农艺学论文


在育种上,选择是指一个群体中的不同基因型的差别繁殖。选择是一项独立的育种技术,也是贯穿育种过程始终的步骤。每一种育种方法都离不开选择,每一个育种步骤都需要选择。正确的选择是植物育种成功的关键,选择的正确与否取决于育种目标和性状鉴定方法,育种目标规定选择的方向,性状鉴定和选择的方法决定选择的效率。
育种选择方法可分为两大类: 表型选择和基因型选择。
1 表型选择
[1]表型选择是指直接测定目标性状的表型值,根据表型值是否符合育种目标决定去留。表型选择多在目标环境下、性状充分表达的关键时期进行,表型选择的效率通常用选择进展或选择响应来衡量。选择进展是指入选亲本的子代平均表现值距原始群体的平均值间的离差。对于一个目标性状,选择进展的大小取决于 3 个因素: 狭义遗传力、选择差和选择强度。性状的遗传力是一个相对值,取值域为[0 ~1],选择差是入选群体的平均值与原始群体的平均值的离差,选择强度是指入选群体平均值相当于多少个原始群体标准差,实践中用入选率代表,入选率愈低,选择强度愈高,在相同的入选率下,供选择的群体愈大,选择强度也愈高。
尽管表型选择目前仍是育种上普遍采用的选择方法,但表型选择有以下难以克服的内在问题: ( 1)表型是基因型与环境互作的结果,不同的表型可能是基因型不同造成的,也可能是环境不同造成的,表型受环境影响,基因型和环境对表型的贡献大小未能分割,因此造成选择准确性不高,选择效率低。( 2) 表型选择类似于“黑箱”,不考虑目标性状的遗传结构,对控制该性状的基因数目以及各个基因的贡献大小、互作关系并不清楚,不能有效解析决定同一性状的基因之间的负相关( 一因多效、上位性、连锁) ,因而难以把控制同一性状的基因的最佳等位基因进行有效聚合,目标性状的改良速度缓慢、改良效果有限。( 3) 表型选择的前提是表型准确鉴定,要求表型鉴定结果精确、可靠。然而,目前还有许多性状没有建立有效的表型鉴定方法,而且,对那些已建立了表型鉴定方法的性状而言,在育种中进行表型选择也不容易,如根性状、许多种子性状。
为了提高表型选择的效率,可利用高度遗传相关的性状进行选择,其前提是相关性状的遗传力与目标性状与相关性状的遗传相关系数之积大于目标性状的遗传力。当目标性状与相关性状的遗传相关系数趋近于 1 时,相关性状的遗传力大小决定是否采用相关选择。
2 DNA 标记辅助选择
DNA 标记是 1980 年以来利用 DNA 核苷酸序列的差异( 多态性,如碱基替换、插入、缺失等) 建立的一类全新的遗传标记,数量无限,且不受环境因素或植物生长发育阶段的影响,因此可用不同发育时期、不同组织进行分析。
DNA 标记辅助选择是一种相关选择。DNA 标记遵循质量性状遗传,遗传力可看作为 1,所以能否利用 DNA 标记进行间接选择取决于遗传相关程度即 DNA 标记与控制目标性状的基因之间的连锁强度。
DNA 标记辅助选择可分为前景选择和背景选择( 图 1) 。【1】
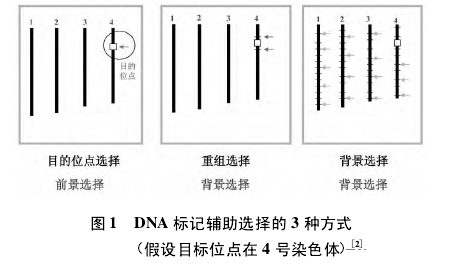
前景选择是对目标基因进行选择,首先要对目标基因进行定位,找到与目标基因连锁的标记,依据连锁的程度可将这种标记分为紧密连锁标记和完全连锁标记。完全连锁标记是根据控制目标性状的基因的序列开发的标记。随着越来越多的基因被克隆,开发完全连锁标记已成为一条重要途径。根据用于开发标记的差异序列是否导致功能变异( 功能基序) ,将完全连锁标记分为功能标记和基因标记。
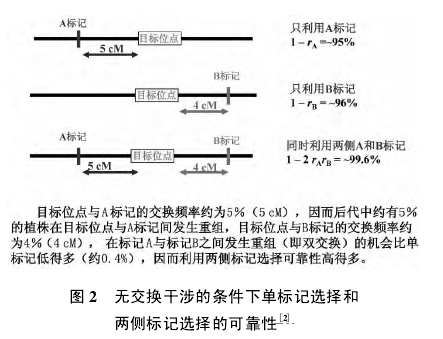
紧密连锁标记位于控制目标性状的基因的侧边,不是控制目标性状的基因的一部分,在减数分裂过程中与控制目标性状的基因会发生交换重组,因此这类标记叫做随机 DNA 标记。利用功能标记进行前景选择,选择准确率达到 100%; 利用随机 DNA 标记进行前景选择,由于不是完全连锁,目标基因选择的准确率取决于与标记的连锁程度以及是单侧标记辅助选择还是双侧标记辅助选择( 图 2) 。
背景选择是对遗传背景进行全基因组选择,要求用大量标记,并且要知道这些标记的位置和分布是否均匀。通过背景选择,可打破目标性状与不利基因之间的连锁,克服连锁累赘,提高选择精确性和速度。随着新一代测序技术的发展和以重测序、芯片技术为基础的高通量基因型分析平台的建立,对遗传背景的选择实现了全基因组均匀覆盖[3,4]。【2】
DNA 标记辅助选择可用于各种育种程序,如标记辅助回交育种[5]、标记辅助基因聚合、标记辅助纯系选育、标记辅助轮回选择等。
1记进行前景选择实现目标基因的快速转移,通过背景选择加快轮回亲本回复( 由 6 代减到 2 代) ,利用共显性标记可减少隐性目标基因回交导入时每一代的自交分离步骤和显性目标基因回交导入时为了纯合化的自交代数( 传统方法需要自交 2 代鉴定,而利用标记辅助选择只需自交 1 代) ( 图 3) 。
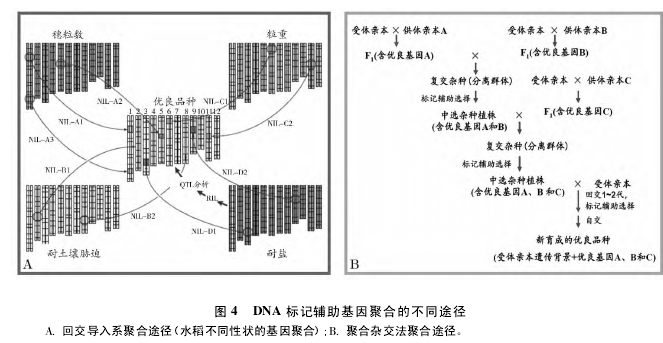
DNA 标记辅助选择能同时无损选择多个性状,用一片叶或半粒种子就可分析数十个性状,且能同时选择控制同一性状的不同基因和同一基因的不同等位基因,使得表型选择不能实现的控制同一性状的所有基因的最佳等位基因聚合得以实现。基因聚合可分为 2 类: ( 1) 控制不同性状的基因聚合; ( 2)控制同一性状的多个基因及其等位基因聚合。标记辅助基因聚合可采用不同途径: ( 1) 通过标记辅助回交,在导入一个目标基因的基础上,再与另一个含有目标基因的亲本杂交,对杂种后代通过标记辅助选择,用含有 2 个目标基因的植株与第三个含有目标基因的亲本杂交,重复上述步骤,直至将所有目标基因全部聚合。( 2) 为了聚合外源基因,可将栽培品种与野生种杂交、回交,培育含有某个野生种染色体片段的导入系,通过标记辅助选择鉴定导入系所含的导入片段并分析其表型效应,然后按照育种目标把控制同一性状或者不同性状的导入系进行杂交,通过标记辅助选择理想的重组单株实现基因聚合( 图 4A) 。( 3) 通过聚合杂交的方法,先将具有目标基因的亲本两两杂交,通过目标基因的前景选择培育出所有目标基因杂合的基因型,然后通过自交结合目标基因前景选择培育出所有目标基因杂合的基因型,实现基因聚合( 图 4B) 。在培育杂合基因型时,目标基因少的亲本先杂交,导致严重的相斥连锁的杂交先进行,每个世代多做杂交以保证有足够大的群体可供选择,如果所需群体过大时,可先回交。在培育纯合基因型时,一是可采取双单倍体技术,如果没有含所有目标基因的双单倍体,可将互补的双单倍体杂交进行下一轮选择,二是将尽可能多的少数基因杂合的基因型提升到下一代[6]。【3-4】

标记辅助纯系选育,一方面可用标记对亲本进行目标基因前景选择和遗传背景分析,另一方面是在早期世代( F2代) 进行标记辅助前景选择,提升理想的单株种成 F1株系或用于诱导双单倍体,从而减少种植规模( 从 2 000 株到 100 株) 。
标记辅助轮回选择是对亲本用标记进行目标基因前景选择和遗传背景分析,在基础群体如 F2群体通过标记辅助选择鉴别绝大多数基因具有有利等位基因的植株,如 2 亲本 F2群体各等位基因的频率为0. 5,10 个基因完全纯合的植株频率为( 1 /2)10,即1 024株有一个理想植株,如进行标记辅助选择,F2代将有( 3/4)10的植株入选,即大约 18 株有 1 株入选,其中10 个基因完全纯合的植株占( 2/3)10,即55株中有 1 株完全纯合,提高群体中理想基因型植株的频率。然后将上述入选植株进行杂交重组,并重1应小的基因控制的性状或一次选择多个主效基因时利用[7]。
3 基因组选择
基因组选择是 2001 年提出的一种基因型选择方法[8],它利用所有标记位点估测个体的育种值,根据育种值进行选择。与 DNA 标记辅助选择相比,基因组选择有几个特点[9]: ( 1) 不需要事先知道标记与性状之间是否关联; ( 2) 克服了等位基因多样性和遗传背景效应的影响,标记的效应,不像 DNA标记辅助选择,要么是0,要么是1( 依赖预设的显着性阀值) ,而是介于 0 ~1; ( 3) 仅估测育种值,选择育种值最高的单株,不能鉴定、导入新基因; ( 4) 育种周期更短; ( 5) 更适合于复杂性状的基因型选择。
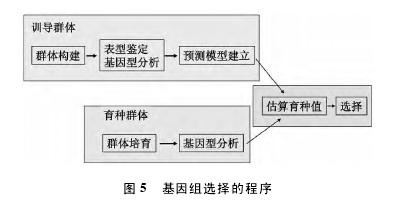
基因组选择的程序: ( 1) 构建与育种群体遗传相似的训导群体; ( 2) 对训导群体进行多点表型鉴定和基因型分析; ( 3) 利用大量标记位点构建表型预测模型; ( 4) 培育育种群体并分析标记基因型;( 5) 根据育种群体的标记基因型数据用预测模型估算育种材料的育种值; ( 6) 根据育种值的高低进行育种群体的选择( 图 5)[10]。【5】
基因组选择的关键在于预测模型的构建。现已构建了多种预测模型,并已开发出适合不同预测模型的计算机软件[9,11,12]。但不同预测模型假设性状的遗传结构不同,还没有一种预测模型适合所有性状、各类群体。每种预测模型都有优缺点,rr - BLUP预测对大多数性状而言是最准确的。各种预测模型预测的准确性( rA) 可用如下公式进行计算评估:【6】
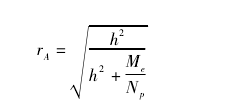
式中: h2为性状狭义遗传力,Np为训导群体的植株数,Me为独立的染色体片段数目,取决于有效群体大小( Ne) 和以摩表示的基因组大小( L) 两个因素,Me≈2 NeL,理想的状况是 Me与有效的数量性状位点数目( 数量性状位点方差和的平方与数量性状位点方差平方和之比) 有关。影响预测准确性的因素除了预测模型外,还与训导群体大小、训导群体与育种群体连锁不平衡( LD) 一致性、标记密度、性状的 h2和遗传结构等有关。训导群体愈大、训导群体与育种群体连锁不平衡一致性愈高、h2愈高、性状遗传结构愈简单( 涉及的位点数目愈少) ,预测的准确性就愈高。所需标记密度取决于有效群体大小与 LD 之间的关系,即相邻标记在 2 个群体中的平均 LD 要相等的话,每厘摩的标记数除以有效群体大小的结果应相同,基因组选择每 50 kb 一个标记,至少需要数百个到数千个标记,1 000 个位点控制的性状与 10 个位点控制的性状相比,要想获得同样的预测准确性,需要的标记密度要高 4 倍。用多个训导群体、在多个环境下对性状进行测定,也有利于提高预测准确性。
基因组选择应用的主要挑战是基因型与环境互作问题,其次是群体结构问题,还有就是表型鉴定的准确性和成本问题[9,11,13]。基因组选择可能导致稀有等位基因的丧失[9]。
4 展望
生物技术与信息技术各自的快速发展和它们之间的结合促进了基因型选择在育种上的应用。与表型选择相比,基因型选择的显着优势是: ( 1) 可在生长发育的任一时期进行选择,如种子表达的性状可提早到苗期选择,从而能快速淘汰非目标植株,甚或通过提取种子 DNA 可将下一代植株表达的性状提早到当代分析选择; ( 2) 不受环境影响,可准确测定单株基因型,可在早期世代进行异季选择; ( 3) 减少分离群体种植规模,因为在育种早期通过基因型选择能去除大多数分离后代,尤其是高度遗传性状的分离后代; 在植株发育早期通过基因型选择能通过高选择压,优化选择强度,对植株个体大的植物尤为有用。
基因型选择在精确性、准确性和育种效率上优于表型选择,甚至可以将质量性状和主效的数量性状位点的 DNA 标记辅助选择与复杂性状的基因组选择相结合,进行分子设计育种,即在定位控制所有农艺性状有关的基因位点并分析这些位点的等位变异及其对表型的贡献的基础上,设计理想基因型[14],利用 DNA 分子标记选择目标性状基因,将所有位点的有利等位基因组合起来,利用全基因组的DNA 多态性优化遗传背景[15,16]。
由于越来越多的作物已经完成了全基因组测序[17],基因组重测序的成本也越来越低,因此对大量种质资源( 如 1 000 份常用育种亲本) 进行重测序变得可能[18],将改变基因定位、亲本选配和后代选择的模式,通过考察这些进行了重测序的种质的表型,将序列变异与表型鉴定结果进行关联分析,找出功能标记,将这些功能标记输入育种群体进行表型预测检验,如得到证实,就可基于基因组序列进行选择[19]。
当前基因型选择未能普遍采用,除了经济成本偏高外,主要原因是育种家与生物技术专家之间的知识差距( knowledge gap) 、实验室到育种机构的应用差距( application gap)[7],推广基因型选择知识、开发简便实用的工具盒[20],将会加快基因型选择在作物育种上的应用。
基因型选择与表型选择并不是相互排斥的,何况当前基因型选择还面临着如何解决基因互作、基因与环境互作等问题,因此,在实践中可将基因型选择与表型选择结合用于复杂性状选择[21]。

我国是谷子的起源中心,谷子在我国有着悠久的栽培历史,且分布很广。我国的谷子种植面积占世界谷子总种植面积的80%。谷子具有抗旱节水、耐瘠、低投入的特点,是粮饲兼用作物。小米营养丰富且平衡,与大米和小麦相比,谷子蛋白质易于消化,生物价高。谷子曾...
引言作物育种学是农学、植保、园艺及作物育种相关专业的核心专业课程,肩负着引导学生了解作物育种并致力于作物育种事业的神圣职责[1]。结合教学目标及教材内容,当前作物育种学的教学任务主要是围绕育种目标制定、作物品种类型、品种改良或选育方法及生...
遗传标记就是遗传物质特殊的、易于识别的多态性表现形式,最初通过遗传作图应用于染色体上基因的定位。自1923年Sax[1]提出利用标记和遗传图谱进行辅助选择育种,以克服常规育种费时、费力等缺陷的遗传改良设想以后[1],基于DNA序列多态性而研究生物...
航天诱变育种是将植物干种子搭载返回式航天器(卫星),经过空间诱变作用产生变异,在地面选择有益变异培育新种质、新品种的育种方法,与常规育种相比,航天诱变育种具有诱变频率高、变异幅度大、育种周期短和有利突变体多等特点[1-7]。1航天诱变育种技...
太空环境具有高真空、微重力、地球磁场和高能带电粒子辐射等特点。众多复杂的空间环境因素对生物体的生长发育会产生极大的影响。1太空诱变育种的概念和特点太空诱变育种是指利用返回式近地卫星、高空气球等方式搭载生物体种质材料,在近地空间的物理和化学...
1作物育种方法研究概述通过对作物育种方法的研究,可以有效的提升作物的生产效率,为社会提供更加多的粮食储备,对于社会经济的发展和社会稳定性的维护都具有着非常重要的意义。针对这样的情况,在进行作物育种方法研究的过程中,就要充分的注意到对于高新...
品种对我国农业生产力提高的贡献率约为35%,而作物育种较发达的国家可高达50%,加快品种的优质化、专用化、多样化和特色化,尤其是优选,可以满足实际生产需要的优良品种,是育种工作的主要目标。近几年,由于我国城镇化进程加快,农村劳动力减少,人工成...
保障粮食安全是我国国民经济与社会发展的首要目标。我国人多地少,农业资源总量不足,人均资源量远低于世界平均水平,新时期粮食安全始终是关系我国国民经济发展、社会稳定和国家安全的全局性战略问题,因此实施国家粮食安全战略具有极端的重要性和紧迫性[1]...
我国科技查新工作开始于上世纪80年代,农业科技查新是一种建立在文献查询技术上,应用手工与计算机检索结合的模式为评价农业成果、技术咨询与专利申请提供资料的一种查询模式,下面就针对农作物育种科技查询工作的特点与方法进行深入的分析。1、农作物育种...
由于对作物氮吸收利用的作用机制尚不明确, 还难以应用于生产实际。因此, 必须加强对作物耐低氮性和氮效率的生理机制和分子生物学机理研究, 为筛选出耐低氮、高产氮高效两者兼顾的品种, 发展绿色、高效农业奠定基础。...