�������ᣨ�ԣ�������ͣ������棩��ʳ��Ŀ���ã��������ᣩȮ�ƣ��ã�����壩����ԭ������ظ�ԭ�����Ĺ���Ȯ�֣�Ҳ���й�Ψһ��һ������Ȯ�֣����ڹ��Ҷ�������������ڽ���ʮ�������ᱻ�������˵��ڵ�ʡ�ݣ��������⣬���ϵ������滷�����ຣ�����ص�ԭ�������������������������������Ⱥ��Դ��ʧ��Σ�գ����ܵȣ�����������������Լ������ǰ�ſ�ʼ�˶Բ������ʶ���о�����Ҫ�о������ڲ��ἲ�����Ρ�������������ֳ������Ʒ�ּ���ȷ��棬�����о��漰����ѪҺ����������������ȣ���������������Ⱥ�Ŵ������ԣ��̣�ȣ�������������ѡ��ѡ������̩���ȣ���������������Һ���棨֣���ȣ������������ȷ��棬��ϸ�������ӷ�����о����١��������о�����Ӧ�ö�����ϸ����¡����������¡����óɹ����ʣ���ȣ�������������������δ��չ�����о��ı�������ϸ����¡�����ѳ�Ϊ������ϡ������Դ���·�����Ŀǰ�ڹ����ѽ����˴���è�������ȣ����������������䣨����˳�ȣ����������������q��������ȣ���������������ϡ����Ƥ������άϸ��ϵ����ϸ������ѧ���Ժ��Ŵ���Դ�����о������о�������֯�鷨�����ˣ������Ƥ������άϸ��ϵ���������������������������������ԡ��䶳���ա����ε��ױ��ϸ�����ͽ������о���Ϊ������ϸ����¡�о��ṩ�˹���ϸ����
�����������뷽��
������������ �� �� �� �� �У£ӡ��ģͣţ͡��ģ��ƣ��������ԣãͣ��������ȵ���ø���Ҷ�����������ƻ����Һ��̥ţѪ�塢�ţǣơ���ƣǣƾ����ԣǣ���﹫˾�������������ԣ���������﹫˾���ǣ������Ⱦ�ϡ�̨����Ⱦ�Ͼ������Ϻ��������﹤�̼�����������˾��
����̨ʽ���Ļ����ԣţ�������湫˾��������̼�����乺������˾����������̨���Զ���������˾������ӫ�������������˾��
����������Ƥ����֯�Ļ�ȡ�봦�� ��ֻ��������ᣨ������ĸ����������ë����ϴ��������ȡƤ����֯Լ��������������������Ỻ��Һ������������塡���������������壬�У£ӣ���Һ���� ���£������˻�ʵ���ҡ��ãУ£���Һ������ϴ��֯�飸�������Σ���������Ƭ��ȥ��֯����ë�������ֱ�Ƥ�����ۿƼ���ȥ�����ǣ���Ƥ����֯����Լ��������С��顣��֯�龭�У£���Һ����ϴ�ӣ��Σ���ϸ������Һ���ģ��ƣ��������ӣ�����̥ţѪ�塢�������ɣգ�������ù�ء�������μ�磯������ù�ء�����μ�磯���̱�Ƥ�������ӣ�����ϴ�ӣ��Σ��ã�����ϸ������Һ����������
����������ϸ������ Ϊ��������֯����������������̥ţѪ��Ϳ���ԣ���ϸ������ƿ���ã�����磩�ײ���Ȼ�������ܽ���֯��ת����ϸ������ƿ��������̽�뽫��֯��ڷ���ƿ�ײ���ÿƿ�������顣������ƿ���÷���ãϣ���������������������룳����ϸ������Һ��ת����ƿ����������ÿ�����������ϸ������Һ���۲�ϸ�������������ϸ������֯����Χ�������������Χ��Ƭ����ʱ���д������ãУ£���Һϴ�ӣ��Σ��� �� �� ̽ �� �� ȡ �� ֯ �飬�� �룲 ���̣����������ȵ���ø�ͣ����������Ҷ�����������ƻ����Һ�ڣ���������ϸ�����������룲����ϸ������Һ��ֹ�������ڣ����������������ϸ���������壬��ϸ������Һ��������ϸ�����������ԣ�×�����������̵��ܶȽ�����������ƿ����������������������������ϸ���ﵽ����������������ϣ��ٴΰ����ã��ã��������д���������
����������ϸ�������븴�� �����е�ϸ���ﵽ�����������������ʱ���Խ���ϸ�����棬���У£�ϴ�ӣ���ø���������ĺ���ÿһ���ԣ�������ƿϸ���������мӣ�����Ԥ������ ���ϸ������Һ ���ģ��ƣ��������ӣ�����̥ţѪ�塢�������������������Ƶ�ϸ������ܣ��ã�����磩������ϸ������У��Σ�����壩�ڣ� ��ƽ�⣰�������ת�Ƶ�����������±��䱣�棲�����ϸ�������Ͷ��Һ�����档����ϸ��ʱ����������ڣ�����ˮԡ�п���ҡ���ⶳϸ�����Ƶ������������Ĺܣ����룹����ϸ������Һ���ȣ����ij���ϸ�������룵����ϸ������Һ��������ϸ�����Ƶ��ԣ���ϸ������ ƿ �� ����������� �� �� �� Һ ȥ �� Ư �� �� �� ��ϸ����
����������ϸ������������ ȡ��ƿ��������������ϵģƣ���ϸ������ø���ţģԣ����Һ��������ϸ������Һ��ϸ��ϡ������×�����������̡���×�����������̺ͣ�×�����������̡����� �� �ȣ��� �� �� �룲���� ϸ �� �� �� �壨�ΣգΣã����У�ÿ�ף����̣��ڣ��мӣ��ף����ӣ����С�������������������У�ÿ���������������Һ���ӵڣ��п�ʼÿ���ڹ̶�ʱ��ȡ������������������ϸ����ϡ�ͺ���Ѫ����������ϸ��������ȡ��ƽ��ֵ����������Ϊ�����꣬��ϸ������Ϊ�����꣬����ϸ���������ߡ��������¹�ʽ���ϸ��Ⱥ�屶��ʱ�䣨���������������������硡������壬�Уģԣ���
����
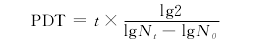
����ʽ�У�����������ʱ�䣻�Σ������״μ�����õ�ϸ�������Σ�����������ʱ����ϸ��������
����������ϸ�����ε�������ӫ����� �ο��£������飭���ȣ����������������������ġ���ϸ����Һ��ÿ�ף����̵�������Ԥ�ȷ��øDz�Ƭ���ΣգΣã��ģ��װ��ڣ�������ϸ��������������������ϡ�������������ÿ���뺬������ȩ�ģУ£ӣ����·�����������������ȩ���У£�ϴ�ӣ��Ρ��ڸDz�Ƭ�ϣ��ӣ����̺��������ƣ£ӵģУ£ӣ����ã��������������ú��������������������ԣ������أ������ͣ����� �ƣ£ӵģУ£�ϡ��һ���������������졡�����飭��������������������������������䡡���������壬�����磯���̣��ӣ����ᣭ���̣ģңɣãȡ��ɣ�㣮���ڣ������������Ĺ��У�ϡ�ͱ���Ϊ���ã����������ø������Ļ�����ٶ����Ŀ���ϡ��Һ�Ͷ�������Һ���ӳ�����������������Һ����һ������μ�̣�������ȡ���Dz�Ƭ���ٽ������ڣ���μ�̵�һ����Һ�ڣ�ϸ�������¡�������ʪ���ڷ������衣ȡ���Dz�Ƭ��ϸ���泯�ϣ����뺬������ �ƣ£ӵģУ£ӣ����ã�������������Һ���ظ��������ϡ��ú������������������� �ԣ������أ������ͣ������ƣ£ӵģУ£���Һϡ�Ͷ������������������ƣɣԣã���������������䡡�����������塡������������飭�������塡�ɣ�ǣ��ȣ��̣������������磯���̣��ӣ����ᣭ���̣ģңɣãȡ��ɣ�㣮����ϡ�ͱ���Ϊ���ã����������Ⱥ����ij�ȥ�������ڸDz�Ƭ�ϼ�һ�ζ�������μ�̣����Dz�Ƭ���ڶ�����Һ�ڣ�����������ǣ��ܹ��ţ����·������衣ȡ���Dz�Ƭ��ϸ���泯�ϣ����������壬���뺬������ �ƣ£ӵģУ£ӣ����ã�������������Һ���ظ��������ϡ��μӺ��������� �ԣ������أ������ͣ���μ�磯���̡��Уɣ��������������������壩�ģУ£ӷ����������ãУ£�ϴ�ӣ��Ρ��ÿ�ӫ������Ƭ����������������������ǣԣͣ��ӣ���������£������裬�£������磭�������գӣ�����Ƭ����ӫ�������¹۲졣�ƣɣԣ��ã�������������⣬�У��ã�������������⡣ͨ���������Ƚϣ�ȷ��������Ⱦɫ����
����������ϸ�����ͷ��� �ο��ƣ��������ȣ����������������������ġ��������̡��ƣ���ϸ����Һ�ԣ�×�������������ܶȽ����ڣԣ�������ƿ�������������������ˮ����ʹ����Ũ��Ϊ������μ�磯���̣����ã��衣����ø����ϸ�������ĺ��룰������������죯�̡��ˣãɻ���ϸ�������� �澲�ã��������������Ĵ���״�����Ϻ����ģ������壬���������״������Ͼ��ã�����������ϸ���������壬���룰���������������̴���� ������ �� �� �� ϸ ������ �� �� �� ϸ �� �� Һ �ӣ���������ε����IJ�Ƭ�ϣ���б��Ƭ��Һ����չ����������ﲣƬ������������۲죬��ϸ��������չ����������IJ�Ƭ���ڲ�Ƭ�ϼ����δ��ǣ������ȾҺ��Ⱦɫ��������ˮ��ϴ������ڲ�Ƭ�ϵμӣ��ζ��ױ������ϸDz�Ƭ�����;��¹۲��ɢ���õ�Ⱦɫ�岢���ա��ãУ����������������е���Ⱦɫ�壬�ɶ����к�����������Σ����ţ�����塡�ԣ飩ͼ�����������Σɣӣ��ţ��������£ҡ������������ӣУ�������Ⱦɫ�����ݲ�����
��������������
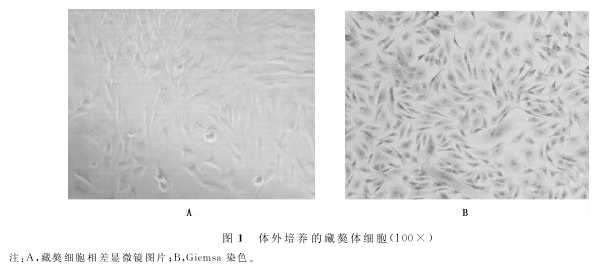
��������������Ƥ������άϸ���������������������̬���� ����֯�鷨�����ˣ������Ƥ������άϸ��ϵ����֯�龭���������������䣬��Χ�ɼ����λ���·ʯ�������ϸ���γ�����������������������ʱ���ӳ��������������䣬��֯����Χ���ִ�Ƭϸ�����������������֯����Χ��ϸ��������������ϸ��Ϊ��������ϸ����������������ϸ���Ͷ����ϸ�������������֯������������������ʱ���д���������������ϸ������Ѹ�٣�����դ��״������״�������ڣ���������������������ϣ�ͼ���������ٴν��д������������������IJ���Ƥ������άϸ��������պ�̨ ����Ⱦ���� ������ⶨ��ϸ������ʴ������������������������̬�ʹ���ʱ��δ�����Ա仯�����������IJ���Ƥ������άϸ���������δ���������ϸ�������ٶ����Լ����������д���ϸ���ѱڡ�
����
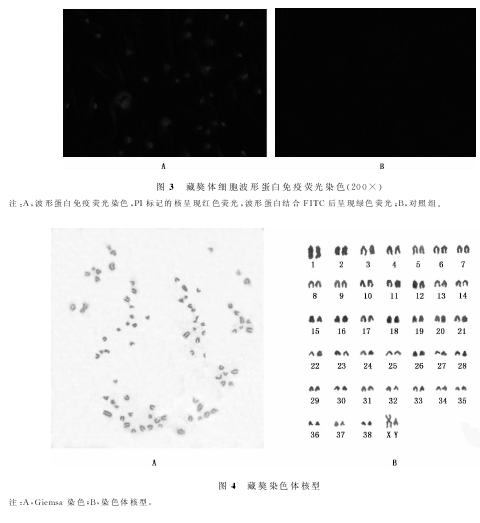
����������ϸ�����������ص� �ƣ�������Ƥ������άϸ����������ͼ������ͼ����֪����������ϸ�����������DZ���ں�ʼ������ֳ�����������ԵĶ��������ڣ��������䣩��Լ�ڵڣ���ﵽƽ̨�ڣ�ƽ̨���Ժ�����˥���ڡ����Խϵ�ϸ���ܶȣ���×�����������̣���������ʱ��DZ�����ӳ����������䣩���������������߽�ƽ����Լ�ڵڣ���ﵽƽ̨�ڡ���������ϸ���ﵽƽ̨��ʱ��ϸ���ܶȷֱ�Ϊ������×�����������̡�������×�����������̡�������×�����������̡��������õģУģԷֱ�Ϊ�����������������������������裬���ڲ��鶯����ϸ�������УģԷ�Χ��
����

����������������ϸ�����ε�������ӫ�������� �ò��ε�����¡�����������ӫ��Ⱦɫ���������ȳ������Ե����Խ��������ɫӫ���ǵIJ��ε�����Ҫ�ֲ���ϸ������Χ����ͼ������
����������ϸ�����ͷ��� �ƣ���������ϸ��������Ⱦɫ����ͱ���Ϊ��������������ͼ����֪������Ⱦɫ�売�������ȫ����Ⱦɫ���Ϊ����˿��Ⱦɫ�壬��Ⱦɫ��ϴ�Ϊ������˿��Ⱦɫ�壬��Ⱦɫ��Ϊ����˿��Ⱦɫ�塣
����������
����Ŀǰ���������б����ɹ��ؿ�¡�˹����ʣ���ȣ������������ͻ��ǣ��ˣ���ȣ���������������������δ��չ�ⷽ����о������б��������˵¹�����Ȯ����С�ʵȣ��������������ȸ�Ȯ����Ȫ�ȣ�������������Ȯ�����άϸ����������������ϸ��������ر������١����о�����֯���������ɹ������ˣ���������άϸ�����⣲��ϸ���������á���ֳ����ǿ��������������䶳�����˸���ϸ��������ܣ������̶���ܣ�����ϸ��ˮƽ�����˲����Ŵ���Դ��ҲΪ���ڵIJ�����ϸ������ֲ�ṩ�˹���ϸ����
������֯�������dz��õĶ�����ϸ��ԭ���������������ڱ����˳�ʱ��ø������ϸ��������ƻ����ã�������֯���������ܽϺõر�����Դ��֯������������ϸ������ϸ������ֲ������Ч������ҪӰ�죨�У�������ȣ����������������о�����֯�鷨�����IJ������άϸ�������˽Ϻõ�ϸ����������������߲�����ϸ������ֲ����Ч�ʡ����������IJ������άϸ���������Լ���̬�ص����ѱ�����Ȯ�����άϸ�������ƣ���С�ʵȣ����������������о����ö������Ƥ��ϸ����ø������ʱ�ϳ��ʹ��������ڽ������ص㣬ͨ��������ø����ʱ�䣬�ڴ���������������ƿ��ȥ����Һ������������Һ�����������δ��������õ��˽ϴ������γ���άϸ��Ⱥ�塣
����������������Ķ�����ϸ���ܵ�������������������������������ص�Ӱ�죬����Ⱥ��Ⱦɫ��������Եĸı�����ֳ�����仯�ϴⶨϸ�����������ܽϺõط�ӳ��ϸ������ֳ����������Ҳ������ϸ���Ľ���״�������о��ⶨ�˲������άϸ���������ߣ�ƽ̨���ܶ����ֵ�ﵽ������×�����������̣���̣Уģ�Ϊ���������裬����ϸ����ֳ����ǿ�������ٶȽϿ졣���ε��ף��������������ϸ����һ����Ҫ��ϸ���Ǽܳɷ֣���Ƥ����֯�в��ε����ڳ���άϸ�������������Ϊ���������ij���άϸ��������־֮һ���أ��ȣ����������������������IJ������άϸ�������˲��ε��ױ����о���֤ʵ����ϸ�����ֲ��ε��ױ������ԣ����ϸ����̬�����ɳ����ж�ϸ��Ⱥ�����ڳ���άϸ�����͡�
�������������Ķ�����ϸ���������֯�������ͬ�����Ŵ�������������������Ⱦɫ����졣��ϸ��Ⱦɫ���������Ҳ��ϵ����ϸ����¡��̥�Ͷ���ķ����������ʣ���ȣ��������������Բ������άϸ��������Ⱦɫ���������о�������Ⱦɫ��������״�����ж����ܷ���Ϊ������ϸ������ֲ�о��Ĺ���ϸ����ͬʱ������Ⱦɫ������о����ຣ��������ķ��ࡢ�������Ŵ���������Ҫ�ο���ֵ����ǰ���о�����������Ѫϸ��Ϊ���Ͻ��в���Ⱦɫ����ͷ�����Ϊ�˽�һ���о��ຣ����������������ϸ����Ⱦɫ������������������������ģƣ���������ϸ��������ϸ���Ŵ�ѧ��������ͳ��������ʾ���ƣ���������ϸ��������Ⱦɫ����ͱ���Ϊ������������������һ�������ķ�������Ⱦɫ�����ͺ��ı��壬�б�Ҫ���������������Ż�����ֹ�����Ŵ����������Ӷ����������ص�Ⱦɫ����졣Ⱦɫ����ͷ������֤ʵ����Ⱦɫ�売�������ȫ����Ⱦɫ���Ϊ����˿��Ⱦɫ�壬��Ⱦɫ��ϴ�Ϊ������˿��Ⱦɫ�壬��Ⱦɫ��Ϊ����˿��Ⱦɫ�壬���ѱ�����������Ѫϸ������ ��Ⱦɫ��� �ͷ������ �� һ �� ���� �� �ȣ��������������Ǿ��ȣ�����������������ͼ����������������Գ�Ⱦɫ�������������ʾ�˲���Ⱦɫ�����͵Ļ���������Ȼ��������Ⱦɫ��걾�д�����һЩ���۲��СȾɫ�壬��Ŀǰ�ķ�������ȷ��Դ�����һ�������ѣ�����Ӧ��Ⱦɫ���Դ���ӫ����Ⱦɫ��ͷ���̽�뼼�����н�һ���о���
�������о������˲������άϸ���꣬�䶳������Լ������ܣ������̶���ܣ�����ϸ�����������������������������ߡ����ε��ױ��Ⱦɫ����ͽ������о�������Ƥ������άϸ��ϵ�Ľ��������Ա����������������Դ��Ϊ�Ժ�չ�����������ѧ��ϸ������ѧ�о��ṩ�����õIJ��ϣ�ҲΪ���IJ�����ϸ����¡�о��춨�˻��������������IJ������άϸ����Ϊ����ϸ�����òع�ȥ����ϸ����Ϊ������ʽ��в�����ϸ������ֲ�о������������IJ����¡��̥��ֲ���ع��������ѹ������������¡�����������߲��ֳᷱЧ�ʣ������������������Ⱥ����Ҳ�ǹ��ڼ̲��ᾫҺ���漼��Ӧ���ڲ��ֳ֮ᷱ��������һ����Ҫ�IJ��Ḩ����ֳ��������������������ƶ����ֳᷱ���\���ķ�չ��Ӧ�á�ͬʱ�����о��������IJ�����ϸ������������ϵ��ΪȮ����ϸ�������ͽ�ϵ�ṩ�ο���
�� �� �� ��
������˳�����£������ᣬ�ȣ����������άϸ�������������ع��߹����ۣʣݣ�ũҵ���\��ѧ������������������������������������������
����������������ţ���ΰ�ã��ȣ����qϸ��ϵ�Ľ�����������ѧ���Թ۲�ۣʣݣ�����ѧ�о�����������������������������������������
��������С�ʣ����������𣮵¹�����Ȯ����άϸ�������������������о��ۣʣݣ��й�������ҽ����������������������������������������
���������Ǿ����Ż�Ƽ������Ⱦɫ����ͷ����ۣʣݣ��ຣ������ҽ��־������������������������������������
���������������أ�֣���࣬�ȣ�����èƤ������άϸ���ڲ�ͬ����Һ�е�����ѧ�����о��ۣʣݣ����ݴ�ѧѧ����ũҵ��������ѧ�棩������������������������������������
���������ţ�Ԭ���������࣬�ȣ����ᣨ�ԣ�������ͣ������棩�ĺ����о��ۣʣݣ��ຣҽѧԺѧ������������������������������������������
�������������ã�������ã��ȣ��������Ѫ�嵰����ָ����о��ۣʣݣ���������ҽ������������������������������������
������֣��壬��������ʺ죬�ȣ�����Ũ�Ⱥ�ƽ��ʱ��Բ��ᾫҺ�䶳Ч����Ӱ��ۣʣݣ�����ũҵ��ѧ������������������������������������
��������Ȫ���Ƹ��������ң���������Һ�ԣ£�����Ȯ��������άϸ������ѧ���Ե�Ӱ��ۣʣݣ�ҽѧ�о���־������������������������������������
�����������ܣ���̩��������Ʒ����Դ�����뿪�����ãۣʣݣ�������̬ѧ������������������������������������������
����������̩�������ܣ�۳������������ȮƷ�ֱ�����ѡ���о��ۣʣݣ�������̬ѧ��������������������������������������
���������£���������ʡ��ӣ��ģ����ͣ��ȣ�����䡡�ʡ��£��������죮�¾�������誣������ܣ����룮����ϸ������ѧʵ��ָ�ϣۣͣݣ���������ѧ�����磬����������
���������ƣ����������ҡ��ɣ��¾��������˨���룮����ϸ������———��������ָ�ϣۣͣݣ���������ѧ�����磬����������
���������ʣ��硡�ǣ��У��롡�š��ӣ��ã��ʡ��ˣ��������죮�У������������������졡����������壭�����������������䡡���������塡��桡������������塡������������������塡�����������㡡����졡���������������������������������������䡡������衡����磭������������죭�������䡡����������ۣʣݣ��ԣ�����������������������������������������������
���������ʣ��硡�ǣ��ȣ��硡�ӡ��ǣ��ϣ衡�ȡ��ʣ��������죮����������䡡�������������塡�������������������������㡡������������䡡�������������䡡�������塡����ۣʣݣ��ԣ�����磭������������������������������������������
�������� �ˣ�����͡��ˣ��ʣ��硡�ǣ��ϣ衡�ȡ��ʣ��������죮�ţ��������䡡��������������������������������������㡡������ۣʣݣ��ã�����硡�ӣ�������ã��������������������������������
���������̣顡�ѡ��ƣ��̣�����ڡ��ӣ��̣顡�١��أ��������죮�ϣ�������䡡��������������㡡�����������桡�ԣ�������ͣ������桡�����䡡������塡���������������졡�ģΣ�������������ۣʣݣ��ʡ��ǣ�������ǣ��������������������������������������
���������У������䡡�ң����죭�ң����������ƣ��£����ƣ��������죮�ģ������졡��������������������������������塡����������塡��桡���������㡡�������������������������塡��桡���������ۣʣݣ��ʣ������졡��桡�����������������䡡������������������������������������������������������
�������� �أ����ȡ��أ��٣顡�٣��̣顡�̣��������죮�գ���������������£���������䡡���������������������������������������������������������桡�������壭�����䣭�����������������䡡�������������桡������������ۣʣݣ��У��������������졡�У����������������졡�У裬����������������������������4.