境下代谢调控规律及网络,也是植物代谢组学研究的一个重要方面。以非生物胁迫为例,植物在非生物胁迫下会启动一系列生理、生化响应,代谢组学通过监测胁迫下代谢物种类、含量变化为揭示这些复杂响应提供了一个极好的平台。Bowne等[49]研究了不同旱胁迫抗性的小麦品种在旱胁迫下的代谢变化。笔者所在的研究组利用广泛靶向的代谢组学方法研究旱胁迫下水稻叶片的代谢物变化,结果表明旱胁迫能够显着提高脱落酸(abscisic acid)和包括脯氨酸(proline)在内的多种氨基酸、五羟色胺及其衍生物、部分酚胺及黄酮的积累,而溶血磷脂胆碱(lysophosphatidylcholines,LPCs)的含量则明显下降[28].
Yamakawa等[50]发现水稻内源葡萄糖及ADP葡萄糖的含量在高温胁迫下大量积累。大量研究揭示植物体内腐胺、亚精胺、精胺等多胺的含量与植物抗非生物胁迫能力显着正相关[51~53],提示多胺在提高植物逆境抗性中发挥重要作用[54].
2.2 用于代谢物积累的遗传基础研究
解析遗传变异如何控制性状的表达是现代生物学研究的基本目标之一。由于植物代谢物对于植物及人类的重要性,植物生物学家一直致力于揭示调控植物代谢背后的遗传基础[12,55,56].植物样品中代谢物含量作为代谢性状(m-traits)往往也是受到多基因控制的数量性状,代谢性状的自然变异材料为研究其遗传调控提供了难得的遗传资源。通过数量性状座位(quantitative trait loci)分析代谢性状的自然变异有助于揭示控制这些变异的遗传学基础,解析代谢性状形成的遗传结构。目前绝大多数代谢性状的遗传学基础研究都是通过两个或少数几个亲本通过杂交或回交获得的连锁群体(linkage population)。例如,Schauer等[57]利用GC-MS技术进行番茄片段替换系(野生型番茄Solanum pennellii基因组片段由栽培种Solanum lycopersicum M82替换)代谢谱分析,共鉴定了控制74个已知代谢物的889个mQTLs.Matsuda等利用一个水稻回交群体(back-crossed inbred line,BIL)进行非靶向代谢组分析及代谢物QTL定位,获得了802个代谢物QTLs(mQTLs)[58].对于拟南芥类黄酮[59]、硫代糖苷[16]等次生代谢物mQTL定位也获得了部分具有显着效应的位点。
传统的mQTL定位都是基于限制性片段长度多态性(restriction fragment length polymorphism,RFLP)、简单序列重复(simple sequence repeat,SSR)等低密度标记[15,60].利用这些标记定位了大量的代谢相关的遗传位点。但是由于标记密度过低,很难直接获得决定代谢物含量的候选基因[61].笔者所在的研究组首次将高密度连锁图谱应用mQTL研究中:利用广泛靶向的代谢组学手段对一个获得高密度连锁图谱的水稻重组自交系(recombinant inbred line,RIL)群体的两个组织分别进行代谢谱分析及代谢物QTL(mQTL)作图,共获得了900个代谢性状超过2800个mQTL[62].研究表明,水稻代谢组遗传调控模式在不同组织中存在着很大的差异,具有明显的组织特异性。对mQTLs的全基因组分布研究发现,代谢组的遗传调控存在明显的热点区域,且热点区域在不同组织中也不尽相同。在此基础上通过对部分高精度(<100 kb)、大效应(解释超过40%变异)位点的分析,获得了24个控制代谢物含量变异的候选基因,并对其中部分基因进行了功能鉴定[62].
利用连锁群体进行mQTL定位在植物代谢组学研究中已经得到广泛应用。但是该方法存在着群体重组位点较少、群体构建费时费力、无法覆盖多个不同品种等问题[61].近年来随着高通量基因型鉴定技术的发展,利用全基因组关联分析(genome-wide association study,GWAS)进行数量性状定位研究在植物学研究中受到重视[63,64].笔者所在的研究组通过对水稻自然变异群体的代谢组学分析,揭示了水稻籼粳亚种间代谢谱差异,获得了籼粳亚种分化的生物标志物(biomarker)[13].
进一步对代谢性状进行全基因组分析(metabolic genome-wide association study,mGWAS),获得了数百个控制代谢物自然变异的显着位点。大部分代谢物的自然变异受到多个位点控制,但相比复杂的农艺性状来说其遗传结构相对简单。
对籼粳亚种代谢组的遗传分析表明,控制两者的遗传结构存在明显差异,揭示了两个亚种代谢组差异的遗传基础[13].
2.3 用于代谢相关功能基因鉴定及途径解析
功能基因组学是后基因组时代一门重要学科,其主要任务就是鉴定每个基因的功能,其中就包括在基因组中占有相当大比例的代谢相关基因的功能。在过去10年中,通过代谢组学和其他组学,特别是转录组学的结合,大大加快了植物中代谢相关功能基因的鉴定。模式拟南芥拥有高质量的全基因组序列信息和非常完备的各类公共资源(包括cDNA全长序列、各种形式的突变体和丰富的转录组学数据信息等)及简单而高效的遗传转化方法。这些都为利用代谢组学和转录组学的整合分析进行代谢相关基因(特别是那些缺失或过量表达并不能引起可见表型变化的基因)功能鉴定奠定了良好的基础。通过对拟南芥代谢组、转录组的整合分析及后续的反向遗传学验证,成功鉴定了若干参与拟南芥类黄酮生物合成的基因。同样的策略也成功的被用于参与花青素(anthocyanin)[65]及原花青素(proanthocyanidin)[66]合成基因的鉴定。除此之外,该策略还被成功的用于胁迫条件下代谢相关基因的克隆和鉴定[67].代谢组与基因组整合进行基因功能鉴定主要是通过关联分析来实现,其原理是"共犯原则(guilt-by-association)",也就是说认为参与同一生物过程中的基因(或代谢物)通常是受同一个调控系统控制的[10].换言之,参与同一生物过程中的基因(或代谢物)有着相同或者相似的变化规律。这样,如果你所感兴趣的生物途径中某一步基因其功能已被鉴定,通过对不同组学数据的关联分析可以推测参与该生物途径的其他基因,并可以利用反向遗传学或生化手段进行功能验证。随着RNA测序(RNAseq)技术的发展,在非模式植物,如很多药用植物中通过代谢组与转录组数据关联分析也有不少成功的案例。Winer等[68]以罂粟(Papaversomniferum)为材料,通过对利用靶向代谢组学和RNA测序技术获得的转录组数据的关联分析,成功发现了一个参与吗啡生物合成的基因簇(由分属于5个酶家族的10个基因组成),并利用基因沉默技术鉴定了其中6个基因的功能。同样利用"共表达"(co-expression)的研究策略,Seki等[69]从甘草(Glycyrrhiza uralensis)中鉴定了参与甘草甜素(glycyrrhizin)合成的两个细胞色素P450基因。
另外,通过代谢组学与基因组学(第二代测序技术为代表的高通量测序)的结合进行代谢相关基因的克隆及鉴定则是该领域最新进展。笔者的研究组利用广泛靶向的代谢组学手段对524份不同水稻品种中超过840个代谢物(已知和未知)进行定量分析,研究了水稻代谢组的自然变异[13].在此基础上通过表型(代谢物含量)与基因型数据的全基因组关联分析,预测了36个代谢相关基因的功能,并对其中的5个进行了功能验证。在此基础上重构了水稻中包括类黄酮、酚胺等在内的代谢途径[13].与此类似,利用基于靶向代谢组学的代谢物全基因组关联分析也成功预测并通过转基因手段鉴定了水稻中参与酚胺合成的两个酰基转移酶编码基因[46].
3 展望
代谢组学是系统生物学不可或缺的重要组成部分。植物代谢组学通过近年来的发展,特别是与基因组学、转录组学等其他组学技术的结合,已经在植物生理学、遗传学、功能基因组学及相关生物技术研究领域发挥着越来越重要的作用,成为后基因组时代的研究热点。植物代谢物在种类及含量方面的极端多样性使得植物代谢组的研究显得尤为重要。作为一门新兴的学科,植物代谢组学还处于发展的初级阶段,无论是在方法学还是在应用研究方面都还存在着亟待解决的问题。未来主要的发展方向包括:1)开发基于现有独立分析平台的更为广谱、原位、即时、通用的检测方法;2)建立实现多种分析平台并行分析及数据整合的新型植物代谢组分析平台,真正实现对植物样本内代谢物的无偏性、高灵敏、高通量分析;3)开发准确、高效的植物代谢物鉴定或注释工具;4)进一步实现代谢组学与其他组学(基因组、转录组、蛋白组、表型组等)数据的整合,以便更好地阐述生物过程的分子机理,同时促进其生物技术应用。

随着新一代测序(Next generation sequencing)技术的迅猛发展,许多植物全基因组序列开始陆续测定完成,从大量的测序信息中大规模获取核基因序列,使得开发不同物种通用的核基因条形码成为可能。...

在植物水分生理研究中,植物水势可直接反映出植物水分亏缺程度、抗旱性强弱,因此它是公认的最常用的水分生理指标之一(付爱红等,2012).作为SPAC连续体中的中间环节,植物水势会同时受到大气水势、土壤水势的双重影响,同时,植物体内也会形成根-茎-叶...

在农作物种植过程中,因为长期使用化学制品导致昆虫的变异性逐渐增强,一种昆虫可能会有多种特征,对于各种药品经常产生抗药性,造成化学制品防治虫害的效果趋于减弱。...
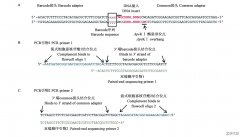
GBS技术现已应用在很多方面,对植物的准确鉴定、遗传多样性研究以及可持续发展都具有重要的意义。...

随着PCR技术的发展,其他技术与PCR技术的结合也逐渐增多,比如将毛细管电泳技术与多重PCR技术相结合,可以使基因分裂效率以及检测的准确度得到有效的提高。...
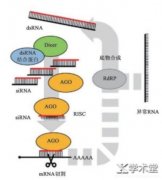
虽然RNAi技术在生物安全性方面的研究已经取得了一定的进展[72],但相关机制尚未明确,RNAi技术在作物改良中的应用潜力有待于进一步挖掘。...
菌根就是真菌加植物的根,它是指真菌与植物根系形成的互惠共生体。能够侵染植物根系形成菌根的真菌叫做菌根真菌。1989年,Harley根据参与共生的真菌和植物种类及它们形成共生体系的特点,将菌根分为7种类型,即丛枝菌根、外生菌根、内外菌根、浆果鹃类菌...
近年来,组学技术(基因组学、转录组学、蛋白质组学和代谢组学)和基于组学的数学模型在生命科学领域变得越来越流行,而基于组学分析的系统发酵优化和遗传改造策略在产油微生物中也得到广泛应用。...
微生物能够产生多种具有生物活性的次级代谢产物,其中大部分活性次级代谢产物来源于放线菌[1],这些物质具有抗菌、抗肿瘤、杀虫、免疫抑制和免疫激活等功效,被广泛地应用于医药业、农业、兽业、食品工业等领域。近年来,随着抗生素耐药菌的不断出现,深入研...

藻类蛋白质组学相关研究未来的挑战主要是进一步扩展对不同藻类系统基本组成部分的了解,而在基因组水平上,随着越来越多的藻类基因组计划的建立或完成,藻类蛋白质组学研究逐渐变得更为便利,针对蛋白质组学获得的数据分析更可信可靠。...