��������


����ȫ����ɺ�����ɺ���Į����ռ��½�������33%��40%������Щ��������ˮ�ֳ����������˴��ά��ֲ������棬Ȼ�������Ƥȴ�ܹ��㷺�ֲ���������һЩ����ռ�����︲�ǵ�70%���ϣ������Ƥ�������ࡢ���¡�̦�����������ϸ����������ֽ������������ڵر��γɵ�һ���װ���ĸ������������㣬�����ڻ�Į�����������ȶ���Ӫ�����ơ�ˮ��ѭ���ȷ������ŷdz���Ҫ�����ã�ͬʱ���ٽ������Ƥ����������Ҳ����Ϊ������ɳ�������ƻ�Į��������Ч�ķ���֮һ��������������Ҫ����̬�����Լ��Ի�Į�������صĹ��ף������Ƥ�ڽ�Щ���������˹������ڶ��о���Ա�Ĺ㷺��ע��
���������Ƥ��Ϊ��Į�������͵�����������ʽ������һֱ���ڲ��ϵķ���������̵��У��ڽ�Ƥ�ķ�����������У�������������IJ�ͬ��һ�㽫�����Ƥ����Ϊ���Ƥ�����½�Ƥ����Ƥ��3����Ҫ���������ͣ����Ƥ��������ȺΪ˿״���壬�����Ž�Ƥ�ķ��������¡�����Ϊ��Ƥ�����������Ⱥ����Ƥ�������������½�Ƥ����Ƥ�����Ž�Ƥ�ķ������棬��Ƥ������Ⱥ��ṹ���ϱ仯������ͬ��������Ⱥ�Ʊػ�Ӱ���Ƥ����̬���ܣ������о�֤ʵ����ͬ��������������Ƥ�ڹ�̼���̵�Ч�ʡ���ѹǿ�ȡ������ʵء�������϶�ȵȷ���������Բ��Ȼ��Ŀǰ���ڵ��½�Ƥ��Щ��̬���ܼ��������Է����������ڻ��Ʋ�û�н���������о��������ڽ�Ƥ�ķ�����������У�������ɼ��������仯������о��dz�ȱ��������չ�˷�����о����������ڴӱ����Ͻ�ʾ��ͬ��������������Ƥ����̬���ܡ��������Լ�������֮��IJ��죮
����1�������뷽��
����1.1�������ſ�����Ʒ�ɼ�
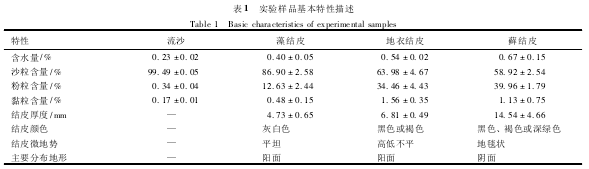
����������λ���ڸ���ɳĮ���ϱ�Ե��ɳ��ͷ����(37°32'N��105°02'E������1339m)�����ڰ����Ƹ�ԭ��Į���Į��ԭ���ɵش��������͵Ĵ�½�Լ���ɳĮ���õ�����ƽ������9.6�棬ƽ��������Ϊ180mm��������Ҫ������7��9��;����ƽ�����ʪ��Ϊ40%����ƽ����ɢ������3000mm����Ҫ��������������(5��9��)���õ�������ƽ������Ϊ2.6m·s��1������5m·s��1����ɳ��ÿ����200d���ң�������·�����ᆳ����Ϊȷ����·�������У���1956�꿪ʼ�õ��������˹�ֲ���ָ����������ڵ����棬�õ���Ŀǰ��̬�����õ��˸��ƣ������������Ƥ��ɳ�������γɣ�
����ʵ�����ý�Ƥ����ɳ��Ʒ��2010��6�²ɼ���ɳ��ͷ�������˹�ֲ���ָ�������Ʒ������������1����Ƥ����ɳ��Ʒ�û����������ɼ���װ�����������У�ParafilmĤ��ڣ�������Ʒ�����˻�ʵ���Һ�һ�����ڽ�����ص�ʵ�������
����1.2Ҷ����a�����IJⶨ
������Ƥ�еĹ����������Ҷ����a��ʾ��Ҷ����a�ⶨʱ����ͬһ��Ʒ��Ϊ���ݣ�һ��ֱ�Ӳⶨ��Ƥ���ܵ�Ҷ����a��������һ������ȥ������Ҷ״�����“��Ҷ”���֣�֮���ٲⶨ���е�Ҷ����a�������ⶨʱ����Ʒ�����в������ĥ��֮��ȡ1g������������10mL��ͪ��4����ȡ��ҹ��Ҷ����a��������Carcia-Pichel�ȵ���ɫ��ʽ���м��㣮
����1.3���������ֱ�ӹ۲����
������Ƥ�����������ȷ�����ý�Ƥ-ˮ���Һ��������ֱ�ӹ۲���������������Ƚ���Ƥ��Ʒ��150Ŀɸ��Ȼ��ȡ0.1g��ɸ��Ƥ��Ʒ����10mL����ˮ�����Ⱥ��ڰ�������12h��֮��ȡ0.1mL��Ƥ-ˮ���Һ�������¹۲죬�������������̬�������������ͬʱ����������������������㷽���������ķ�����
����1.4��������������ȷ��
��������Ƥ�������������¹�150Ŀɸ��Ȼ��ɸ��Ľ�Ƥ��Ʒ����ˮϡ�ͳɲ�ͬŨ���ݶȵĽ�Ƥ-ˮ���Һ�����Ⱥ��ڰ�������12h��֮����ÿһϡ��Ũ�ȵĽ�Ƥ-ˮ���Һ�зֱ�ȡ3��0.1mL��Ƥ-ˮ���ҺͿ����ţ��൰����������������������������I������������������������Ƥ�е�ϸ��������ͷ��߾�������ϸ����ƽ������37���������������߾��������ƽ������28������������3��5d����о��������
����1.5��Ƥ�������Բⶨ
������Ƥ��������������ˮ�����ʵ����(����ɳ���������������ֱ���ֱ�Ϊ��0.05mm��0.005��0.05mm����0.005mm)���л�̼���ܵ��������������������������ķ������вⶨ����Ƥ������α꿨�߽��вⶨ;��ƤpH�͵絼���ڽ�Ƥ5��1ˮ���Ƚ���Һ����YSI-Proplus�Ͷ����ˮ�ʷ����ǽ��вⶨ;��ƤK+��Na+��Ca2+��Mg2+��Mn2+������M3��������Һ����ԭ�����շֹ��ȷ����вⶨ��
����1.6ͳ�Ʒ���
�������о��в�ͬ����������Ƥ��Ҷ����a����������������������������������֮��ķ����������One-WayANOVA���з���;�������������������������������Ƥ��������֮�������Է�����BivariateCorrelations���з��������е����ݷ�����SPSS13.0�����Ͻ��У�
����2����������
����2.1�����Ƥ�ķ�������
�������Ƥ������Ϊ����Ⱥ�䣬������˿״���壬���������(Microcoleusvaginatus)��צ��α֦��(Scytonemajavanicum)�����Ƥ�ϱ���ƽ̹�����飬������ɫ��dz������Ƕ�һ�㣾80%�����½�Ƥ���Խ���״����Ϊ��������ͽ���(Collematenax)�����½�Ƥ������ӣ�����ϴֲڣ�������ɫ�������Ҷ״��ֲ��ڽ�Ƥ���棬�Ҵ�ʱ����ٳ��֣�����ʱ�������Ƕȿɴﵽ80%��ʪ��ʱ��������Ƥ���森��Ƥ�������ͶԳ�Ϊ��������Ҷ��(Bryumargenteum)���ڶԳ�(Didymodonnigrescens)�ȣ���Ƥ��ȿɴﵽ15mm�����汻��̺״����ֲ���������ǣ��Ƕ���ʱ�ɴﵽ100%��������ʾ�������������Ƥһ�㰴��“���Ƥ��→���½�Ƥ��→��Ƥ”��ģʽ���з������森��һЩ����£���Ҳ��ֱ�������Ƥ���ȷ��������Ӷ������Ƥֱ�����浽��Ƥ(ͼ1)��

����2.2��Ƥ�й�������������仯
������Ϊ��Ƥ����Ҫ��������Ⱥ����������ǽ�Ƥ�л�̼����Ҫ�����ߣ��ڽ�Ƥ�����������������桢�ָ������ж�������Ҫ���ã����о�ͬ�����֣����Ž�Ƥ�ķ������棬��Ƥ�еĹ���������������ӵ�����(ͼ2)�����ѵ��µ�Ҷ״������“��Ҷ”����ȥ����Ƥ�еĹ���������ʣ���������������࣮��ͼ2�����ʾ���������������������Ƥ�����½�Ƥ����Ƥ����������г��½������ƣ�Ȼ�����ܴ����Ƥ�����½�Ƥ���������������������������½������ܵ�����������ȴ�������ӣ���˵���������浽���½�Ƥ��ʱ�����������Թ���״̬���ڵģ�Ҷ״���еĹ������ǹ������������Ҫ�����ߣ�
������������(M��vaginatus)�����н�Ƥ�����ж���������������Ⱥ���еĵ�һ�����֣���������(�Խ�Ƥ�ƣ���ͬ)���ɴﵽ0.33mm3·g��1�������Ƥ����Ƥ����(P��0.05)��צ��α֦��(S��javanicum)�ڽ�Ƥ�е����������������壬Ϊ�ڶ������֣�α֦��(S��javanicum)�����Ƥ����Ƥ�е����������ϵͣ��ڵ��½�Ƥ�е��������ϸߣ����ɴﵽ0.13mm3·g��1��������(����һ��:Nostocsp��)�����Ƥ�����½�Ƥ�Լ���Ƥ�е����������ܵͣ�������������������0.02mm3·g��1�����ڵ��½�Ƥ��������(Nostocsp��)������ȴ���Ը�������������������(P��0.05)����߿ɴﵽ0.06mm3·g��1(ͼ3)��

����2.3��Ƥ�����������������仯
����3�������������ڲ�ͬ����ν�Ƥ�е�����
�������������Ƶı仯����(ͼ4)��������ɳ�����Ƥ��3�����������������������һ�����ӣ�֮���½�Ƥ�����ǵ��������ֳ����½������ƣ�������浽��Ƥ��3������������������ٴ����ӣ��������ǵ�������������Ƥ�δﵽ���ֵ��ϸ��(�Ծ���/��Ƥ�ƣ���ͬ)�ﵽ18×105CFU·g��1������ﵽ3.4×105CFU·g��1�����߾��ﵽ16.5×105CFU·g��1��
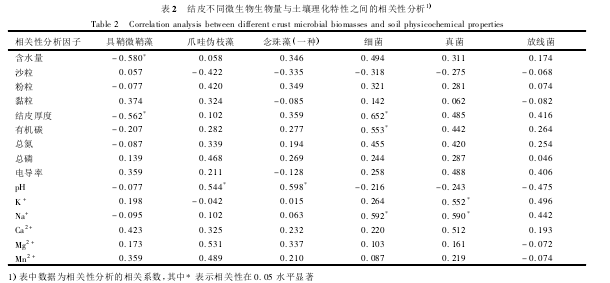
����2.4��Ƥ������������������������֮��Ĺ�ϵ
�������Ž�Ƥ�ķ������棬��Ƥ��ȡ���ˮ�������ӣ���Ƥ�ʵ���ɳ���������٣������������������(��1)��ͬʱ��Ƥ���л�̼���ܵ���������Ҳ�����ӣ����������ڽ�Ƥ��������(����δ��ʾ)��ͨ����Ƥ��������������������ָ����ط������֣�����(M��vaginatus)���������Ƥ��ȼ���ˮ�������������(P��0.05);α֦��(S��javanicum)��������(Nostocsp��)����������pH�����������(P��0.05);ϸ���������Ƥ��ȡ��л�̼��Na+���������������(P��0.05);���������K+��Na+���������������(P��0.05;��2)��
����3������
�����ڸɺ�����ɺ���Į����������ˮ�ֵ�ȱ����ֲ���������棬�������Ƥ��������ص�������̬����ȴ�����ڸõ����㷺�ֲ����������Ƥ��������Ĺ����У��ܶ��о���֤ʵ����(������˿״����)������Ի�Į������ǿ����Ӧ���ܹ�������ɳ��������ֳ�������������γ����Ƥ�����Ƥ������Ƥ��������ij����Σ�������ȴ�ܹ�ͨ�����Ʊ����������������ڽ�Ƥ������ֳ����Ŀ����ԣ��Ӷ��ٽ���Ƥ�ķ������森�ڱ�ʵ���з��֣�ɳ��ͷ�����������Ƥһ�㰴��“���Ƥ��→���½�Ƥ��→��Ƥ”��ģʽ�������棬����һЩ���������£���Ҳ��ֱ�������Ƥ���ȷ��������Ӷ������Ƥֱ�����浽��Ƥ��
������Ƥ�ķ��������ܺܶ���Χ������Ӱ��(�������ṹ�����ͣ�����ǿ�ȣ����ε�)����������һЩ��Դ����������(��ˮ�֡�Ӫ����)����ʹ��Ƥͣ����ijһ��������Σ���ɳ��ͷ��������ʵ���о����������������Ƥ�ڴ����ڵ����Ƥ�����ڵĵ��½�Ƥ����Ƥ�������棬�������Ž�Ƥ�ķ������棬�����ʵ����ƣ���Ƥ��������Ҳ��Ӧ�����仯(����ɫ���������Ӵֲ�)����Щ�仯һ��������˽�Ƥ�����������������һ����������������(������Ӫ�����������ʵ�)�ĸ��ƽ�һ���ٽ��˽�Ƥ�ķ������森�����������о�����������Ƥ�ڲ��������ڽη������棬��Ҳ���о�����һЩ�����������Կ�����ʹ���ڵĽ�Ƥ�������浽���ڽ�(����ţ����ڵĸɺ������µ�)����ˣ���ʹ��ͬһ�о��������������IJ����Կ��ܵ��²�ͬ�Ľ�Ƥ��������ģʽ���Ӷ���ͬһ������ֶ��ֲ�ͬ��������ε������Ƥ��
�������Ž�Ƥ�ķ��������������������ӣ���˽�Ƥ�������������Ϊ�����۽�Ƥ����ˮƽ��һ������ָ�꣮�ڽ�Ƥ��������У����ࡢ���¡����ڲ�ͬ�Ľ�Ƥ����γ�Ϊ��Ƥ��������Ⱥ����������ڵ����Ƥ�У����¡�����������ϵͣ���Ҫ�Ĺ������Ϊ��������(M��vaginatus)��צ��α֦��(S��javanicum);������Ƥ���浽����ʱ�����¡�����Ϊ��Ƥ��������Ⱥ������½�Ƥ�еļ��ͽ���(C��tenax)�Լ���Ƥ�е���Ҷ��(B��argenteum)�ͺڶԳ�(D��nigrescens)����ʱ�����»��������������������࣬ռ�ݽ�Ƥ�ֹ���������������������Ҷ״�����“��Ҷ”����ȥ��������������������ǽ�Ƥ�н��еĹ��������Ⱥ����ʵ���о����ֵ��½�Ƥ��(���¸Ƕ�Լ70%��80%)�����������������������������������30%;��Ƥ��(��Ƕ�100%)���������������������������10%��
��������������������γɵĹ����壬��˾��������������ԣ����ܱ��о������������������������������Ƥ�����½�Ƥ�����½������ܵ�����������ȴ�������ӣ���ͱ����ڵ��½�Ƥ�У����������Թ���״̬���ڵģ�����Ҷ״���еĹ������ǹ������������Ҫ�����ߣ�֮�����Ž�Ƥ��һ������Ƥ�������棬ʵ������ʾ��Ƥ�е��������������½������ƣ������������������ij��֣����������ռ���ռ�����£�Ȼ��������ij��֣���Ƥ��ȡ���ˮ����������(P��0.05)����˱�ʵ��������(M��vaginatus)��������ʾ���Ƥ��ȼ���ˮ���ʸ���ع�ϵ�����⣬��ʵ��������(M��vaginatus)���������Ž�Ƥ�ķ����������½�����Ҳ��������(M��vaginatus)���ᡢ�����������������������¡�����ȡ����α֦��(S��javanicum)��������(Nostocsp��)Ϊ�̵����࣬���������ڵ��½�Ƥ������������������������ڽ�Ƥ��������Ե�������������£�
�������Ž�Ƥ�ķ������棬�������������ƣ�������������������(���л��ʺ����������ʵص�)����Ӱ��������������������Ҫ���أ���ʵ���о�������������������(ϸ������������߾�������)���Ž�Ƥ�ķ������������Ӻ���٣�����ٴ����ӵ����ƣ������ڲ�ͬ����ν�Ƥ�еĹ�ϵΪ:��Ƥ�����Ƥ�����½�Ƥ�������Ƥ��ȣ����������������ڵ��½�Ƥ�������½�������������ڴ������µij������£���Ϊ���о����������д����ļٸ�(�������)�����ױ�����������Щ�ٸ�Ҳ�п���ͨ�����ڻ�ѧ�����������������������������ʵ����ϸ��������Լ����߾������������ܵ�����������������һ�µı仯���ɣ�����ϸ���������������л�̼��Na+�����������йأ����������������K+��Na+�����������й�(P��0.05)��
����4������
����(1)ͨ�����ڸ���ɳĮ����Եɳ��ͷֲ���ָ��������Ƥ�ĵ��鷢�֣��õ����������Ƥ����һ����ģʽ���з������棬�����ڵ����������Ƥ�����½�Ƥ����Ƥ3����Ҫ�Ľ�Ƥ��������Σ�
����(2)���Ž�Ƥ�ķ������棬��Ƥ������������������ӣ������Ƥ����Ϊ���½�Ƥʱ����Ƥ����Ȼ����������(���������й���������������)���ӣ�����������������(�����������й���������)������ȴ�½�����ʱ�����ദ�ڹ���״̬;����������ֺ����������ռ䱻ռ�ݣ������Ƥ��������������һ���½���
����(3)���Ž�Ƥ�ķ������棬�������������ƣ�Ϊ��������������ṩ��������������Ƥ�в�ͬ������Ⱥ�������ı仯һ�����Ƕ����������仯����Ӧ;��һ����Ҳ�����˽�Ƥ��ͬ�����������̬���ܵ�������Խ�Ƥ�ڻ�Į��������Ӧ������������Ҫ�����壮
����

Ԥ���Ϳ���ҽԺ��Ⱦ���ѳ�ΪҽԺ��������Ҫ����֮һ���Ի�������������Ϊ���ݣ����ƶ�Ԥ��������ҽԺ��Ⱦ����Ժ����������2011-01-2013-12�����ڼ��ҽԺ���������걾��1036�ݣ�ͨ�����Ѽ�������ѧ���Ϲ���������ȫԺ������Ա���ҽԺ��Ⱦ��...
�����������T-RFLP��������������
������ĩ��Ƭ�γ��ȶ�̬�Է���������TerminalRestrictionFragmentLengthPolymorphism,T-RFLP�����и�ͨ�����ͳɱ������Ͷ������ص㣬��������̬ѧ����̽��Ⱥ��ṹ�����ܼ��䶯̬�仯�е�һ��ʵ�÷���Ĺ��ߡ�������ѧ����Ϊ���������һƪ��ĿΪ����...

���������ٶȲ�����ø���Ǻ��������߷�����
���������������������ѧ���������ʵ���ѧ�е���Ҫ���ݣ�����ʵ��Ӧ����Ҳ�dz��㷺������ҩ����ļ���������ҩ���ɸѡ�ȡ�...
�Ե�����˹�ؾ���5������������п�¡������
����ϸ����������˹�ؾ���Listeriamonocytogenes��LM�����ǽ�20��������������ʶ��һ���������������²���������4���¿�������ֳ����ȾʳƷ�̶���Ⱦ���࣬��Ⱦ��������ԼΪ25%,WHO������Ϊ20����90���ʳƷ���Ĵ��²���֮һ��Ҳ����Ϊ...
������ʩ�ʶ�����������̼����Ӱ�����
�����������������л��ʵĻ��Բ��֣������������Ծ�ijɷ֣���Ȼֻռ�����л��ʵ�3%����[1],�������ֵĹ�Ӧ��ת����ѭ��������Ҫ�����ã�������仯�ɷ�ӳ���������ƶȺ����������ı仯�Լ���������Ⱦ�̶�[2-4].������ʩ��Ӱ������ͨ����ˮ���¶ȵ�����...
������40����²���ά��ø���IJ�ø��������
������ά���ǵ�������ḻ���л���Դ֮һ����ά��ø���ܹ�Эͬ������ά��Ϊ�����ǵ�һ��ø���ܳƣ������������Ͼ���ø��Ҳ��Cxø���������Ͼ���ø���Ͼ�����ø�ȡ���ά����ԭ��ͬ���ǻ����ͣ�SSF�������Ҵ�������WilliamFreder-ickGauss������ù���...

�����������о������ߵ���ø��Ĺ��ܷ���
���ߵ���ø��(immunoproG teasome,IP)������һ������ĵ���ø��,������������������,���ܹ�����ϸ������������ϸ�������Լ���������Ӧ������Ҫ��֯�����Ը�����I��(MHCGI)��ԭ��ʡ�Tϸ����ֳ��....

������Ϣѧ�ӽǷ�������ѿ�߸˾�������ø
����ѿ�߸˾���һ�ָ�����������������ϸ��, �ɺϳɿ������������谭�²�����ֳ�����������������������Ǿۺϳɵĸ߷��Ӷ���, ����������Ϻ��, з�Ǻ�����֫������ǡ��͵�ֲ���С�...

���﹤��Ҳ����֮Ϊ�����̣��������ִ����\���ֶ���������������Ҫ�IJ�Ʒ��������и�ҵ��չ����ǰ��ҽҩ��ҵ����ʳƷ���������������������﹤�̾�ʵ���˹㷺Ӧ�ã������Ų�����������á�...

���ij�����ȺӰ�����ء���Ҫ���ܼ����÷���
������Ⱥ��������������л��干���Լ��²����������Ⱥ�䡣������Ⱥ��������Լ��������ϸ��������ϸ����������������Ⱥֱ�ӻ��ӵز���ߵȶ�������������������е������Ͳ������̡�...