病毒学论文


猪瘟(Classical swinefever,CSF)是由黄病毒科瘟病毒属的猪瘟病毒(classical swinefe-vervirus,CSFV)引起的急性、热性、高度接触性病毒性传染病。猪瘟病毒为正股单链RNA病毒,基因组编码一个由3898个氨基酸残基组成的多聚蛋白,经细胞和病毒的蛋白酶催化裂解产生Npro、C、Erns、E1、E2、p7、NS2、NS3、NS4A、NS4B、NS5A,和NS5B等12个蛋白终产物。
其中,NS3是由2049个碱基编码的683个氨基酸残基组成的非结构蛋白。
NS3蛋白是一个多功能酶蛋白,具有丝氨酸蛋白酶(Serineprotease)活性、核苷三磷酸酶(Trip hosphatase,NTPase)活性和RNA激活的解旋酶(RNA helicase)活性,丝氨酸蛋白酶负责病毒多聚蛋白的加工,使之产生成熟的病毒蛋白,核苷三磷酸酶和解旋酶与病毒的复制相关。此外,NS3蛋白在病毒与宿主细胞的相互作用中也起到十分重要的作用。
在感染致细胞病变型的CSFV的细胞中,细胞病变效应越明显,检测到的NS3含量越高,表明NS3与CPE的发生有着紧密的联系。致细胞病变型的CSFV感染的同源细胞内NS2-3和NS3均出现,但是NS3含量明显高于NS2-3,从而推测猪瘟病毒蛋白NS3是导致细胞病变效应的一个重要因素,且NS3的浓度需要达到一定水平是导致CSFV感染的细胞中出现特异细胞病变效应的关键。
因此,本研究以猪瘟病毒NS3蛋白为对象,将其与真核表达载体pCMV-Myc重组,以构建真核表达载体pMyc-NS3,该载体的成功构建为下一步验证NS3蛋白与其宿主细胞相互作用的蛋白奠定基础。
1、材料与方法
1.1菌株、细胞和载体
PK-15细胞、293T细胞及pCMV-Myc真核表达载体由国家口蹄疫参考实验室保存,pG-BKT7-NS3质粒已构建好,E.coliDH5α菌株购自宝生物工程(大连)有限公司。
1.2主要试剂
ExTaq酶、限制性内切酶、DL10000 Marker均购自宝生物工程(大连)有限公司,T4DNA连接酶购自NEB公司,小量质粒提取试剂盒和DNA凝胶回收试剂盒购自Axygen公司,Lipo-fectamine2000购自Invitrogen公司,胎牛血清和MEM细胞培养基均为Hyclone公司产品,Anti-Myc抗体购自Clontech公司,HRP标记的二抗购自Sigma公司,FITC标记的荧光二抗购自Santacruz公司。
1.3NS3基因的克隆及序列鉴定
根据CSFVNS3基因(GenBank登录号AF092448.2)序列,运用Primer5.0设计1对引物,上游引物P1:5′-CGGTCGACCGGGCCTGC-CGTTTGCAAGAA-3′,划线部分SalⅠ酶切位点;下游引物P:5′-GCGGTACCTAGACCAAC-TACTTGTTTTAGTG-3′,划线部分为KpnⅠ酶切位点,引物由上海生工生物工程技术服务有限公司合成。以pGBKT7-NS3为模板,扩增NS3基因:94℃预变性5min,94℃变性50s,56℃退火50s,72℃延伸2min,35个循环,72℃延伸10min。PCR产物纯化回收后送至上海桑尼生物科技有限公司测序。
1.4融合表达NS3基因的真核重组表达载体的构建及鉴定
将载体pMyc-NS3和NS3PCR回收产物用SalⅠ和KpnⅠ酶切消化,酶切产物经10g/L低熔点琼脂糖凝胶电泳,胶回收酶切后载体和NS3基因并用T4DNA连接酶连接,转化DH5α感受态细胞,挑斑,提取质粒经酶切鉴定为阳性的质粒命名为pMyc-NS3,并送至上海桑尼生物科技有限公司测序。
1.5pMyc-NS3重组质粒转染
使用无内毒素质粒提取试剂盒提取pMyc-NS3重组质粒。按照Lipofectamine2000产品说明书,通过脂质体介导法将重组表达质粒pMyc-NS3转染至生长良好的PK-15和293T细胞。
1.6NS3蛋白表达产物的检测
转染24h后提取细胞总蛋白,Western blot对NS3蛋白的表达进行检测,经细胞裂解液裂解后取20μL加入等量的蛋白上样缓冲液中煮沸15min进行SDS-PAGE电泳。转膜,过夜封闭后加入V(一抗)∶V(PBS)=1∶100稀释的一抗,室温孵育2h,PBST冲洗3次,并在摇床上缓慢摇动,每次10min。加入V(辣根过氧化物酶标记的二抗)∶V(PBS)=1∶4000稀释的二抗,室温下摇床上作用1h。之后,用PBS冲洗3遍,每次10min。转移至底物溶液,室温下观察蛋白表达情况。
1.7间接免疫荧光观察与分析
细胞接种于12孔板,待单层细胞密度为70%左右时。转染质粒pMyc-NS3,24h后吸弃培养基,PBS洗3次,用φ=4%的多聚甲醛室温固定15min,PBS洗3次,再用φ=0.5%Triton-100的PBS室温透膜15min,PBS洗3次,室温封闭2h后,PBS洗3次,加入一抗过夜孵育。PBS洗3次,加入FITC标记的荧光二抗,室温避光孵育1h,PBS洗3次,在荧光显微镜下观察。
2、结果与分析
2.1NS3基因的PCR扩增
经琼脂糖凝胶电泳检测,可见与预期大小相符的2049bp左右的片段,基因序列测序结果显示该片段为CSFVNS3基因(图1)。
2.2pMyc-NS3重组质粒的鉴定
双酶切结果显示(图2),切出预期大小的2个片段。

双酶切和测序表明,NS3基因成功克隆至pMyc-CMV真核表达载体。
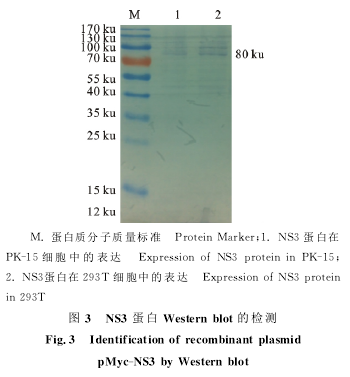
2.3融合蛋白NS3的Western blot鉴定
Western blot结果表明,转染pMyc-NS3的PK-15细胞中出现了与预计大小相符的80ku的融合蛋白条带(图3)。
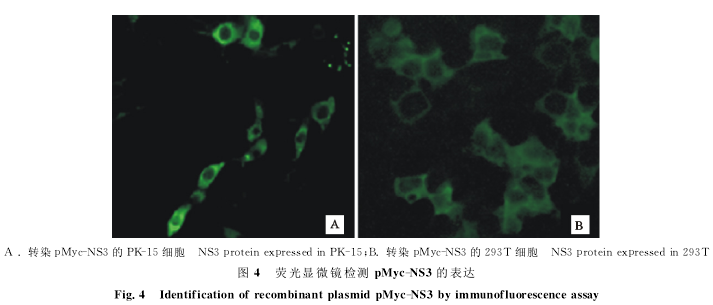
2.4融合蛋白荧光检测
将转染重组质粒pMyc-NS3的PK-15和293T细胞在荧光显微镜下观察,NS3蛋白在这两种细胞中均能表达(图4),NS3蛋白主要定位于细胞质。
3、讨论
NS3蛋白与猪瘟病毒体外致细胞病变有关,通过对NS3蛋白与细胞病变关系的研究,将有助于了解猪瘟病毒致病机理和持续性感染形成机制。在致细胞病变型瘟病毒感染的细胞中可检测到NS3和NS2,而非致细胞病变型瘟病毒在其感染宿主细胞中只表达NS2-NS3聚蛋白,因此非结构蛋白NS3可作为致细胞病变型猪瘟病毒在蛋白水平上的特异标志蛋白。猪瘟病毒NS3可能直接或间接作用于重要的宿主细胞蛋白而导致细胞受到损伤。
研究表明,黄病毒科成员C型肝炎病毒(Hepatitis CVirus,HCV)的NS3蛋白与诱导宿主细胞凋亡有关。
Prikhod’ko等将表达NS3蛋白的质粒转染至Vero细胞和HepG2细胞48h后,Vero细胞和HepG2细胞均发生凋亡。并证实NS3蛋白是通过与Caspase-8死亡效应结构域结合而诱导细胞凋亡,但NS3蛋白不能与其他带有DED的凋亡蛋白(如FADD)结合。Borowski等进一步证实,细胞中HCVNS3蛋白能与蛋白激酶A催化亚基特异性相互作用,从而抑制由AMP依赖性PKA介导的蛋白磷酸化。
因此,HCV感染细胞中的NS3可能通过抑制PKA的一系列过程而有助于HCV严重影响细胞正常功能。
酵母双杂交技术(yeasttwo-hybridsystem)是由Fields和Song在1989年通过利用酿酒酵母转录激活因子GAL4蛋白的特性时建立的,可以识别和分析相互作用的蛋白。但酵母双杂交系统也存在着一些局限性,如产生“假阳性”结果等。其结果往往需要一些其他方法的验证。
pMyc-NS3载体的成功构建以及在细胞内的表达,为进一步验证酵母双杂交筛选的与NS3蛋白相互作用的宿主细胞蛋白奠定基础。
非洲猪瘟(ASF)是由非洲猪瘟病毒(ASFV)引起猪的急性、热性、出血性和高度接触性传染病,无明显季节性,一年四季均可发生,通常在感染后10 d内死亡,病死率可高达100%...

寨卡病毒(Zika virus,ZIKV)是一种人类致病性蚊媒黄病毒。1947年,科学家首次从乌干达寨卡森林的恒河猴体内分离出该病毒。自发现以来,ZIKV仅在非洲、亚洲及南太平洋岛屿有小规模暴发,并未引起人们注意。2015年,ZIKV在南美暴发,造成数百万人感染。自此,Z...

水貂阿留申病(Aleutiandiseaseofmink,ADM)又称浆细胞增多症(Plasmacytosis),是由阿留申病毒(Aleutianminkdiseaseparvovirus,AM-DV)感染引起的水貂自身免疫系统功能紊乱,并发自身免疫的慢性、传染性疾...
前言肝星状细胞(hepaticstellatecell,HSC)活化及向肌成纤维母细胞(myofibroblast,MFB)转化是肝纤维化形成的中心环节,但机制复杂。既往对HSC基因表达及其调控的研究,主要集中于mRNA的编码区;然而,非编码小分子RNA在基因表达调控中的关键作用突破了传...
建立对多种病毒的快速、高通量、低成本的分子检测方法,对保障养殖鱼类健康、防止疾病发生具有重要的意义。淋巴囊肿病毒(lymphocystisdiseasevirus,LCDV)、大菱鲆红体病虹彩病毒(turbotreddishbodyiridovirus,TRBIV)、细胞肿大病毒属虹彩病毒(Meg...
痘病毒科(Poxviridae)病毒是在细胞质中复制的双链DNA病毒,大约编码200多种蛋白质。该科分为两个亚科:脊椎动物痘病毒亚科(Chor-dopoxvirinae)和昆虫痘病毒亚科(Entomopoxviri-nae)。脊椎动物痘病毒亚科包含八个属[1].许多病毒在进化过程中形成了有利...
猪流行性腹泻是由猪流行性腹泻病毒(PEDV)引起的,以猪呕吐、腹泻、脱水为特征的急性高度接触性肠道传染病,各种年龄的猪均可感染,以哺乳仔猪发病和死亡最为严重。PEDV属于冠状病毒科(Coronaviridae)冠状病毒属(Coronavirus),基因组为不分节段的线性单股正链RN...
猪流行性腹泻是由猪流行性腹泻病毒(porcineepidemicdiarrheavirus,PEDV)引起的一种以仔猪腹泻、呕吐和脱水为主要临床症状,最终破坏电解质平衡导致病猪死亡的的一种急性腹泻病[1].PEDV属套式病毒目,冠状病毒属,有囊膜,囊膜上包裹着花瓣状的纤突。其...
新冠病毒精准检测是加强疫情防控的重要手段。因此,有必要持续开展新冠病毒检测标准物质研究,完善新冠病毒精准测量系统,以更好地为我国新冠疫情防控提供全面的计量技术支撑。...

了解BVDV的分型、加深对BVDV病毒粒子结构和功能的研究, 有助于研制出针对流行BVDV病毒株的有效的疫苗, 以更好的预防控制BVDV的传播。...