园艺学论文


杏属(Armeniaca vulgaris Lam.)共有 10 个种,中国有 9 个种,13 个变种,2 000 余个品种和类型(赵锋 等,2005)。‘大果’杏源产于山东省德州市,属于华北生态群品种,树势强健,树姿半开张,不完全花比率低,成花容易,坐果率高,结果较早,果实品质优良。该品种早熟丰产,耐瘠薄,耐干旱,设施栽培成功率较高(王金政 等,2005),其栽培管理较其他品种简单,且用途极为广泛,现已在山东省及黄河流域其它地区广泛种植,并成为该地区经济效益较高的杏树品种(魏敏,2003)。杏在荒山绿化、退耕还林和果树种植调整中发挥着重要作用(陈学森 等,2001)。杏树开花早,易遭早春晚霜危害,轻则造成减产,重者甚至绝收(孙仲序 等,2005)。据统计,2001 年山东邹平因低温冻害,全县杏经济损失 8 000 万元;同年山东泰安‘大果’和‘凯特’杏受冻均在 90%以上。因此解决杏树早春晚霜冻害具有重要的意义。
植物的抗寒性是其对低温寒冷环境的长期适应,由遗传变异和自然选择获得的一种能力(Glerum,1985),是对低温长期适应的一种遗传特性。由于木本植物的多年生和木质化构造的特性,对低温比草本植物有更高的抵抗能力,因此,从木本植物中应该更容易分离到有较强抗寒能力的基因(Welling et al.,2002),这也是改良林果树种的一个重要方向。
植物 EBP1 与人体内 EBP1 的结构和功能相似性,是近年发现的一种新型蛋白质,被认为是重要的转录因子和转录共调节因(Yoo et al.,2000)。EBP1 是增殖相关蛋白 2G4(PAZG4s)家族的一员,真核细胞中 PZG4 蛋白高度保守(Zhang et al.,2003),参与多个信号传导通路,与细胞周期、蛋白质的转录翻译相关,影响细胞的增殖和分化(Zhang et al.,2003;Horvath et al.,2006)。AmEBP1是从沙冬青(Ammopiptanthus mongolicus Cheng f.)中克隆(Cao et al.,2009)的与低温胁迫相关的基因,该基因是第 1 个被证实能提高转基因植物抗寒性的核糖体相关蛋白基因,通过促进核糖体的合成以及冷诱导中转录因子和下游起保护作用的蛋白的翻译而起作用。所以 AmEBP1 既对低温信号的转导过程有重要作用,也在植物对低温的长期适应中发挥重要作用。花粉管通道法转化外源基因,操作简便,已在番茄、番木瓜、苹果、核桃等果树上取得成功(牛庆霖 等,2013)。本研究中将沙冬青 AmEBP1 通过花粉管通道法导入‘大果’杏幼胚中,经抗性筛选及分子检测,在改良 WPM 培养基上培养阳性植株,探讨转基因植株的抗寒性,为下一步抗寒杏新品种选育提供参考。
1、 材料与方法
1.1 试验材料
将沙冬青 AmEBP1 完整的开放阅读框用 PCR 方法克隆出来,插入 PBS-T 克隆载体,PCR 方法鉴定与 LacZ 编码方向一致的中间载体,用 XbaⅠ和 SalⅠ酶切,同时双酶切载体 pCAMBIA2300(中国科学院遗传与发育生物学研究所惠赠),用 T4DNA 连接酶,以 6︰1 浓度比,16 ℃链接 12 h,获得表达载体 pCAMBIA2300-AmEBP1,冻融法转化到大肠杆菌 DH5α 中,卡那霉素筛选阳性菌株,4 ℃下保存备用。授粉试验于泰安市泰山林业科学研究院试验基地进行,采集‘凯特’杏花粉 4 ℃保存。
1.2 花粉管导入外源 AmEBP1
采用碱裂解法提取目的 AmEBP1 菌液(Lu et al.,2001)。经纯化后的质粒 DNA 溶于 pH 7.6 的TE 缓冲液中,A260/A280比值在 1.9 左右。
于 2012 年 4 月初‘大果’杏花大蕾期人工去除个别花蕾未膨大和未开放的花,对开放的花去雄授‘凯特’杏花粉,3 h 后取 5 μL AmEBP1 DNA 溶液涂抹柱头 2 ~ 3 次(Tang et al.,2005)。以未转化的花为对照。于盛花期后 30 d,取幼胚,用 70%酒精、0.1%升汞灭菌,无菌水冲洗 3 ~ 4 次,剥去种皮,接种在改良 WPM 培养基(附加 Kan 50 mg · L-1+ 6-BA 0.8 mg · L-1)上培养,筛选出的芽团用作分子检测。阳性材料接种到改良培养基(WPM + IBA0.2 mg · L-1)上生根成苗培养用于抗寒试验。
1.3 转基因植株的 PCR 检测
按 CTAB 法提取转化植株和对照植株基因组 DNA,AmEBP1 质粒 DNA 作为阳性对照:正向引物:5'-ATGTCGGATGATGAAAGGGAGGAGAAGGA-3';反向引物:5'-TAGGGAAGGGAGGTCAATCTTGAGGTGT-3'。反应体系:反应总体积为 25 μL,包括 2.5 μL 10× Buffer、0.4 mmol · L-1dNTP、2 mmol · L-1MgCl2,1 U Taq DNA polymerase、1 μL 正向引物、1 μL 反向引物、1 μL 模板 DNA(DNA 浓度 10ng · μL-1),ddH2O 加至终体积 25 μL。反应程序:94 ℃,5 min 预变性;94 ℃ 变性 40 s,55 ℃扩增40 s,72 ℃延伸 1 min,35 个循环;最后在 72 ℃延伸 5 min。扩增产物用 1%的琼脂糖凝胶电泳检测。
1.4 转基因植株的 Southern 检测
CTAB 法提取 PCR 阳性的转基因植株和对照植株 DNA,由北京美莱博医学科技有限公司进行PCR 标记法 Southern 检测(牛庆霖 等,2013)。具体方法:XbaⅠ和 SalⅠ双酶切 AmEBP1 质粒 DNA,DIG-dUTP 标记(地高辛杂交检测试剂盒),得到 1 200 bp 的片段为标记探针;使用美莱博 DIGD-110“地高辛杂交检测试剂盒Ⅱ(化学发光法)”(Sambrook et al.,1989)进行洗膜、信号检测。
1.5 转基因植株的抗寒能力测定
将 Southern 杂交检测为阳性的植株与对照,在培养基(改良 WPM + 6-BA 0.8 mg · L-1)上进行芽团扩繁增殖到一定基数,每瓶 4 株,改良培养基(WPM + IBA0.2 mg · L-1)上继续培养,选择长势相近的对照苗进行试验。抗寒能力测定参照孙海伟等(2012)的方法并作改进,4 ℃预处理 4 h,然后分别在–4 和–8 ℃冷冻处理 3、6、12、24 和 48 h,每处理 5 瓶。冷冻处理结束后,在 4 ℃,黑暗条件下解冻 3 h,然后 24 ℃光照培养 2 d,统计存活率。将 Southern 检测为阳性的植株与长势相近对照苗置于 4 ℃光照培养 24 h,然后置于 0、–4、–8、–12 和–16 ℃各处理 12 h,之后 4 ℃解冻 12 h,每个处理取 5 瓶,每瓶 4 株,采用膜渗透—电解质外渗法(郭卫东 等,2009)测定叶片相对电导率(REC)以表示细胞膜透性,重复 3 次,用 DPS 软件进行分析、比较。
MDA 的含量测定参照牛庆霖等(2013)的方法。根据测定所得的 REC,应用 EXCEL、DPS 软件进行非线性回归分析,参照郭卫东等(2009)的方法,测定转基因植株的半致死温度 LT50(Semi lethal temperature)。
2、 结果与分析
2.1 转 AmEBP1 基因组培杏苗的分子鉴定
2.1.1 PCR 检测
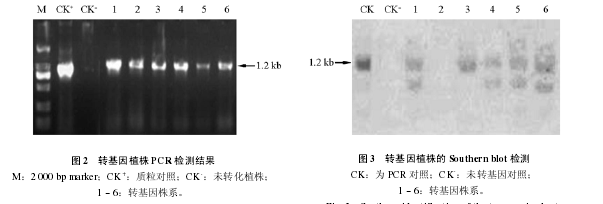
经花粉管通道法转化 AmEBP1 基因的‘大果’杏幼胚,在改良 WPM 培养基上由芽团进行多代扩繁。提取嫩叶 DNA 为模板,以载体质粒 DNA 为阳性对照,未转化的‘大果’杏为阴性对照,进行 PCR 扩增。结果显示在扩繁的材料中有 6 个转基因株系 PCR 结果为阳性(图 2)。
2.1.2 Southern 检测
将 PCR 扩增结果阳性的 6 个株系转基因‘大果’杏和对照植株同时采用地高辛标记法进行Southern 检测,结果表明,未转基因的对照植株没有出现杂交带,转基因植株中有 5 个株系分别出现 1 ~ 3 条杂交带,说明 AmEBP1 基因已经通过花粉管通道法整合到这 5 个株系的基因组 DNA 中(图3)。将阳性株系扩增到一定数量,用于下一步抗寒试验。
2.2 转基因株系抗寒能力检测
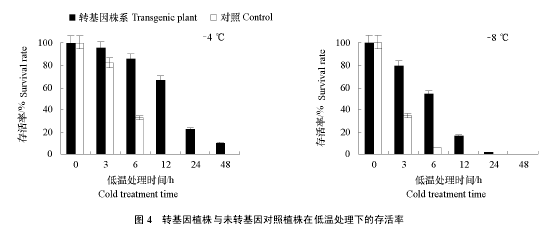
对得到的 5 个转基因阳性株系进行增殖扩繁,接种到改良 WPM + IBA0.2 mg · L-1培养基上继续培养。选取长势相近,苗高 8 ~ 10 cm,叶片浓绿,生长健壮的组培苗和对照苗进行抗寒试验。转基因植株在–4 ℃下处理 3、6 和 12 h 后,复苏培养均有较高的存活率(95%、85%和 66.7%),处理12 h 后存活率急剧下降,处理 48 h,解冻复苏培养后仍有 10.2%的植株存活;对照植株在–4 ℃处理 3 h 后受冻害现象明显增加,处理 12 h 后基本全部死亡(图 4)。–8 ℃处理转基因植株,3 h 后冻害明显,处理 12 h 后,解冻复苏培养仅有 16.3%的植株存活(图 4;图 5,A6h、A12h),24 h 后基本死亡(图 5,A24h);–8 ℃处理未转基因对照植株 12 h 后(图 5,B12h),基本不能复苏。以上结果显示:–4 和–8 ℃低温处理下,转基因植株比未转基因对照植株的存活率高。
2.3 低温处理对影片细胞膜透性的影响
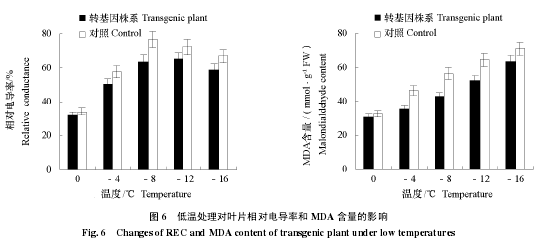
如图 6 可见,随着处理温度的降低,植物叶片受伤害程度加重,在–8 ~–16 ℃,转基因植株叶片相对电导率值一直低于相应的对照,说明转基因植株受低温损伤程度较对照较轻;对照 REC 值在–8 ℃时达到峰值,而转基因植株 REC 在–12 ℃时最大,二者峰值相差 11.2%;低温处理过程中,植物材料体内 MDA 一直在积累,在–4 ~–16 ℃条件下,对照植株始终高于转基因植株,说明转基因‘大果’杏低温处理下叶片损伤程度比对照轻。
2.4 转基因植株的半致死温度
应用电导法测定不同低温处理的植物组织,在不同温度下植物组织电解质外渗的累积量总是呈“S”形曲线,Lyons 和 Raison(1970)把低温对植物细胞膜伤害过程用 Logistic 曲线加以表示,并将曲线的拐点作为低温半致死温度。半致死温度主要反映了温度、水分与抗寒性之间的数量关系,大量研究表明,植株叶片半致死温度 LT50的高低可以表示抗寒性的相对大小(沈洪波 等,2002)。
本研究表明,转基因株系的 LT50是–5.80 ℃,未转基因对照为–3.67 ℃,理论上说明转基因株系的抗寒能力明显好于对照植株,这与转基因植株在抗寒试验中存活率的结果一致。

3、 讨论
温度是限制植物生长、发育及其地理分布的一个重要因素,低温造成的冻害对杏及其他果树和经济林木威胁很大(Shinozaki & Yamaguchi-Shinozaki,1996;Thomashow,1998)。关于木本植物抗寒基因的研究与草本植物相比较为落后。自费云标等(1994)从沙冬青中分离出了第 1 个来自于木本植物高热滞活性的 AFP,随后,Cao 等(2009)采用 cDNA-AFLP 方法,从沙冬青中克隆得到了能提高植物抗寒性的核糖体相关蛋白基因 AmEBP1,并试验证明 AmEBP1 增强了转基因植物的抗寒性是通过促进核糖体的合成以及冷诱导中转录因子和下游起保护作用的蛋白的翻译而进行的。随后沙冬青极强的抗冻能力引起了分子生物学家的极大兴趣,利用木本植物分离得到相关抗性基因,再转化到木本植物,现在已经成为植物抗性育种的新途径(Song et al.,2013)。近年来,果树上的转基因研究日趋深入,果树大多为杂合体,后代基因易分离,因此利用转基因手段,将目的基因转化到受体材料,按无性方式进行繁育,以获得目标性状的果树新品种。1988 年基因转化首先在核桃(Juglans regia L.)上取得突破,McGranahan 等(1998)获得了转 gus 基因核桃再生植株,现已在20 多种果树上获得了转化体或转化植株。王淑芳等(2001)将胆碱脱氢酶基因成功转化到番茄中,得到转基因番茄;王燕霞(2009)利用农杆菌介导法将抗寒转录因子 CBF 基因转入‘嘎拉’苹果中,PCR 检测并获得转基因阳性株系。中国在樱桃(Cerasus)、草莓、苹果等果树转基因方面做了许多研究工作,并获得了转基因植株,特别是樱桃的转抗菌肽基因已由农业部批准进入田间试验,该项研究处于国际领先水平。本研究中将源于沙冬青的抗寒基因 AmEBP1 转化到‘大果’杏中,探讨其转化后的抗寒性能,以期解决早春寒冷低温对冻花、冻果的影响,为抗寒杏品种的选育提供参考。
基因转化的方法主要有农杆菌介导和外源 DNA 直接导入,花粉管通道法是 DNA 直接导入的一种方式,其方法是将外源基因,通过花粉管进入胚珠,在植物体进行受精的过程中,胚囊内的精细胞与卵细胞均没有细胞壁,融合形成合子,早期未分化的合子近似于感受态细胞,易于接受外源DNA。方清(2002)将红树(Rhhoraa picutota)总 DNA 导入番茄,选育耐盐番茄;冯莎莎(2007)将 pWBvecloa 质粒 DNA 导入富士苹果,对转化植株进行了相关抗性分析;王晓蔓(2009)对影响核桃花粉管通道法转化因子进行了研究。花粉管通道法转化技术已经在果树转基因育种中有所运用,本研究利用花粉管通道法,将沙冬青抗寒基因 AmEBP1 转化到‘大果’杏中,利用幼胚培养,经卡那霉素筛选,PCR、Southern 检测得到 5 个转基因的阳性株系,并将阳性株系在改良 WPM + 6-BA 0.8mg · L-1)培养基上增殖扩繁,在改良 WPM + IBA0.2 mg · L-1培养基上生根成苗,得到转基因株系,进而对转基因组培苗的抗寒性能进行了分析。
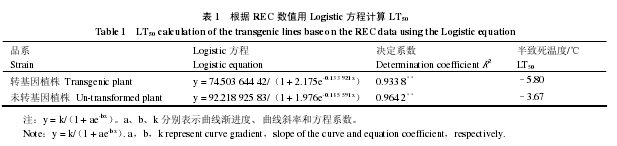
在木本植物抗寒性测定中应用电导法测定不同低温处理的植物组织,在不同温度下植物组织电解质外渗的累积量总是呈“S”形曲线,Lyons 和 Raison(1970)把低温对植物细胞膜伤害过程用Logistic 曲线加以表示,推导 Logistic 方程的二阶导数并令其为零,计算方程的拐点,得到 x = ln a/b,求出各株系相对电导率变化曲线的拐点温度,拐点温度就是该株系的 LT50,能较好地反映植物的抗寒力。本研究中转基因‘大果’杏的 LT50是–5.80 ℃,未转基因对照 LT50是–3.67 ℃,二者相差2.13 ℃,说明转基因‘大果’杏的抗寒能力较对照有了明显提高;抗寒试验表明,–4 和–8 ℃条件下,转基因‘大果’杏比对照有较高的存活能力;低温处理过程中,植物材料体内 MDA 与 REC 含量转基因‘大果’杏始终低于对照,说明转基因‘大果’杏低温处理下叶片损伤程度比对照轻,能够在相对的低温环境下生存。
本研究通过花粉管通道法得到 AmEBP1 转化的幼胚,又将幼胚培养成苗,并对转化苗的抗寒性能进行了初步分析,得到抗寒性能较好的‘大果’杏转基因植株,为下一步抗寒杏选育提供了参考。

山葡萄是东北地区的特产,可酿造出优质葡萄酒,深受广大消费者欢迎。山葡萄从建园开始,到进入盛果期,一般需5年左右。目前大面积栽培山葡萄多数采用篱架,少部分为小棚架。山葡萄整形与修剪目的在于调节生长和结果,合理配置枝蔓。1整形山葡萄在东北可以露地越...
辣椒属(Capsicum)主要有5个栽培种(邹学校,2009),即一年生辣椒(C.annuum)、中华辣椒(C.chinense)、浆果状椒(C.baccatum)、灌木状椒(C.frutescens)和茸毛椒(C.pubescens),其中一年生辣椒是分化最多、栽培最广的1个种,而中华辣椒是...
芋艿[Colocasiaesculenta(L.)S.]俗称芋头,为天南星科植物芋的块茎。万年芋艿属多子芋类中的香沙芋艿品种,主要分布在江苏海门、启东、靖江、上海崇明等长江下游地区。因生长于黏性壤土(俗称犟黄泥)中,其质地细腻,干香可口,易酥不糊,营养丰富,...
大马士革玫瑰(RosadamasceneMill.var.kazanlika)为蔷薇科蔷薇属灌木,花香纯正,是萃取玫瑰精油和加工玫瑰纯露的优良品种。玫瑰精油含有多种化学成分(如香茅醇、香叶醇等芳香物质、有机酸等有益美容的物质)[1],被认为是玫瑰精油的极品。大马士革玫瑰...
直播青菜是指在前茬作物收获后将青菜种子直接播种的一种轻简栽培方式,省去了育苗移栽程序。直播青菜具有省工、节本、高产、高效等特点,在国外应用较为广泛,但在我国应用不多。在长江流域青菜生产区,由于一直采取育苗移栽的方式,对直播青菜栽培技术研究...
番茄植株生长发育的最适温度为昼温24~26℃,夜温18℃左右,超过30℃则生长缓慢,到40℃就停止生长;地温的最高界限为33℃[1].然而,在光热资源丰富的华北地区,夏季温度偏高,温室6-8月温度很容易上升到35℃甚至更高[2],这样的温度对番茄栽培而...
长江流域及其以南地区,水资源相对丰富,许多地方适宜发展水生蔬菜种植,各地也有较多的地方水生蔬菜品种资源。慈姑作为水生蔬菜的一种,近年来越来越受到消费者的欢迎。长江中下游地区慈姑种植面积较广,生产上慈姑主要采用育苗移栽,对育苗环境和育苗技...

葡萄为温带水果,为葡萄科葡萄属木质藤本植物。葡萄种植过程中易发生多种病虫害,严重制约了葡萄产量和品质的提升。因此,探究葡萄病虫害绿色防控措施对促进葡萄产业绿色健康发展有重要意义。...
黄金梨(HuangjinPyrusL.)是韩国园艺场罗洲支场1967年用新高梨廿世纪梨培育成的中晚熟品种,1981年命名[1],1984年定名。1997年引入我国山东烟台。该品种为梨中珍品,外观诱人、品质优良、较耐贮运,是目前世界公认的高档优质梨新品种,具有较高的栽培推广...
随着社会经济不断发展进步,人们的物质生活水平有了显著提高,但同时经济发展和其他社会因素对城市原有的场所感弱化、人们生活环境质量下降、大众审美层面上的城市景观功能衰退,城市地域文化特征流失等负面影响也愈发凸显,从城市园林绿化角度来看,这主要...