细胞生物学论文


椎间盘由髓核、纤维环和软骨终板 3 部分组成,是人体中最大的无血管组织。 四岁时椎间盘就几乎没有任何血管,间盘营养严重减少,髓核细胞群随之发生明显变化,脊索细胞被更多的能适应无氧环境的软骨细胞和纤维母细胞所代替; 十岁时颈椎间盘已经成为一个无血管的组织,其营养供应主要来自于软骨终板的渗透。 软骨终板中央部分渗透性较周围部分高。 软骨终板中央部分有 1/3 区域可以渗透,而周围部分仅约 1/10 区域可以渗透; 软骨终板在髓核处溶质渗透占整个软骨终板的 85%,在软骨终板接近内呈纤维环处下降到 35%,在软骨终板接近外层纤维环界面处几乎完全不能渗透。 椎间盘营养通路主要有终板途径和纤维环途径: 椎体内的血管不断分支,在软骨终板下骨质形成血管袢,包括氧气在内的绝大部分营养物质通过这些血管袢经扩散作用透过软骨终板、椎间盘基质,最终到达内部的椎间盘细胞,而乳酸等代谢废物经过相反途径排出,此即终板途径,Du 等通过动态增强 MRI 清楚观察到这一现象[1]; 小部分营养物质通过分布到最外层纤维环的血管末梢来提供养分,即纤维环途径[2 -3]. 但随着机体的老化,供应椎间盘周围的血管数量减少,软骨终板逐渐钙化,氧气等营养物质的供应及代谢废物的排除受到阻滞,椎间盘内逐渐形成缺氧微环境。 这种微环境对椎间盘退变的发生发展如何产生影响,椎间盘退变是否与缺氧环境有关,这些都成为近年来学者们研究的重要课题。 骨钙素是骨转化的重要指标,能反应骨质疏松的状况[4]. 最近研究发现骨钙素参与诱导椎间盘软骨终板的退变钙化过程,影响椎间盘退变[5]. 本实验拟通过体外培养髓核细胞,应用免疫学及组织化学原理,对组织切片或细胞标本中的某些化学成分进行原位的定性、定位或定量研究,判断缺氧环境是否是骨钙素( OCN) 变化的因素。
1 材料与方法
1. 1 主要试剂和仪器
永生化人髓核细胞株( 上海拜力生物) ,基础培养箱,HF100 三气培养箱,骨钙素( OCN) 检测试剂盒( 一抗,二抗,中杉金桥公司) ,磷酸盐缓冲液( PBS) ,4% 多聚甲醛试剂,曲拉通 X-100,H2O2溶液,牛血清白蛋白( BSA) ,DAB 显色试剂盒,CKX31-A12PHP 倒置显微镜( Olympus 公司) .
1. 2 免疫细胞化学染色检测髓核细胞常氧与需氧下 OCN 表达,采用免疫组织化学 SABC 法
制作髓核细胞爬片,于 37℃常氧培养箱,和模仿椎间盘缺氧( 3% O2) 条件的培养箱中培养; 于规定时间节点取出细胞爬片,PBS 缓慢洗涤 3 次,每次 5 min,室温干燥; 4% 多聚甲醛固定 20 min,PBS 缓慢洗涤 3 次,每次 5 min; 3%Triton-100 处理 20 min,PBS 缓慢洗涤 3 次,每次 5 min; 3%H2O2处理 15 min,PBS 缓慢洗涤 3 次,每次 5 min; 山羊血清或者 BSA 封闭 2 h,PBS 缓慢洗涤 3 次,每次 5 min; 一抗( 兔抗人 OCN 单抗( 1∶ 200) ) 4℃ 过夜封闭; 将玻片从 4℃ 取出,室温复温 30 min,PBS 缓慢洗涤 3 次,每次5 min; 室温孵育二抗( 生物素标记的羊抗兔 IgG 抗体) ,结合 2 h,PBS 缓慢洗涤 3 次,每次 5 min; 三抗( SABC) 封闭,室温下结合 2 h,PBS 缓慢洗涤 3 次,每次 5 min; DAB 显色 3 ~10 min,待细胞出现特异性染色后用水洗涤,镜检; 盐酸乙醇分化,氨水反蓝,镜检; 脱水,封闭; 拍摄,细胞核均染为蓝色,OCN 表达阳性细胞胞质均染为棕色。
1. 3 免疫组化图像分析
用 Image-Pro Plus( IPP) 6. 0 分析免疫组化图片,根据染色染料颜色的深浅( 光密度) 及分布面积大小来确定目标蛋白的量。 染色区域分布面积与目标蛋白量成正比。 染色的颜色深浅( 光密度) 与目标蛋白量的定量关系符合朗伯-比尔定律,是对数关系。 每张切片在高倍镜下( ×100) 随机选取 8 个视野,应用图像分析系统进行半定量检测,结果以平均灰度值表示。 平均灰度值高( 染色强度弱、透光度强) 表示OCN 表达水平低; 反之则表示表达水平高。 平均光密度 OD 值( 吸收光的物质的光学密度,与染色物质的量相关) 高,表示 OCN 表达水平高; 反之则表示表达水平低。 将图片上各点的光密度值累加起来,得到IOD. 此值与目标物质的总量成正比。 IOD 值除以细胞个数,得到单个细胞的 mean density,此值反映了单个细胞 OCN 表达的水平。
1. 4 统计学处理
应用 SPSS 19. 0 统计学软件进行数据处理。 检测数据以 m ± s 表示,组间比较采用单因素方差分析Tamhane's T2检验,以 P <0. 01 为差异有统计学意义。
2 结 果
2. 1 生长曲线分析
检测结果见图 1 ~2 在培养 48、72 h 时,缺氧组髓核细胞较常氧组细胞增殖速度减缓 >40%.

2. 2 常氧与需氧环境中髓核细胞 OCN 的表达髓核细胞 OCN 的表达检测结果见图 3 ~4 和表 1.
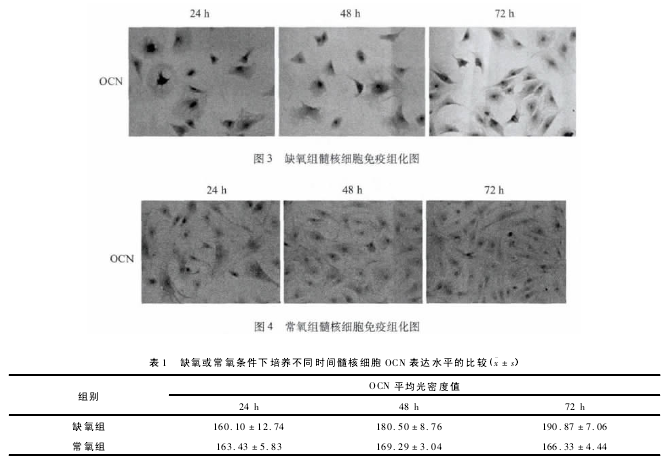
缺氧组与常氧组均可见 OCN 表达阳性细胞,且与常氧组比较,缺氧组多数阳性细胞的细胞质染色较深。 在培养 24 h 时,缺氧组髓核细胞 OCN 表达水平与常氧组比较,差异无统计学意义。 在培养48、72 h 时,缺氧组髓核细胞 OCN 表达水平均明显高于常氧组( P < 0. 01) ,且缺氧组髓核细胞 OCN表达水平随缺氧时间延长而逐渐升高( P <0. 01) ,常氧组各时间段之间髓核细胞 OCN 表达水平无统计学差异。
3 讨 论
近年来在椎间盘组织工程及生物学研究方面取得的进展,为椎间盘退行性变的生物学治疗提供了较好前景。 大量的研究表明,OCN 参与了椎间盘软骨终板的退变钙化过程,OCN 敲除可能抑制软骨终板的退变钙化[6]. Idelevich 等研究证实 OCN 过表达能促进血管平滑肌细胞和软骨细胞株的分化及矿化[7],而 OCN siRNA 能抑制主动脉血管的骨化和矿化[8].
本实验在体外成功构建了人髓核细胞缺氧模型,通过免疫组织化学方法,对不同缺氧时期 OCN 在体外培养髓核细胞中的表达进行定量检测,结果显示在培养 48、72 h 时,缺氧组髓核细胞 OCN 表达水平均明显高于常氧组( P <0. 01) ,且缺氧组髓核细胞 OCN 表达水平随缺氧时间延长而逐渐升高,说明缺氧可诱导髓核细胞 OCN 的表达上调,缺氧环境下髓核细胞的骨钙素过表达,这个结果也可以间接说明缺氧可能对椎间盘软骨终板矿化起到促进作用。
本实验有助于进一步揭示椎间盘退变的病理机制,为椎间盘退变的防治开辟新思路与新途径。
参考文献:
[1] DU H,MA S H,GUAN M,et al. Dynamic contrast enhancedmagnetic resonance imaging study of the nutrition pathway forlumbar intervertebral disk cartilage of normal goats[J]. Orthop Surg,2011,3( 2) : 106 - 112.
[2] GRUNHAGEN T,SHIRAZI-ADL A,FAIRBANK J C,et al. Intervertebral disk nutrition: a review of factors influencingconcentrations of nutrients and metabolites[J]. Orthop Clin North Am,2011,42( 4) : 465 - 477.
[3] URBAN J P,SMITH S,FAIRBANK J C. Nutrition of the intervertebral disc[J]. Spine,2004,29( 23) :2700 -2709.
[4] SCHAFER A L,SELLMEYER D E,SCHWARTZ A V,et al. Change in undercarboxylated osteocalcin is associated withchanges in body weight,fat mass,and adiponectin: parathyroid hormone( 1 - 84) or alendronate therapy in postmenopausalwomen with osteoporosis( the PaTH study) [J]. J Clin Endocrinol Metab,2011,96( 12) : 1982 - 1989.
[5] ZHANG Z M,JIANG L S,JIANG S D,et al. Differential articular calcified cartilage and subchondral bone in postmenopa-usal women with osteoarthritis and osteoporosis: two-dimensional analysis[J]. Joint Bone Spine,2009,76( 6) : 674 - 679.
[6] NG K W. Regulation of glucose metabolism and the skeleton[J]. Clin Endocrinol( Oxf) ,2011,75( 2) :147 -155.
[7] IDELEVICH A,RAIS Y,MONSONEGO-ORNAN E. Bone Gla protein increases HIF-1alpha-dependent glucose metabolismand induces cartilage and vascular calcification[J]. Arterioscler Thromb Vasc Biol,2011,31( 9) : 55 - 71.
[8] DUCY P,DESBOIS C,BOYCE B,et al. Increased bone formation in osteocalcin-deficient mice[J]. Nature,1996,382( 6590) :448 -452.

诱导性多能干细胞(inducedpluripotentstemcells,iPSC)是利用基因工程技术将动物或人的体细胞重编程而得到的一种类似胚胎干细胞(embryonicstemcells,ESC)的多能干细胞类型[1].本研究小组前期研究发现,维甲酸能诱导iPSC分化成神经细胞,并且能形...
卵母细胞体外成熟(IVM)是指将不成熟的卵母细胞,在体外模拟体内成熟的微环境,培养至成熟卵子(MⅡ)的过程。此项技术开始于动物实验,早在1935年Pincus发现兔子的不成熟卵可以在体外成熟,并且这个成熟过程是自发的。30年后,Edwars提出人类的不成熟卵母...
体外动物细胞培养已广泛应用于现代医学、生物学和分子遗传学等多个领域,是从事细胞水平及分子水平研究所采用的基本方法。在高原环境下心肌细胞的体外培养尚未见报道。本文拟通过心肌细胞系的体外培养,观察其生长和增殖状况,并通过绘制生长曲线,初步探讨...
藏獒(TibetanMastiff)属食肉目(Carnivora)犬科(Canidae),是原产于青藏高原地区的古老犬种,也是中国唯一的一个大型犬种,属于国家二级保护动物。由于近几十年来藏獒被大量贩运到内地省份,甚至国外,加上当地生存环境...
引言Introduction功能矫形治疗是口腔正畸学中矫治患者下颌发育不足、后缩畸形的主要方法。颞下颌关节是人体关节中能保持终生改建的惟一关节,而髁状突软骨作为下颌骨的主要生长中心,对矫形力的反应直接而又敏感,是临床功能矫形治疗的主要功能区域,同时...
肠神经嵴干细胞(entericneuralcreststemcells,ENCSCs)来源于迷走神经嵴(vagalcrest,VC)和骶神经嵴(sacralcrest,SC).VC来源的ENCSCs定植于整个消化道,构成肠神经系统的绝大部分肠神经节.该过程发生障碍,将造成肠道不同部位缺乏神经节细胞,局部肠管痉...
目前,国内外研究多集中于干细胞向神经细胞分化的研究,而对体细胞的研究相对较少。干细胞存在伦理学、免疫排斥以及潜在的致瘤性等问题限制了其在临床上的应用,而具有部分胚胎干细胞特性的人羊膜上皮细胞(humanamnioticepithelialcells,HAEC)不存在上述...

对男性、女性的外周血淋巴细胞进行染色体核型分析发现,在“男1”、“女2”视野中均可找到分散较好、形态较清晰且数量为46条的淋巴母细胞染色体。介...
血管平滑肌细胞(vascularsmoothmusclecells,VSMCs)是动脉发生钙化的主要细胞,最近研究发现血管钙化类似于骨的形成,它是由细胞介导的主动调节过程,同时存在平滑肌细胞表型转变,并与多个成骨相关类因子相关[1].以往体外研究动脉病变的平滑肌细胞...

在生命体中,从最小的单元―――细胞到高级的组织、器官,它们在识别及相互作用过程中不仅存在着生物化学反应,而且存在着力学传导,这些活动广泛参与并影响着有机体的生长、发育、分化等生理及病理阶段。...