����ѧ����


����-Ѫ�ܽ�����ϵͳ( renin-angiotensin sys-tem,RAS) �ļ��������Ѫѹ,Ѫ�ܽ����آ�( an-giotensin ��,Ang��) ��Ϊ��ϵͳ������ЧӦ����,���ϵ����궯��ƽ����ϸ��( human umbilical arterysmooth muscle cells,HUASMC) ȱ���յ����� 1( hy-poxia-inducible factor-1,HIF-1α) �ı���,����������Ѫ����Ƥ�������� ( vascular endothelial growthfactor,VEGF) ����,���д� HUASMC ��ֳ[1]������,�ǵ��¸�ѪѹѪ���������ӵ�ԭ��֮һ.HIF-1α ��ȱ�������е���Ҫ����,�������Ҫ����Ũ��Ӱ��,��������ת¼�ͷ���ˮƽ���� HIF-1α �ı���,�������λ�����е���,����һϵ�д���������Ӧ,������̬���ڡ�������Ӧ�ȷ��淢����Ҫ����.����,Ang����Ѫø�ȷ�ȱ���̼�Ҳ���ϵ�HIF-1α �ı���[2].Ang���ͨ����ͬ������ת¼����ˮƽӰ�� HIF-1α �ı���[3],Ҳ���ڷ����ˮƽӰ��ϸ�� HIF-1α ����̬,��: �ڷ����ˮƽӰ��HIF-1α ���ǻ�.�������ǻ�ø( prolyl hydroxylases,PHD) ��ȱ���յ����� 1 ��������( factor inhibitinghypoxia-inducible factor-1,FIH-1) ���ǻ� HIF-1α �Ĺؼ�ø.���鶯���� 3 �� PHD: �� PHD1��PHD2 ��PHD3,����Ի��Դ�СΪ PHD2 > PHD3 > PHD1,PHD2 �� HIF-1α �ĵ��ڳ���������,�dz���״̬��ά�� HIF-1α ��̬ˮƽ�Ĺؼ�����ø[4].����ʱPHD2 �� FIH-1 �ǻ� HIF-1α,ʹ֮Ѹ�ٽ���; ȱ��ʱ,PHD2 ʧȥ���ԡ�FIH-1 ø����������,HIF-1α��������,�ڰ��ڻ��۲�תλ���,�ٽ������ε�����Ӧ�����ת¼,����ϸ���Ե�����һϵ�з�Ӧ.Pagé ��[5]�о�����,�� HUASMC ��,Ang��ͨ������ϸ���ڿ���Ѫ��Ũ�ȸı� PHD2 �Ļ���,�ڷ����ˮƽ�ı� HIF-1α ���ǻ�,Ӱ����� HIF-1α���ȶ���,PHD2 �����뵰�ı��ﲻ�� Ang���Ӱ��.Ang��� HUASMC �� FIH-1 ��Ӱ�켰�������������δ������.���о���ͨ���۲� Ang���HUASMC �� HIF-1α��VEGF �Լ� PHD2��FIH-1 �� p-ERK �����Ӱ��,̽�� PHD2��FIH-1��p-ERK �� Ang��Ӱ�� HIF-1α �����е�����; ���� ERK �ź�ͨ·���Ƽ� PD98059 ��� ERK �ź�ͨ·����,̽��ERK ͨ·�� Ang �������½鵼 FIH-1 Ӱ�� HIF-1α�Ŀ��ܻ���.
����1 ���Ϻͷ���
����1. 1 ��Ҫ����
����HUASMC �������� Scien Cell ��˾; Ang�������� Sigma ��˾; ϸ����������̥ţѪ�塢�ȵ�������ø��RIPA �ѽ�Һ������� Gibco ��˾; ��ù�غ���ù�ع������� Sigma ��˾; �ÿ� HIF-1α���ÿ�PHD2 ������� Novus Bioscience ��˾; �ÿ� VEGF���ÿ� FIH-1 ����Ӣ�� Abingdon ��˾; HRP ��ǵĶ���������� Amersham Biosciences ��˾; PVDF Ĥ�������� Millipore ��˾; ECL ��ɫ�Լ��й��� GEHealthcare ��˾.
����1. 2 HUASMC �ĸ��ա���������������
������ϸ������ܴ�Һ����ȡ��,Ѹ�ٸ��µ� 37��,��ϸ����Һ���� 15 mL ���Ĺ�,1000 r/min ���� 5min,������,�Ժ� 20% ̥ţѪ���ϸ�������� 15 mL����,�Ƴ�ϸ����Һ,��5 mL ��Һ��ת��ϸ����Һ��һ����ϸ������ƿ��,���� 37�桢5% �� CO2������������.��ϸ�������ں� 80% ʱ,��������Һ,PBS����ϴ�� 3 ��,�����ȵ�������ø l mL ���� 2 ~ 3min ����ø,���뺬 10% ̥ţѪ���ϸ����������ֹ����,��5 mL ��Һ������,ʹϸ����ȫ����,���������.ȡ0.1 mL ����Һ,���������¹۲����,����ϸ����Ϊ5 ×10^8/ L,��ϸ����Һ�������µ�����ƿ��,ÿ24 h ��Һһ��.ϸ����Ϊ 3 ��: ( 1) ������: ��������������ϸ��6 h; ( 2) Ang����:�� Ang���������( ��Ũ��Ϊ 10^- 6mol / L) ����ϸ�� 6 h; ( 3) Ang�� +PD98059 ��: �� PD98059 �������� ( ��Ũ��Ϊ 10^- 5mol / L) Ԥ����ϸ�� 1 h ��,���뺬 Ang���������( ��Ũ��Ϊ10^- 6mol / L) ����ϸ�� 6 h.
����1. 3 Real Time PCR ��� HIF-1α��VEGF��PHD2 ��FIH-1 �������Trizol ����ȡϸ�� RNA,��ת¼�� cDNA,�� β-actin Ϊ�ڲ��� Real Time PCR,��¼ Ct ֵ,�ֱ���HIF-1α��VEGF �Լ� PHD2��FIH-1 �Ļ������.HIF-1α �ϡ���������ֱ�Ϊ 5'-TGC TTG GTG CTG ATTTGT GA-3'�� 5'-GGT CAG ATG ATC AGA GTC CA-3',���ﳤ�� 209 bp.VEGF �ϡ���������ֱ�Ϊ 5'-CCT CCG AAA CCA TGA ACT TT-3'�� 5'-AGA GATCTG GTT CCC GAA AC-3',���ﳤ�� 637 bp.PHD2�ϡ���������ֱ�Ϊ 5'-CTC ACT GAC CTA CGCCGT GT-3' �� 5'-CGC ATC TTC CAT CTC CAT TT-3'.FIH-1 �ϡ���������ֱ�Ϊ 5'-ACA GTG CCAGCA CCC ACA AC-3'�� 5'-GCC CAC AGT GTC ATTGAG CG-3'.PCR ����Ϊ: 95�� ���� 5 min,94�� 20s,72�� 20 s,72�� 5 min,55�� 10 s,40 ��ѭ��.�ڲ�β-actin �ϡ���������ֱ�Ϊ 5'-GTG GGG CGC CCCAGG CAC CA-3'�� 5'-CTT CCT TAA TGT CAC CCACGA TTT C-3',���ﳤ�� 540 bp,�� PCR ����Ϊ:94�� ���� 5 min,94�� 30 s,66�� 45 s,72�� 45 s,72�� 10 min,40 ��ѭ��.
����1. 4 Western blot ��� HIF-1α��VEGF��PHD2��FIH-1��ERK �� p-ERK �ĵ��ױ���.���鴦��ϸ����,��������,PBS ϴ2 ��,�� RIPA�ѽ�Һ,����30 min,4�� 12000 r/min ����20 min,��ȡ����Һ��װ�� -80�汸��.ȡ�ܵ��� 60 μg,100����� 5 min,10%SDS-�۱�ϩ����������Ӿ���뵰��,תӡ�� PVDF Ĥ,5%��֬ţ�̷��60 min,��һ��( ϡ�Ͷ�1��1000) ,4���ҹ,HRP ��ǵĶ�������1 h,TBS-TƯϴ5 ��( ÿ��5 min) .ECL �Լ�����Ӱ,X ��Ƭɨ���,Image J ͼ�������������.
����1. 5 ͳ��ѧ����
����������x ±s ��ʾ,SPSS 17.0 ��������ͳ�Ʒ���.�����Ƚ��� ANOVA �������,���Ƚ��ö������� t ����; ˫�� P <0.05 ��ʾ������ͳ��ѧ����.
����2 �� ��
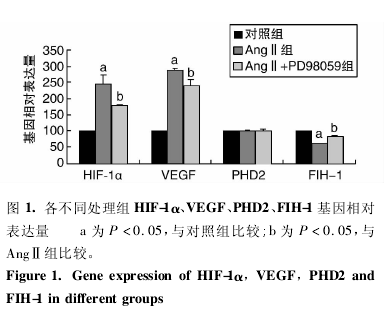
����2. 1 HIF-1α��VEGF��PHD2 �� FIH-1 ����ı���
�����������Ƚ�,Ang���� HIF-1α��VEGF ���������������,FIH-1 ������オ��( P �� < 0. 05) ,PHD2 ������������ű仯.�� Ang����Ƚ�,Ang�� + PD98059 �� HIF-1α��VEGF ����������Ž���,FIH-1 �����������( P �� < 0. 05 ) ,ͬ�� PHD2 ������������Ըı�( ͼ 1) .
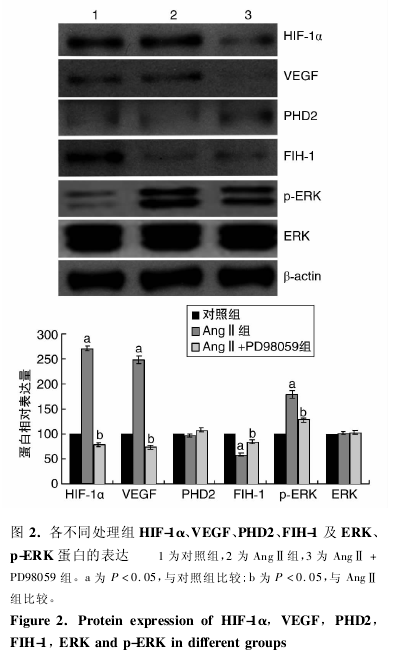
����2. 2 HIF-1α��VEGF��PHD2��FIH-1 �� ERK��p-ERK ���ı���
�����������Ƚ�,Ang���� HIF-1����VEGF ���ױ�����������,FIH-1 ���ױ������Ž���( P �� <0. 05) ,�� PHD2 ���� ERK ���ױ��������ű仯,�����ữ ERK ( p-ERK) ���ױ�����������( P <0. 05) .��Ang����Ƚ�,Ang��+ PD98059 �� HIF-1����VEGF ���ױ������Ž���,FIH-1 ���ױ�����������( P �� <0. 05) ,�� PHD2 ���� ERK ���ױ��������Ըı�,p-ERK ���ױ������Ž���( P < 0. 05; ͼ 2) .
����3 �� ��
������Ѫѹʱ Ang���������� AT1R ��Ϻ�ͨ�������ź�ͨ·�ٽ�ƽ����ϸ����ֳ������ĤǨ��,�ǵ���Ѫ�����������ԭ��֮һ.Ang���ϵ� HIF-1α�ı����ٽ� HIF-1α ���ΰл��� VEGF �ı���,��ƽ����ϸ����ֳ������Ҫ����.�������о�һ��[1,6],���ǵĽ����ʾ,Ang������ HUASMC ��HIF-1α �� VEGF �ı������������,˵�� Ang���ϵ� HIF-1α ����,���д� VEGF ���������.
����HIF-1α ��ϸ��ȱ��ʱ���յ������Ĺؼ�����,ͨ�����������ν� 100 ������ȱ����ػ����ת¼,��ȱ����Ӧ�Է�Ӧ������������Ҫ����.HIF-1α �����ˮƽ�ĵ�������Ũ��Ӱ��: ����ʱ,HIF-1α ����������������������ͨ·�������ø����,��������; ϸ��ȱ��ʱ,�ѻ��ڰ����� HIF-1α ���,��л��� DNA ���,������ת¼�����.
����PHD2 �Ƿ����ˮƽ���� HIF-1α ������ø,ͨ���ǻ� HIF-1α �ٽ��併��[7].���Ƿ���,Ang����HIF-1α ����������,�� PHD2 ����͵��ױ��������ű仯.���� Pagé ��[5]�о����,�迼�� Ang��Ӱ�� PHD2 ����͵��ױ���,�����ų��������ز��� Ang��� HIF-1α �ĵ�������.
����FIH-1 ����һ���ڷ����ˮƽͨ���ǻ� HIF-1α��Ӱ������̬�Ĺؼ�ø.FIH-1 �ĵ��ױ�������֯ϸ���бȽ��ȶ�[8],�Ե����������Ա� PHD2 ǿ,0. 2% �ĵ���������ʹ PHD2 ���ǻ�����ʧ��,��FIH-1 ���ǻ������Դ���[9],ȱ�����ı� FIH-1 ���ı���[10].���о����� FIH-1 ��ת¼������PKC �ź�ͨ·��Ӱ��,��� PKC ͨ���� FIH-1 ���� DNA �� CDP/Cut ����,������ FIH-1 ��ת¼����[11].�����ʾ,Ang�����Ž��� HUASMC �� FIH-1 ����͵��ױ����ͬʱ,���ż��� ERK �ź�ͨ·,���� p-ERK ���ױ���; �� ERK ���Ƽ� PD98059��� ERK �ź�ͨ·�Ļ��,p-ERK ���ױ������Ž���,FIH-1 �����ϵ�,HIF-1α��VEGF ������½�.
������ʾ ERK ͨ·�� FIH-1 ��Ӱ�������� PKC ͨ·��Ӱ����������: ERK �ź�ͨ·��������ữ�� ERK�鵼 Ang���µ� FIH-1 �ı���,ʹ HIF-1α �Ľ������,����ϸ���� HIF-1α ���ױ����Լ� VEGF ����͵��ױ���,������� Ang���ϵ� HIF-1α ���ױ���Ļ���; Ȼ�� ERK �ź�ͨ·�鵼 Ang���µ� FIH-1 ����ľ������û����д���һ���о�.ͬʱ,HUA-SMC �� HIF-1α ��������ϵ������ܵ�һ�ع���FIH-1 �����½�����,���� Ang������ת¼ˮƽ�ϵ��� HIF-1α �Ļ������[3].
������������,���Ƿ������ữ ERK �ź�ͨ·�鵼Ang���µ� HUASMC �� FIH-1 ����͵��ױ���,�ñ�����µ������� Ang���ϵ� HIF-1α ���ױ���Ļ���֮һ; ERK ���Ƽ������� Ang��� HUASMC �� HIF-1α��VEGF ������յ�����.��Щ���Ϊ���θ�ѪѹAng���Ѫ��ƽ����ϸ����ֳ�ṩ����˼·.
����[�����]
����[1]Richard DE,Berra E,Pouyssegur J. Nonhypoxic pathway mediatesthe induction of hypoxia-inducible factor 1alpha in vascular smoothmuscle cells[J]. J Biol Chem,2000,275 ( 35) : 26 765-771.
����[2]G��rlach A,Diebold I,Schini-Kerth VB,et al. Thrombin activatesthe hypoxia-inducible factor-1 signaling pathway in vascular smoothmuscle cells: role of the p22 ( phox) -containing NADPH oxidase[J]. Circ Res,2001,89 ( 1) : 47-54.
����[3]Pagé EL,Robitaille GA,Pouysségur J,et al. Induction of hypoxia-inducible factor-1 by transcriptional and translational mechanisms[J]. J Biol Chem,2002,277 ( 50) : 48 403-409.
����[4]Su Y,Loos M,Giese N,et al. Prolylhydroxylase-2 ( PHD2) exertstumor-suppressive activity in pancreatic cancer[J]. Cancer,2012,118 ( 4) : 960-972.
����[5]Pagé EL,Chan DA,Giaccia AJ,et al. Hypoxia-inducible factor-1alpha stabilization in nonhypoxic conditions: role of oxidation andintracellular ascorbate depletion[J]. Mol Biol Cell,2008,19 ( 1) :86-94.
����[6]Lauzier MC,Pagé EL,Michaud MD,et al. Differential regulationof hypoxia-inducible factor-1 through receptor tyrosine kinase trans-activation in vascular smooth muscle cells [J]. Endocrinology,2007,148 ( 8) : 4 023-031.
����[7]McMahon S,Charbonneau M,Grandmont S,et al. Transforminggrowth factor beta1 induces hypoxia-inducible factor-1 stabilizationthrough selective inhibition of PHD2 expression[J]. J Biol Chem,2006,281 ( 34) : 24 171-181.
����[8]Bracken CP,Fedele AO,Linke S,et al. Cell-specific regulation ofhypoxia-inducible factor ( HIF) -1alpha and HIF-2alpha stabilizationand transactivation in a graded oxygen environment[J]. J BiolChem,2006,281 ( 32) : 22 575-585.
����[9]Stolze IP,Tian YM,Appelhoff RJ,et al. Genetic analysis of therole of the asparaginyl hydroxylase factor inhibiting hypoxia-induc-ible factor ( HIF) in regulating HIF transcriptional target genes[J].J Biol Chem,2004,279( 41) : 42 719-725.
����[10]Fukuba H,Yamashita H,Nagano Y,et al. Siah-1 facilitates ubiq-uitination and degradation of factor inhibiting HIF-1alpha ( FIH )[J]. Biochem Biophys Res Commun,2007,353( 2) : 324-329.
����[11]Li J,Wang E,Dutta S,et al. Protein kinase C-mediated modula-tion of FIH-1 expression by the homeodomain protein CDP / Cut /Cux[J]. Mol Cell Biol,2007,27 ( 20) : 7 345-353.

�ֿ��������Ӧ���Ѫ���ع���Ѫ�������Ĺ�ϵ̽
�ֿ��������ӣ�RELM������һ������֢��ص�ȱ���յ���˿�������ӣ����д���ֳ��Ѫ��������Ѫ�����������������ӵ�����[1-4].RELM���зֲ������ԣ���֬���ļ���Ѫ�ܼ�����ȱ��������Th2ϸ���鵼�����Է�Ӧ�ķ���֯�б�������[5].��Ѫ��ƽ����ϸ����Ѫ����Ƥ...

֬������CTRP3����Ѫ�ܼ�������л�Լ����Ĺ�
���ż����ı仯�����Բ����������ǵĹ�ע�����Լ��������ص��ǽ�չ���������̳��������ʸߣ�ռȫ��������ԭ���60%���ϡ�����Ѫ�ܲ��������dz��������Բ�������Ӱ�����ǵ����彡����...
��������PTAH��Ⱦ�ļ��е����ú�Ӧ��
��������ʹҺ���м��Է��Ӹ�Ƶ������Һ����Ӽ����ײ���Ӷ�ʹҺ���ڲ�Ѹ�ٲ��Ⱥ�ijЩ��ѧ��Ӧ�ӿ졣�����������ô�ԭ��������������̶���֯���������Ϲ̶�Һ�̶���֯[1-2]�ȣ���ȡ�����õ�Ч��������û�������������Ϲ̶�Һ�̶��ļ�����...
̽��IL-17A�ڷ���ά�����������е�����
�ط��Է���ά��(idiopathicpulmonaryfibrosis��IPF)����δ�����÷��ڳ��ˡ������ڷβ��������Խ�չ����Ѱ���ͼ����Է���Ϊ����������һ�ּ����Էμ�...
��������������̥����ϸ���Ļ����о�
����������polychorinatedbiphenyls��PCBs����һ��ȫ���Ի�����Ⱦ���������ɲ������صĶ������ã������ǶԷ��������е�̥����Ϊ���У��ɵ����˶�ʧ���������ˡ������ٻ������̽��͵����ԣ�PCBs�����Ի����в����������ͨ�����ڻ���������...
̽��IL-37�Ե��˾���ϸ������ĭ�������ü���
��������Ӳ�������ӣ���һ���ۼ�ȫ��Ѫ�ܵ����Բ��䡣���Ӳ�������֬�ʴ�л���ң�ͬʱҲ��һ��������֢�Լ�������������ϸ����ϸ�����Ӳ������䷢�����չ������ϸ��Ǩ�Ƶ���Ƥ��ת��Ϊ�����������õľ���ϸ����ͨ����������壨���������������...
̽��BK��TGF-��1�յ���PASMCs��ֳ������
�ζ�����ѹ(pulmonary��artery��hypertension��PAH)��һ�����صĽ�չ�Լ���������Ҫ�����Ƿ�Ѫ���������������ߣ�������չ�ɵ��»�������˥�߶�������1�ݡ���Ѫ���ع��Ƿ�Ѫ���������������ߵ���Ҫ���������������ζ���ƽ����ϸ��(pulmonaryarterialsmooth...
���������������˿������ϵ����û�������
���������Ϊ������ҽѧԺ����ҽԺԺ���Ƽ�����Ҫ�����ǻ�����...

MSCs�����������µ����ߵ�����������
����ʸ�ϸ����mesenchymal stem cells,MSCs����һ�ֶ��ܸ�ϸ�����������Ҹ��ºͶ���ֻ�DZ�ܡ�1970�꣬AJ Friedenstein���״η�������Ϊ�ɹ�ǰ��ϸ���Ĺ���MSCs����ֱ��2006�����ϸ������ѧ��MSCsίԱ�������˽綨MSCs����ͱ���...
����FF����֢�Ĺ�ϵ�������û�������
����FF��neuropeptideFF,NPFF�����ڲ��鶯�������������ģ������ţ���з�����������ұ��ֳ��߶ȱ�����[1].NPFF��Ҫ�ֲ��������Լ�������ϵͳ�������Բ�λ������ߡ�2000�꣬Bonini��[2]�״η����NPFF����Ҫ��2���������NPFF1�����NPF...