发育生物学论文


引言
开花植物(被子植物)作为陆生植物中最大的族群,现已超过了250 000种。开花对于所有开花植物来说是生活史上的一个质变过程,是植物个体发育过程的中心环节;而对于人本身来说,色彩斑斓、气味芬芳的花不仅愉悦了人的身心,种类繁多的种子与果实也为人类提供了丰富的食物。故研究开花植物的开花过程,阐明其分子生物学上的调控机理无论在理论上还是在应用上都具有重要意义。Yanofsky 等(1990)在拟南芥(Arabidopsis thaliana)中首次克隆了花同源异型基因agamous(AG),标志着高等植物花发育研究进入分子遗传学阶段。
从发育生物学角度来看,高等植物经过一段时期的营养生长后,在合适的外界条件(其中重要的有日照长度、光质及温度)下,才能进行由营养生长(vegetative development) 向生殖生长 (reproductivedevelopment)的转变,才能开始花的发育。总的来说,花的发育过程在时间上大致分为4个阶段:(1)开花过渡(flowering transition),植株响应外界环境以及自身信号,由营养生长转向生殖生长,这个过程受一系列与开花时间相关基因的调控;(2)分生组织特征基因激活,植株响应从不同开花时间调控途径而来的信号,激活分生组织特征基因,决定分生组织属性;(3)花器官特征基因的激活,分生组织特征基因激活位于不同区域的花器官特征基因;(4)花器官形态建成,花器官特征基因激活下游的器官形态建成基因,决定组成各器官的特异细胞类型和组织(Jack, 2004)。
番茄(Solanum lycopersicum L.)是很重要的经济作物,同时也是用于双子叶植物花发育机理研究的一个重要模式植物。通过多年来不断的分子生物学上的深入研究,已有10个与番茄开花诱导及分生组织特征相关的基因被鉴定,将番茄与拟南芥相关基因比较发现两物种在花发育分子生物学上兼具保守性和多样性(表1)。本研究旨在对番茄花发育过程中开花诱导和分生组织特征决定这两个阶段已有的分子生物学研究进展,以及花器官特征决定、花器官形态建成阶段中各器官形态学特征进行综述,并与拟南芥上已有的研究做一个初步的比较,简要阐述两物种在花发育过程中的保守性及多样性。
过去20年,虽然对番茄花发育分子生物学的研究已取得了很多成果,但很少有对这些研究进展整合性的报道。本研究综述了番茄花发育开花诱导、分生组织特征决定这两个阶段已有的分子生物学研究主要进展,为对这番茄花发育调控网络进行更深入更精确的研究提供理论依据。
1 番茄花发育相关基因及其功能
植物花发育的每个步骤都受到严格的遗传控制,通过相应基因特定的时空表达及相互作用来精细调节花发生发育的网络。目前已鉴定出的与番茄开花诱导和分生组织决定相关基因.
1.1 开花过渡
植物的开花时间是由植物的发育阶段和环境因素共同决定的,是植物在进化和驯化过程中适应环境的一个主要性状。开花时间相关基因的突变会使突变体的开花时间提早或延迟。
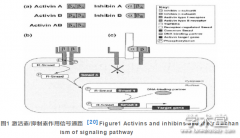
拟南芥是光周期敏感的长日照植物,茎尖分生组织(shoot apical meristem, SAM)先分化产生叶片,适宜的光周期信号存在时便会诱导向花序发育过渡。目前,拟南芥调控开花途径有6条,其中光周期途径、春化途径和温度途径响应环境信号,而自主途径(autonomous)、赤霉素(gibberellin, GA)依赖途径和年龄依赖途径调控植物的发育阶段(Wellmer, Riechmann, 2010)。拟南芥的开花时间相关 基 因 constans(CO)、flowering locus t(FT)、suppressor of overexpression of constans 1(SOC1)和agamous like 24(AGL24) 均促进开花,而 floweringlocus c(FLC)、短营养生长期基因 (short vegetativephase, SVP)则抑制开花;虽然多叶基因(leafy, LFY)也促进开花,但是其主要功能还是分生组织特征决定(图1)。叶是感应光周期的器官,长日照通过光周期途径加速开花,这条途径受CO(一个含有锌指结构和CCT域的转录因子)的调节,叶片在长日照的情况下会积累 CO,从而上调 FT 的表达(An etal., 2004);而 FT 是促进开花的一个决定性因素,FT蛋白是成花激素(florigen)的组成成分之一,成花激素是开花信号,叶片感应光照长度后诱导形成成花激素,通过韧皮部移动到茎尖,改变SAM中基因表达,从而诱导花的形成(Turck et al., 2008)。开花抑制基因 FLC 是自主途径和春化途径的整合基因(Amasino, 2010; Kim et al., 2009),在叶片和顶端分生组织中表达,是春化途径的核心组成,通过抑制FT 的 有 效 表 达 而 抑 制 开 花 ,防 止 提 早 开 花(Wellmer, Riechmann, 2010);另外春化可以降低FLC的表达,从而诱导叶片中FT表达以及茎尖从营养生长过渡到生殖生长(Searle et al., 2006)。另一个开花抑制基因是MADS-box基因SVP,FLC和SVP的功能部分相互依赖,它们之间的相互作用影响SOC1在茎尖的表达,并共同直接影响FT在叶片中的表达(Li et al., 2008)。SVP的表达受自主途径和GA途径的共同调控,同时也受温度途径的调控,在凉爽的天气下延迟开花,温度升高时则加速开 花 ,此 过 程 依 赖 于 FT(Wellmer, Riechmann,2010)。SOC1 受 GA 途径的正向调控,同时通过SPL (squamosa promoter binding factor-like)转录因子家族(如SPL9)受年龄依赖途径的正向调控(Lee,Lee. 2010, Wang et al., 2009)。SOC1 和 AGL24 之间形成了一个正反馈的循环(Liu et al., 2008),并组成复合物上调LFY的表达(Lee et al., 2008)。所有这些开花时间相关基因之间的多重相互作用形成一个网络,最终都归为花分生组织特征基因的上调表达。然而,由于已发现开花时间相关基因例如SVP、SOC1、AGL24和FT也参与花分生组织特征决定 (Gregis et al., 2008; Liu et al., 2009; Xi, Yu,2009),开花调节因子和花分生组织特征基因之间的功能区别越来越模糊。【图1】

番茄是日中性植物,与拟南芥不同,番茄开花主要受自身发育阶段和遗传背景的影响而不受环境因素影响,并且不管在长日照或是短日照情况下均可开花,但是长日照可使开花提前,同时番茄对光强度十分敏感。番茄上通常利用营养分生组织形成生殖结构之前产生的叶片数量表征开花时间(Dielen et al., 2003)。目前,虽然也已鉴定出一些与番茄开花时间相关的基因,但是这些基因调控的开花途径以及基因间相互作用的调控网络并不清楚。已鉴定的番茄开花基因falsiflora(FA)、单花束基 因 (single flower truss, SFT)、无 果 梗 节 基 因(jointless, J)、 复 合 花 序 基 因 (compoundinflorescence, S)和 uniflora(UF)均起到促进开花的作用,仅封顶花基因(terminating flower, TMF)为抑制开花基因(MacAlister et al., 2012)。这种情况可能是因为人为选择而导致生长周期不断缩短;而且大多数番茄栽培种在第三片叶开始膨大的时候就早早地过渡到开花,所以开花延迟的突变体比开花提前的突变体更容易形成(Evans, 1993)。
番茄中已鉴定出的开花调控途径有两条:光周期途径和自主途径。光周期途径受UF和S的调控,SFT和J都是开花自主途径中的调控基因,J的作 用 相 对 较 弱 (Dielen et al., 2003; Molinero-Rosales et al., 2004);J 和拟南芥 SVP 是同源基因,都是自主途径的调节基因,但是二者的不同在于J促进番茄开花而SVP抑制拟南芥开花。
UF是番茄光周期途径中的调控基因,uf突变体仅在冬季开花延迟,并且开花时间强烈依赖于每天的光能积累,主要通过糖的积累产生促进开花的信号(Dielen et al., 2003)。UF是调控番茄开花时间的关键基因,开花时间相关基因S和J均位于其下游,UF对这些基因的表达不具有直接调控作用,并且这些基因在番茄开花调控途径中具体的调控机理尚不清楚(Lifschitz et al., 2006a; Quinet et al.,2006a; Quinet et al., 2011)。s 和 uf 突变体均在冬季延迟开花,在低光照量的情况下s:uf双突变体比两亲本开花延迟更严重(Quinet et al., 2006a)。
番茄SFT蛋白是拟南芥FT蛋白的同源物,都是成花激素的前体。在拟南芥中,SFT过表达会导致植株开花提前,这结果与FT的过表达相似;SFT在 番 茄 上 过 表 达 同 样 导 致 开 花 提 前 (Teper-Bamnolker, Samach 2005; Lifschitz et al., 2006b)。
番茄中成花激素可代替UF调控的光周期途径所需的高光剂量(Lifschitz et al., 2006a),所以SFT可能是光周期途径和自主途径的整合因子。
番茄开花促进基因FA编码的蛋白(Molinero-Rosales et al., 1999)和拟南芥上的 LFY 蛋白功能相似(Weigel et al., 1992),FA虽然是开花促进基因,但其主要功能是分生组织特征调控,调节下游的花器官特征基因,作为花发生和花器官形成的连接因子,与拟南芥上的 LFY 的作用相似(Kato et al.,2005)。sft:fa 双突变体开花严重延迟,形成 100 片叶 后 也 未 开 花 (Molinero- Rosales et al., 2004;Thouet et al., 2012),这表明 SFT 和 FA 位于两条不同的通路上调控番茄开花并相互协同,这与LFY 和FT在拟南芥中的关系类似。有趣的是,在拟南芥中同时过表达LFY 和FT可促使植物在形成子叶后提前开花(Kardailsky et al., 1999; Kobayashi et al.,1999)。目前并没有研究表明 fa 突变体开花延迟与季节(即光照强度)有关,故不能确定与FA相关的开花调控途径。tmf是目前唯一开花提前突变体,TMF开花抑制作用与促进开花的成花激素途径并没有关系。
同时,tmf花序突变为单式花,并且形成变态大叶状萼片,在所有单式花花序突变体中(s和uf)仅有tmf开花提前(MacAlister et al., 2012)。
1.2 分生组织特征决定
在大多数植物中SAM是无限生长的,营养分生 组 织 在 开 花 诱 导 后 成 为 花 序 分 生 组 织(inflorescence meristem, IM),随 后 花 分 生 组 织(floral meristem, FM)在 IM 上形成取代叶的侧生器官(Chandler, 2012)。IM 中央的干细胞增殖与 IM外侧的 FM 发生在时间和空间上是相对独立的(Chandler 2011)。
拟南芥SAM在营养发育阶段形成莲座叶,随后环境因素诱导进入生殖发育阶段(Siriwardana,Lamb, 2012)。拟南芥花发生形态学上大致分为三个阶段:第一阶段IM外缘凸起发生,即苞片发生;第二阶段在IM与外缘凸起之间形成一半球形平滑的凸起,即FM发生;第三阶段FM侧面萼片原基初始形态出现,即萼片原基发生(Smyth et al.,1990;Kwiatkowska, 2006)。 分 生 组 织 特 征 基 因(meristem identity genes)是决定从开花过渡到花器官形成的关键基因,控制 IM 形成 FM 这一过程。
拟南芥中已鉴定出的 FM 特征基因 leaf(LFY)、apetala 1(AP1)、cauliflower(CAL)、fruitfull(FUL) 和unusual floral organs(UFO)均促进 FM 的特异分化(Schultz, Haughn, 1991; Bowman et al., 1993;Wilkinson, Haughn, 1995; Ferrándiz et al., 2000),它们之间的相互作用见图1.LFY诱导花原基的启动,使植物对开花做出响应(Simon et al., 1996)。同时,LFY 对另外的 FM 特征基因--AP1,CAL 和FUL的表达具有直接上调作用,这3个基因均属于MADS 结构域家族,其中 CAL 与 FUL 功能冗余(Ferrándiz et al., 2000,William et al., 2004)。随着AP1的表达,AP1反过来抑制开花诱导基因SOC1(Lee et al., 2010)。同时 AP1,CAL 和 FUL 可以进一步反过来上调LYF的表达,直到LYF的表达水平超过实际花发生所需的阈值(Blazquez et al., 1997,Ferrándiz et al., 2000)。LFY 编码的转录因子通过LFY 蛋白在细胞间的移动成为胞间信号,在 FM 细胞层中或之间非细胞自主性地扩散(Sessions et al.,2000, Wu et al., 2003)。AP1 和 CAL 是同源基因,决定IM上FM的发生(Ferrándiz et al., 2000),在开花第一阶段的FMs中二者的表达活性随LFY的上调而上调(Wagner et al., 1999, Urbanus et al., 2009)。
LFY、AP1分别通过late meristem identity 1 (LMI1)、LMI2 前馈调节 CAL(Saddic et al., 2006, Pastore etal., 2011)。与拟南芥不同,AP1 在番茄中是组成型表达,在第6个营养生长点之后便使植株开花,早于野生型的12个营养生长点,表明AP1在番茄上具有促进植物开花的作用(Ellul et al., 2004)。FUL是一个MADS结构域蛋白,在IM中出现且不在早期FM中出现(Gu et al., 1998)。
拟南芥中 IM 特征基因有 agamous- like 24(AGL24)和 terminal flower 1(TFL1),这些基因的表达保持IM的特征,却抑制了IM中FM特征基因的表达(Shannon, Meeks-Wagner 1993, Yu et al., 2004)。
同时 FM 特征基因 LFY、AP1 和 CAL 抑制了 FM 中AGL24 和TFL1的表达。已有研究表明IM和FM特征基因表达之间的相互作用,决定两种分生组织的边 界 (Weigel et al., 1992, Liljegren et al., 1999Ratcliffe et al., 1999)。TFL1 在 SAM 和 IM 中均有表达且编码同一种移动蛋白(Conti, Bradley, 2007),TFL1 蛋白是花序发育中的转录抑制因子(Hananoet al., 2011)。拟南芥 FM 特征受一个大型且部分冗余的基因网络调控,确保分生组织特征发育的确定性和稳定性,防止转回营养生长的可能。
番茄是合轴生长的植物,与拟南芥单轴生长不同,茎的生长是一个营养生长和生殖生长规律交替的过程。番茄SAM在产生5~12片叶后,形成一个顶花序便完成了主茎的生长。随后由花序下最后一片叶的腋生分生组织(axillary meristem)形成合轴分生组织(sympodial meristem, SYM)继续植株的生长,形成2~3片或更多叶片后再次形成一个花序完成第一段合轴茎生长,如此循环无限生长,形成总状花序(Pnueli et al., 1998)。番茄花发生大致分为三个阶段:第一阶段SAM产生第一个IM和FM,与此同时最后一片叶的腋生分生组织形成合轴分生组织;第二阶段IM上第二个FM发生;第三阶段第一个FM上萼片原基发生(Allen, Sussex, 1996)。
番茄中已鉴定出的 FM 特征基因有 FA,S 和anantha(AN);IM 特 征 基 因 有 SFT,UF、J 和macrocalyx(MC);另外,blind(BL)和 self-pruning(SP)基因与番茄合轴分生组织发育相关(表1)。番茄中开花时间相关基因全部参与了生殖分生组织生长发育,这些基因在整个花发育过程中表现出多功能性,并且各功能之间没有相互联系,这说明番茄花发育过程受一个复杂精密的基因网络调控。
s突变体花序严重分枝,单花序载花量达到200左右,这种突变表型是因为s在IM形成之后随即分化为两个IM,不同于野生型分化为一个FM和一个IM,造成突变体严重分枝(Quinet et al., 2006b)。同时,分生组织成熟延迟也是造成s严重分枝的原因,首先尖端分生组织成熟延迟便分化出更多的IMs,其次IM延迟成熟产生更多的FMs从而造成突变体严重分枝(Park et al., 2012)。尖端分生组织和侧生分生组织均延迟成熟造成s严重分枝,与此发育机理不同,野生番茄品种花序适当分枝是由尖端分生组织成熟延迟造成的(Park et al., 2012)。
j和sft的突变体同时表现为花序转回营养生长。j突变体的花序生长1~3朵花后便转回营养生长形成叶状花序,但植株合轴生长的习性并未改变(Szymkowiak, Irish, 2006),这与sft不同。
sft突变体构型更为复杂,突变花序存在两种表型:花序上的营养分生组织(vegetative meristem,VM)形成局部顶端优势抑制SYM生长,植株的垂直茎由花序继续营养生长构成,形成“假茎”(pseudoshoot)(Molinero-Rosales et al., 2004; Lifschitz et al.,2006a);另一种情况是虽然 sft 花序转回营养生长,但是和j类似,侧生生长位点并未被抑制继续合轴生长(Thouet et al., 2012)。这两种突变花序类型同时存在于sft植株中,sft突变表型的多样性与环境因素紧密相关,低光照条件下sft花序突变为单式花的频率显着增加(Quinet et al., 2006b)。j和sft对花序结构的相似影响表明J和SFT可能存在功能冗余,二者均有抑制IM营养生长的功能。s:sft双突变体的花序与s类似形成严重分枝,与s和sft均不同的是s:sft形成大量花和叶混合生长的花序,说明在花序表型特征上s和sft之间加性作用(additive);j:s 双突变体表型与 j 一致说明 j 是 s 的上位基因(epistatic; Thouet et al., 2012)。
J是形成花梗离层的关键基因,同时也是番茄中形成复合花序的关键基因。J通过抑制FM成熟促进IM构形,番茄花发育第一阶段SAM形成一个FM 和一个 IM,此时叶片中形成的 SFT 蛋白作用于分生组织中一个未知蛋白,此蛋白与J共同参与形成复合物通过抑制FM特征基因FA从而阻止FM成熟促进IM形成,因为FM生长能力有限而IM可无限生长的特点故抑制FM成熟促进IM生长有助于形成复合花序(Thouet et al., 2012),故IM特征基因J的功能可能是通过抑制IM营养生长以及FM的成熟来促进花序形成。
fa和an突变体不能形成花,fa花序完全营养生长发育成多叶分枝,而an的IM不形成正常的FM,细胞增殖生长成为菜花状(cauliflower-like)的变态花 。fa 突变体中因为基因缺失导致的移位,致使由412个氨基酸组成的蛋白仅有187个氨基酸组成,这可能是导致花数量显着减少的原因(Molinero-Rosales et al., 1999)。fa和an双突变体的表型与fa表型一致说明fa是an的上位基因(Allen, Sussex, 1996)。
UF作为调控番茄开花的关键基因同时也是花序发生的决定性基因,uf 突变体花序为单式花(Dielen et al., 1998, 2001, 2003)。可能有两种原因导致这种突变表型:一种是由于花序被单式花取代,故UF是IM特征调控基因(Dielen et al., 1998);另一种可能性与sft类似,uf突变体的IM在第一个FM 发生之后转回营养生长形成叶,然后 SAM 继续生长形成假茎(Lifschitz et al., 2006a)。增加每日光照剂量可挽救uf开花延迟表型,但对单式花突变表型并没有影响,说明UF基因的两个功能促进开花和决定IM特征之间并没有相互联系(Dielen et al.,2003)。同样,提高 SFT 在 uf 中的表达量可使开花延迟的突变表型逆转为开花提前,但是只能部分挽救uf的单式花突变表型(Lifschitz et al., 2006a)。uf:
sft不能形成花表明UF和SFT位于两条平行的调控通路,UF 并不直接调控 SFT 的表达(Quinet et al.,2011)。uf 与其它基因(sp、bl、j 和 s)的双突变体以及j:uf:sp和uf:bl:sp三突变体均与uf一样产生单式花(Quinet et al., 2006a, 2011),表明 UF 位于 BL、SP、J和S的上游。
BL和SP是调控番茄合轴分生组织生长的关键基因,也是调控番茄生长习性的重要基因。sp突变体中由于一个CETS 家族蛋白的变异导致最后连续的合轴茎产生叶片数量逐渐减少,最终以两个连续的花序打顶使无限生长变为有限生长(Pnueli etal., 1998, 2001)。bl 突变体中,由于 R2R3 家族 Myb基因功能缺失导致侧生分生组织部分缺失而不能形成腋芽,在形成一个花序之后因为腋芽缺失不能继续合轴生长成为有限生长类型。
j:sp部分挽救了sp合轴区段有限生长的突变表型,同时也部分挽救了j的叶状花序突变表型,这说明J和SP间存在较为复杂的相互作用关系(Quinetet al., 2011)。
2 番茄花发育过程的形态学特征
番茄属于被子植物中的双子叶植物,花器官发育遵循典型的双子叶植物发育模式。典型的双子叶植物的花发育的基本单位是同心轮,由4个不同器官在花柄顶端按四轮分布,由外到内第一轮由绿色叶状的萼片组成,第二轮是花瓣,第三轮是雄蕊,即产生花粉的雄性生殖器官,在最里面一轮是心皮,是雌性生殖器官,胚珠和种子在里面形成。
Brukhin(2003)通过电镜扫描和组织学分析,观察番茄花原基发生期至盛花期的花被(全部的萼片和花瓣)形态、花药和雌蕊的发育过程,将发育过程分为20个阶段:(1)萼片原基发生;(2)花瓣原基发生;(3)雄蕊原基发生;(4)心皮发生;(5)子房腔出现,花药开始分化;(6)花药横切面呈双浅裂形(bi-lobedshape),胎座原基发生;(7)花药中小孢子母细胞(pollen mother cells, PMC)和绒毡层分化,心皮开始融合;(8)绒毡层细胞双核化,胼胝质在PMC周围沉降,花柱萌发胚珠原基发生;(9)PMC开始减数分裂,大孢子细胞分化;(10)小孢子进入四分体时期,胼胝质在大孢子母细胞(megaspore mother cells, MMC)周围沉降;(11)小孢子周围胼胝质被吸收,小孢子得释放;胚珠开始减数分裂,珠被形成;(12)花萼微开,珠柄血管化;(13)胚囊分化;(14)萼片分离,小孢子形成大液泡并呈近球形;珠被生长至将珠心包于其中;(15)花瓣变为黄绿色,花药中绒毡层完全退化;(16)花瓣微开;雄蕊发育至最长,花粉开始有丝分裂;珠孔形成;(17)花瓣尖端变黄;花粉粒发育成双核细胞;(18)花瓣变黄;花粉粒双核分化,营养细胞核和生殖细胞核形成;倒生胚珠发育完成;(19)萼片发育至最长;脂质体在花粉中累积;(20)花冠完全张开;花药开裂,成熟花粉释放(Brukhin et al., 2003)。从以上的发育过程可看出番茄花形态建成有以下几个特点:(1)每轮花器官均分别由其各自的原基发育而来,并无共同原基的出现;(2)四轮花器官萼片、花瓣、雄蕊和雌蕊的发生是由外向内逐渐发生的;(3)每一轮花器官基本是同时发生,并且每一轮花器官的属性一致。这些特点与拟南芥花形态建成过程类似(Smyth et al., 1990)。
与拟南芥不同的是,萼片是番茄主要商品器官--果实的重要组成部分,萼片从盛花期至果实成熟期形态特征的研究显得尤为重要。郑戌翔等(2011)对番茄果实成熟期的萼片形态进行了初步观察,并分为了基平、上翘、直立和上卷4 种形态类型。番茄花器官发育的这种特殊性揭示了番茄花发育过程可能涉及比拟南芥更为复杂的分子遗传机制。
3 问题与展望
作物花发育过程与作物丰产、早熟育种息息相关。番茄作为研究植物花发育过程的模式植物,近年来随着获得越来越多花发育相关的突变体,对花发育潜在分子机制有了更深入的理解。目前相关基因的鉴定还远不完全,已鉴定基因的调控机理和功能尚不明确,基因间形成的调控网络有待研究,但是利用已有的研究成果可假设调控网络模型并进行验证,能对控制番茄花发育基因网络的组成和结构有更详细和准确的理解。拟南芥和番茄关于花发育的同源基因功能相似,因此可充分利用生物信息学方法,通过拟南芥已知开花基因来预测番茄的开花基因,并通过反向遗传学方法来验证预测的正确性,也不失为一种便捷有效的方法。拟南芥有6条开花诱导途径,其中光周期途径和自主途径在番茄上得到验证,未来可对番茄开花诱导途径进行更全面的探索。
通过突变体的创制、筛选及相关基因的克隆和鉴定在今后的开花研究中仍然会是最主要的手段之一,所以还需要更丰富的突变体材料来完善番茄开花调控网络。由于番茄较拟南芥更广的遗传背景和更加复杂的开花过程,所以番茄开花基因的研究比起拟南芥更具有挑战性。可利用F2代群体、近等基因系等来对相关基因进行研究。
对番茄基因组研究的进展为人们利用基因组学和蛋白组学的方法来研究番茄花发育相关的基因、并了解其作用模式提供了很好的机会。通过深入研究,最终可通过传统育种手段或基因工程手段改造番茄的花发育过程,例如通过改变环境或基因表达使开花提前从而缩短生育周期,或使无限生长型转变为有限生长型便于机械化管理,从而达到高产优质高效的目的。

花是植物最重要的经济器官,花期、花型及花序的分枝等重要性状都是花发育的外在表现.花发育的研究也是分子生物学和发育生物学研究中最活跃的领域。植物生理学的研究表明,成年植物经过一定的营养生长阶段,便由营养型分生组织进入感受态.此时如遇适宜的外界...
茉莉素这一类植物激素主要包括茉莉酸、茉莉酸甲酯、茉莉酸异亮氨酸、12-氧-植物二烯酸等环戊酮的衍生物。早在1962年,茉莉酸甲酯作为一种香料从素馨花中分离出来。至今,经过70多年的研究,人们逐渐认识了茉莉素的生物学功能、合成途径及其信号传导途径。...

目前,基于大量的研究发现,导致花粉败育主要与线粒体的呼吸作用异常、膜脂过氧化、物质代谢紊乱和小RNA功能异常等有着密切的关联。...
真核细胞的基因表达是高度复杂的过程,是遗传调控和表观遗传调控协同作用的结果.遗传调控以基因决定论为核心内容,表观遗传学则主要研究DNA序列以外的调控机制.已有研究结果证实,基因行为可通过表观遗传的行为进行调控(Reik,2007),并可遗传给...
在畜牧生产中,多胎动物的窝产仔数性能具有重要的经济价值。它主要受到情期排卵数、子宫容量及胎盘效率等因素的影响。在排卵数和子宫容量因素相对稳定的情形下,妊娠个体间胎盘发育的差异性可能导致胎盘效率的不同。胎盘是胎儿在母体内生长发育的重要器官。...
哺乳动物卵泡发育分子机制的相关研究已取得了巨大的成果。目前, 已经证明了许多信号通路在卵泡生长发育过程中起着非常重要的作用, 并且其中一些信号通路的研究已经非常透彻。...
, 随着对毛囊发育及毛发生长的影响因子及调控机制研究的不断深入, 已经有越来越多的与毛囊周期及其形态发育相关的基因、活性蛋白和信号通路陆续被发现并验证。相信将会有更多影响毛囊发育和毛发生长的因子和调控机制被发现和阐明, 为动物绒毛产业和人类临床治...