��������������غ����ݶȵı仯����������������о���һ����Ҫ���ݣ����ζ����ַḻ�ȴ�ֱ�ֲ���ֵ�Ӱ����������λ�ú�ֲ�����Ⱥ�IJ�ͬ�����ڽϴ���졣Ŀǰ���������ַḻ�ȵĴ�ֱ�ֲ���֣���û���γ�һ�µĽ��ۡ���ˣ���չ��ͬ����ͬɽ���Լ���ͬ����Ⱥ���ַḻ�ȷֲ���ֵĸ����о����Բ������ַḻ���뺣�εĹ�ϵ����ʮ����Ҫ�����塣
������������صش�����������( ��ɳ����������ŭ��) ����������( ͼ1) ��λ���ȴ������ȴ��Ľ���������ɳ�������ϲ�Ͷ���У�ʹ�����γ�һ����Ϊ�������������������ع���ϡ����ͨ���㣬�� 20 ���� 80 ������������ڽ����˹���ѩɽ���������������� 3 ��ʡ����Ȼ��������ֲ�������Ϊ���; ���ӽ�ɳ���ӹȵ���ɽ��ʯ̲���� 4 000 m�ĸ߲�ʹ�ø��������������ţ�ֲ����ֱ����������������Դ�ḻ���ǵ�������������Ե���Ӱ�ʹ��������нϸߵ��о���ֵ��
����1 �о����ſ�
�������������λ��������������������������ݶ������ᡢ�����ش����ǽ���ش����ش����� 99°22' ��100°19'����γ 26°52' �� 28°52'��ȫ�����Լ 11 275km2��������ʡ��������ء��ؾ��ڵ��������ߡ����ϵͣ�����1 413 ��5 362 m���߲�3 949 m���س�פ�ؽ����� 3 270 ��3 300 m��������������Ҫ�����ϼ������֧���缱���Ľ�����ƣ���ʪ������; 6—10 ��Ϊ�꼾������Լռȫ��� 80%; 11 �������� 5 ��Ϊ����������࣬�����㣬����������ɽ�߹��������������: ���� 1 413 ��2 100 m �ĺӹ�����ƽ������ 14 ��17 �棬���� 2 100 ��3 000 m ��ɽ����ƽ������ 8 ��14 �棬���� 3 000 �� 3 300 m �ĸ�ԭ������ƽ������ 5 ��8 �棬���� 4 000 m �ĸ�ɽ����ƽ������ 0 �棬���� 5 000 m ���ϵ�ѩɽ��ƽ������ ��10 �� �� 8�档���ڽ�ˮ���溣�����߶�����������ÿ����100 m����ˮ������20 ��40 mm; ���ˮ�������ں��� 3 400 �� 3 600 m �������꽵ˮ���� 1 000 ��1 200 mm�������ֲ����سǽ�������Ϊ�磬���ϲ��ϸ߶��������ϵ�: ���ϲ�ɽ��Ϊ800 ��1 000 mm / a���ӹ�Ϊ 600 �� 800 mm / a; ������ɽ��Ϊ400 �� 600 mm / a���ӹ�Ϊ 300 �� 400 mm / a��
����2 �����
����2�� 1 ���鷽������������
������ 2001 �꿪ʼ������������ؾ��� 3 ��ʡ����Ȼ��������ֲ����Դ����ϵͳ���飬�ڴ˻����ϳ����ˡ����ϱ�������Ȼ����������2010 �� 7—8 �º�11 �£�2011 ��6—8 �º�10 �£�������鷶Χ��������Ϊ��λ���� 5 �ζ��������ȫ�ؿ�չҰ��ֲ����Դ���飬�������ߵ������������ 2 �ַ�ʽ���������� 96 ��( �̶����� 10 ��) ������ 51 �����ɼ�ֲ��걾 2 920 �š����� 2010 ��֮ǰ�ļ��ε��飬�ۼƲɼ�ֲ��걾Լ 7 400 �š�
�������ա�����ֲ��־�������й�ֲ��־����������ֲ��־���������б걾�������ٲ��䡶����ֲ��־���м��������������( ԭ�е���) �����ּ�¼�������������ֲ��������¼���ݿ⡣������������ص� DEM ���ݣ������о����ڵĵ����������������������ÿ 300 m ���θ̵߳�ͶӰ���������ͳ��ÿ 300 m ��ֱ�ݶȸ߳��ڷֲ�������ֲ�������������������������ֲ��ḻ�Ⱥ������ܶ��溣�θ̵߳ı仯���ɡ�
����2�� 2 ���ζλ���
������������غ���Ϊ1 413 ��5 362 m����300 m Ϊ���仮�ֺ��ζΣ����� 13 �Σ����� 1�����ݸ������ζεĺ��η�Χ������ GIS ���������� DEM ���в�ֵ���㣬�õ��������ζε���Ԫ������������Ԫ����ÿ����Ԫ������ʵ��½�������˵õ�ÿһ�����ζε�ʵ�����( ͼ 2) ��
����
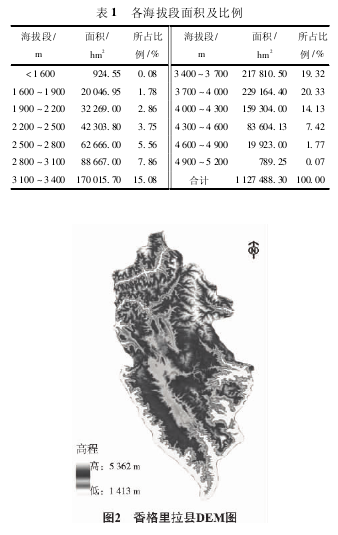
����2�� 3 �����ܶȼ���
�������ڸ����ζε������ͬ������ ��1 600 m �� ��5 200 m ��������ռ���������ȫ�ص� 0. 10% ������3 100 ��4 300 m �����ռȫ�������68. 85%����ȫ�ص���Ҫ����Ϊ��������������ַḻ�ȵ�Ӱ�죬�����ܶȼ��㹫ʽ����:
����D = S / lnA
����ʽ��: D Ϊ�����ܶ�; S Ϊ���ַḻ��; A Ϊ�о��������
����3 ��������
����3. 1 �ơ�������ͳ�Ƽ�������ϵ�ɷ�
ͳ���������������ֲ�����ݿ⣬�о�����������ֲ��3 794 ��( �����µȼ�) ��������163 ��876�������У�����ֲ�� 4 �� 13 �� 33 �֣�����ֲ�� 159�� 863 �� 3 761 ��; ����ֲ����˫��Ҷֲ�� 137 ��675 ����3 106 �֣�����Ҷֲ�� 22 �ơ�188 ����655 �֡�
��������������Ժʿ���й�����ֲ�����ֲ������͵Ļ��֣���������ص�����ֲ�� 876 ���ɻ���Ϊ 15 �����ͣ�������� 2��
����

����3. 2 �ơ������ַḻ���溣���ݶȵı仯
���������������ֲ�����ݿ��У�����ÿ���ֲַ��ĺ��η�Χ������ͳ��ÿ�����ζ�������ֲ��ơ������ֵ��������õ������ζοơ������ֵķֲ�����ͼ 3��
����

�����ơ������ַḻ���溣���ݶȵı仯������ȫ��ͬ���ƺ����ķḻ���溣�εı仯����ƽ���ĵ���ֲ���֣����ֵ��ƫ���ڵͺ��Σ���ƫ���ͣ�����1 900 ��2 200 m ���ζδﵽ��ֵ�������νϵͣ�ֲ�������Գ�����Ҷ�֡�ů������Ҷ��Ҷ�ּ����Ϊ�������ƿ�Ϊ�տ�( Compositae) ( 97 ��) ���̱���( Poaceae) ( 89 ��) ��Ǿޱ��( ��osaceae) ( 80��) ��ޤ��( Polygonaceae) ( 54 ��) �����ο�( Labiatae)( 49 ��) �����λ���( Papilionaceae) ( 48 ��) ��ëݢ��( ��anunculaceae) ( 39 ��) ������( Orchidaceae) ( 31��) ������ֲ������϶�Ŀ�; Ҳ�в��ֺ������ֽ��ٵĿƣ��������( Menispermaceae) ( 2 ��) ���������( Campanulales) ( 2 ��) �������( Scrophularia-les) ( 2 ��) �����ײݿ�( Saururaceae) ( 1 ��) ��������( Piperales) ( 1 ��) �ȣ���Ȼ��������������ֽ��٣��������϶ࡣ�����������棬�Խ�������( Psam-mosilene) ��è �� ʺ �� ( Decaisnea ) ���� � �� ( Kalo-panax) �ȵ�������Զ־��( Polygala) ��ľ����( Schi-ma) ����ǰ��( Cynanchum) ����������( Moneses) ��������ռ���ơ�
�����ֵķḻ���� 3 100 �� 3 400 m ���ζ���ﵽ1 444 ��( ������125 �ƣ�439 ��) ����һ���ζε����ռȫ������� 15. 08%����ֲ��������ů������Ҷ�ֺͺ�������Ҷ��Ҷ��Ϊ�����ֵĵ������߾�����ֵ���溣�����������½����ں��� 3 400 ��4 600 m���ֵ������½��� 1 159 �֣��������ֲ��������Ҫ�Ժ�������Ҷ�ֺߺ��ݵ�Ϊ����������ַḻ�����Ž��͡����� 5 100 m ����Ϊ�����ѩ��������û������ֲ��ֲ���
������ 3 100 ��3 400 m ���ζηֲ��Ŀơ����У����ƿ�Ϊ�տ�( 168 ��) ���̱���( 117 ��) ��Ǿޱ��( 101��) �����ο�( 59 ��) �����ο�( Scrophulariaceae) ( 58��) ������( 56 ��) ��ɡ�ο�( Umbelliferae) ( 56 ��) ��ëݢ��( 52 ��) ���ž黨��( Ericaceae) ( 41 ��) �����λ���( 37 ��) �ȣ���Щ�ƻ���������ֲ������ϴ�Ŀ�; ������Ϊ��������( Pedicularis) ( 43 ��) ���ž���( ��hododendron) ( 37 ��) ���������( Poa) ( 30 ��) ����������( Saxifraga) ( 26 ��) ��ޤ��( Polygonum)( 24 ��) ����ë���� ( Saussurea) ( 24 ��) ��������( Ligularia) ( 20 ��) �����IJ���( Arenaria) ( 18 ��) ��������( Aster) ( 15 ��) ��������( Cotoneaster) ( 14 ��)�ȣ���Щ����Ϊ�´����ʵ�����
�������Ͽơ������ַḻ������ֵ���ֵĺ��ζ��������о�������������ĺ��ζΣ�˵������ḻ�Ȳ���ֻ�����������Ӱ�죬��������ҲӰ�����ַḻ�ȵĴ�С��
����3. 3 �ơ��������ܶ��溣���ݶȵı仯
���������ܶȵļ�������ʾ���������ζοơ������ֵ��ܶ����ź����ݶȵı仯������ͬ�ĸ�֣������ź��ε����ߣ�3 ������ȼ����ܶ������ϳ������������½������ƣ��������ֵ���ֵ�λ��ƫ���ڵͺ��Σ�Ҳ�������ܶ��뺣�λ������ֵ����ϵ( ͼ 4) �����У��ƺ������ܶ����ֵ������ 1 900 ��2 200 m �ĺ��ζ�; ���ֵ��ܶ����ֵ������ 1 900 ��2 200��2 800 �� 3 400 m �� 2 �����ζΡ�
����

����3. 4 ���ķֲ��������溣���ݶȵı仯
����ͳ��ÿ�����ζθ��ֲ���������������ռ�ú��ζ��������ı������ɵó����ֲַ��������ڴ�ֱ�ݶ��ϵı仯��
����1) �ȴ����ĺ����ݶȱ仯���о������г�Լ290 km �Ľ�ɳ�����Ⱥӹȴ���ʹ�����ȴ����������϶ࡣ���溣�ε��������ȴ���( 2 ��7) �����Եݼ�����( ͼ 5) �����У��������ķ��ȴ��ɷ�( 2) ���溣�ε����ߣ��½������ԣ���Ҫԭ���Ƿ��ȴ��ɷֵ�����; �ȴ�������( 7) �ӵڶ����仯Ҳ��Ϊ���ԣ����Һ��� 4 000 m ���о����ȴ��������ķֲ�����; �������ȴ���( 4) �溣�����������½����ԣ����ں��� 2 800 �� 3 100 m ������ƽ���½���������3 400 m �����д����ͳ���; �����ȴ����溣�����߱��ֳ�ƽ�����½����ƣ��仯��ƽ�����ȴ����ĺ����ݶȱ仯��ӳ���ȴ��ɷ��溣�����߱����½����ձ���ʵ��
����2) ���´����ĺ����ݶȱ仯�����´��ֲ��ɷ�Ϊ���������ֲ����ϵ�����壬�����͵�������Ⱥ��Ĺ������������ԣ�ΪȺ��ṹ�и������Ҫ��ɳɷ֣��Ҳݱ����־Ӷ࣬�༯�����С��ߺ�����������ռ�������ź��ε������������������ơ������뱱����Ϸֲ����溣�������л����IJ������ں��� 2 200 ��2 500 m ���ﵽ���ֵ( 6. 14%) ��Ȼ�������٣����� 4 600 m ����Ϊ��ֲ����ޡ��´����������������´�������ռ�������٣��溣��������4 000 ��4 300 m ���ﵽ���ֵ����4. 76%��Ȼ������ 0( ͼ 6) ��
����

����3) �ŵ��к����ĺ����ݶȱ仯���������ʷ�ϵĸɺ���������Զ��ϵ�ĵ��к������������Ƿֲ������Ƿֲ��������������٣�һ���̶�����������ʷ�������������ص���������ʪ��״��������������ռ������������ 2. 7%���óɷ��溣���ݶȱ仯С�䶯( ͼ 7) ��
����4) ���������й��������ĺ����ݶȱ仯�����Ƿֲ�( 14) ������������Ϊ�´������ȴ�����������Ҷ�ֺ���Ҷ��Ҷ�ֳɷ֣����������������ͣ�������������������������ռ�ı����ϸߡ���ˣ���������صĶ��dzɷִ���ͺ��ε� 8. 45% ���������� 4 000 ��4 300 m ��15. 87%��������4 300m ֮���Ը�ɽ�ݵ��ɽ���Ϊ�����������������ٳ���( ͼ 8) ��
�����й�������( 15) �����溣�ε�������ƽ���������ƣ����ں��� 4 300 �� 4 600 m ���ﵽ���ֵ2. 97% �������й��������ж���Ϊ�����������γ��ڸ�ɽ��ʪ�����У��ں��νϸߵ����������Խϸߣ��������������٣�ֻռ�������� 3. 19%���������溣���ݶȵı䶯�������ԡ�
����5) ��ϵƽ��㡣��ͬ���ζ��´����ȴ��ɷֱ�����ͼ 9��
����

������ͼ 9 �ɿ������ȴ��������غ����ݶ����߽Ͼ��ȵĽ��ͣ���ͺ��δ��ı�����ߣ��� 56. 80%������ߺ��ζ� 4 900 ��5 200 m �����½�Ϊ 0; ���´����ı������غ����ݶȵ������ȶ����ߡ��ں��ε��� 1 600 m �������ȴ������´�������Ϊ1. 00�� 0. 76; �����ں��� 1 750 m �����´������ȴ�������������ȣ��ﵽ��ϵƽ���( floristic equilibriumpoint��FEP) ��
����4 �� ��
����1) �������������ֲ�����ַḻ���溣�ε����߳ʵ����֣����к������ַḻ��ˮƽ��ߡ��ơ���ˮƽ�ķḻ�ȱ仯Ҳ��ʾ�����Ƶ����ơ��ơ������ֵ��ܶ��溣�ε��������ʵ����֡������ݶ�ʵ���Ϸ�ӳ��ˮ��״���ı仯����ˣ����ε�Ӱ��ʵ�ʷ�ӳ���������ӵ��ۺ�Ӱ�졣�ܶ�����£��ڵͺ��ε�����ֲ�������������������ˮ�ֳ�������; �ڸߺ��ε�����ˮ�ֳ�����������ܳ�Ϊֲ��ֲ�����������; ���к��ε���������������ˮ�������õ�������������غ��θ߲�� 4 000 m������������ֵú����ԡ�
����2) �����м�����ЧӦ��˵���ܶ����ֵķֲ�������ص������к������ص�������ͺ�������ߺ������ص�����Խ���������������ַḻ�ȵķ�ֵ�������к���������˵�����м�����ЧӦ�dz��¶ȡ���ˮ�͵��ε�Ӱ�����ַḻ�ȷֲ���ֵĻ�������֮�⣬��һ���������ַḻ�Ⱥ������ܶ��뺣�γ��ֵ����ϵ��Ӱ�����ӡ��ٶ�����������غ��θߵ�2 ��������Ϊ“�м�����ЧӦ��˵”�������Ա߽磬���к������Ǵ���������ܹ����ɴ��������������γ����к��ζ�������ḻ�ĸ�֡�
����3) �ƺ����ĺ��ε����ֺ��ֵĺ��ε����ִ��ڲ��죬һ�������С��ߺ������ֲ��Ŀƶ�Ϊ�´����ʵĴ�ƣ��绢���ݿơ��տơ����οơ��ٺϿơ����IJݿơ��̶���; ���������������������������̶���������������ͷ��������������ί��������´����ʵĴ���Ϊ������ʹ��һЩ�����Դ�������ơ����λ��ơ��̱��ơ�ɡ�οơ�Ǿޱ�Ƶȵ���Ҳ��ֲ����о������С��ߺ������������С��ߺ�������ơ����ḻ�Ⱥ��ܶȽϵ͵�ԭ����һ���棬���о����ĵͺ�����������ɳ������ĸ�ů�ӹȣ��ֲ������ֳ����ִ�ơ������������ֺ�����Ϊ���Ƶ��������Ĵ����㲼���⣬����һЩ����ˮ������( Amethystea) ����ֲ���( ��hynchospermum) ������������è��ʺ����������ȵ�������Զ־����ľ��������ǰ���������������������Լ�һЩ���ȴ����ʵ���������( Euphorbia) ���������( Termina-lia) ����˿���� ( Cuscuta) ��ɽ���� ( Symplocos) �ȣ����Ƕ�Ϊ��С��( ��) ����Ȼ������ζ�����������Ҫ�����С��ߺ��ζΣ����ǿƺ���������ȴ�϶࣬�ɴ˵����о�����ƺ����ĺ����ݶȸ�����ֵĺ����ݶȸ�ִ������Բ��졣
����4) ��ϵƽ����Ǻ����ݶ����ȴ�ֲ����ϵ�ɷ����´�ֲ����ϵ�ɷ��仯�������߱�ֵ�����ӽ� 50%ʱ�ĺ��ζΡ��о��������о����ȴ������´����ı�ֵ�����ں��� 1 750 m ���ﵽƽ�⡣
�����ú��ζ��ڵ���Ϊ�������ʵĹ�����������ʵij�����Ҷ�ֻ�ů������Ҷ�ֵĽ��������Ǵ��ȴ��ɷ����´��ɷֵĹ��ɴ����ܶ��ȴ����ڸ�����չ�ֻ����������´������������������������( Aris-tolochia) �ȡ�Ұ����鷢�֣���������ص��еͺ��ζδ�կ��������Լ��У���Ϊ�Ƶ���������Ļ�㲼�Ĵ����ֽ϶࣬����Щ�����ɷֶ�Ϊ���ȴ��ɷ֣����ʹ�����ȴ��ɷ����ӣ���ʹ��ϵƽ�������������
�����۲� �� �� �ף�
������1�� �����꣮ ����ɽ�����ж�ֲ��Ⱥ������ԵĴ�ֱ�ֲ���֣�J�ݣ� ��������ԣ�2002��10( 1) : 7 ��14��
������2�� �Ŵ�ţ��ﺽ�� ��߶ȿռ���ֲ�����ַḻ���غ����ݶȷֲ���ֵ��о���չ��J�ݣ� ������ҵ��ѧѧ����2009��29( 2) : 74 �� 80��
������3�� ��ΰ���±������ŷ����ȣ� ���ϱ�������Ȼ��������M�ݣ�����: ���ϿƼ������磬2010��
������4�� �����ף� �й�����ֲ�����ķֲ������ͣ�J�ݣ� ����ֲ���о���1991��13( �� IV) : 1 ��139��
������5�� �����ף� �й�����ֲ�����ķֲ������͵������Ϳ����J�ݣ� ����ֲ���о���1993��15( �� IV) : 141 ��178��
������6�� �뽨�ϣ�����ƽ������ȣ� ������м�����ЧӦ��������������ֲ�����ַḻ�ȴ�ֱ�ֲ���ֵ�Ӱ���J�ݣ� ��������ԣ�2006��14( 2) : 107 ��113��
������7�� �뽨�ϣ���ɶ��� ����ҩɽ��Ȼ����������ֲ����ϵ�Ĵ�ֱ�ֲ���֣�J�ݣ� �人ֲ��ѧ�о���2008��26( 3) :271 �� 275��
������8�� �����������ף� ����ɽ����ֲ����ϵ���������ֲ�ͬ�����J�ݣ� ����ֲ���о���1997��19( 3) : 251 ��259